
不同纬度植物群落系统发育与功能性状结构研究
2024-01-25 06:49苗乐乐刘旻霞肖音迪杨春亮王千月
生态学报 2024年1期
苗乐乐,刘旻霞,肖音迪,杨春亮,王千月,王 敏
西北师范大学地理与环境科学学院,兰州 730070
群落中物种的聚集是生态和进化过程共同作用的结果[1],考虑到当地生境条件,群落只能包含环境耐受性允许其存在的物种,同时物种在特定生态条件范围内的生存能力又受到其进化历史的制约。因此量化共存物种的生态和进化相似性对于阐明推动群落聚集的过程至关重要[2]。目前,生态学家一般是基于物种的系统发育相关性来间接估计相似性[3],或者通过量化多个功能特征来直接测量物种的相似性[2,4],这两种方法目前在群落生态学中得到广泛应用,而且研究表明物种系统发育关系与其功能属性相结合的方法提高了预测物种相互作用结果以及这些结果对进化过程影响的能力,同时对于预测生物多样性未来分布动态、生态系统功能和对全球变化的反应以及保护生物资源显示出越来越广阔的前景[5]。
以往对植物群落系统发育结构模式的研究普遍发现,在压力较大、资源有限的条件下,群落在物种之间表现出较高的系统发育相关性(聚类),因为环境过滤和系统发育生态位保守性相结合,会选择更适合植物群落的物种来适应这些极端环境[6]。相比之下,系统发育过度分散表明,竞争排斥和生态位进化可能占主导地位,因为在系统发育上更相关的物种被认为会更强烈地竞争群落中的有限资源[7-8]。Webb等[9]在阐述谱系结构研究方法时,指出生态性状的进化特征对群落构建的解释有着重要影响。植物功能性状是植物与环境长期互相影响的结果,作为影响植物体适应性的形态和物候特征属性[10],它可以进一步反映植物的养分权衡方式以及生存和适应策略。因此,可以将多种植物功能特性结合起来,以反映其沿环境梯度的响应特征,并揭示植物群落聚集的主要驱动因素[11]。
研究证实植物的叶片功能性状沿着环境梯度(气候、土壤等)表现出显著的梯度格局,而且处于相同环境条件下的不同的植物物种功能性状也存在一定差异[12]。纬度梯度作为包含多种环境因素的梯度效应,研究物种多样性的纬度模式对于探索物种对环境的动态响应、理解不同气候区群落聚集过程、预测气候变化对生物多样性的未来影响至关重要[13]。目前,生态学家对于不同气候带生境中的物种共存机制的研究大多是分散在各个纬度带上的独立讨论。比如在热带,程毅康等[14]对海南岛霸王岭热带云雾林群落谱系多样性和功能多样性的研究结果证明了生境过滤是驱动群落构建的关键因子;Kembel和Hubbel研究发现巴拿马热带雨林植物群落的谱系结构随着空间尺度的增大逐渐聚集[15]。在亚热带,对哀牢山森林群落的系统发育结构分析证实了生态位过程在群落构建中起到重要作用[16];在温带,对长白山针阔混交林不同演替阶段群落结构的研究表明随着演替阶段的进行和径级增大,竞争性排斥的作用逐渐明显[17]。以上研究结果虽然较好地解释了对应地带群落的研究问题,但是由于缺乏研究尺度上的连续性,结论有待基于不同纬度群落的连续研究进行验证和拓展。
甘肃省地理环境复杂,植被类型多样,由于栖息地的高度异质性和较少的人为干扰,兼具原始性和脆弱性两方面特征,成为了研究植被动态及其对环境变化响应的理想场所[18]。因此,本文以甘肃省不同纬度梯度的植物群落为研究对象,从甘南高山草甸延伸到黄土高原沟壑地区,显著而多变的气候梯度形成了区域植被和环境差异,丰富了栖息地类型,具有显著的纵向连通性和纬度隔离性,本研究基于物种、系统发育和功能性状拟探究以下内容:(1)群落的系统发育结构和功能性状结构是如何沿纬度梯度变化的? (2)群落系统发育结构和功能格局是否一致? (3) 通过研究密切相关的物种对环境变化的响应特征(即系统发育信号)的相似程度,来评估生态和进化过程对植物群落中物种聚集的相对贡献。
1 材料与方法
1.1 研究区概况
本研究区域选择了甘肃省甘南藏族自治州的玛曲县和合作市、临夏回族自治州的和政县、兰州市等地(图1),研究区位于中纬度温带(33°06′N-36°61′N,100°44′E-104°45′E),南北跨度500 km,海拔1474-4776 m,该区域的气候类型为温带大陆性气候和亚高寒湿润性气候,年平均气温为0-10.3℃,年降水量为266-615.5 mm,主要集中在6-8月,雨热同期。土壤类型以亚高山草甸土、黄绵土和棕钙土为主,研究区优势种为矮嵩草(Kobresiahumilis)、长毛风毛菊(Saussureahieracioides)、小花草玉梅(Anemonerivularis)、白氏马先蒿(Pedicularispaiana)、西北针茅(Stipasareptana)和蓍状亚菊(Ajaniaachilleoides)。

图1 研究区概况图Fig.1 Overview of the study area
1.2 野外调查和试验设计
2021年7月中旬至8月中旬进行野外群落学调查,通过实地考察,沿不同纬度(33°60′N-36°61′N)选择坡向一致的3座山地作为重复,分别在每座山地的北坡随机选取9个0.5 m×0.5 m的样方,记录每个样方的海拔、地理位置和样方内物种个数,多度,高度和盖度,采集 10 片完全熟透的叶子,暂时储存于自封袋中,并于 12 h 内扫描植物叶片,并用环刀法取0-10 cm土深的土壤样品。样地概况如表1所示。
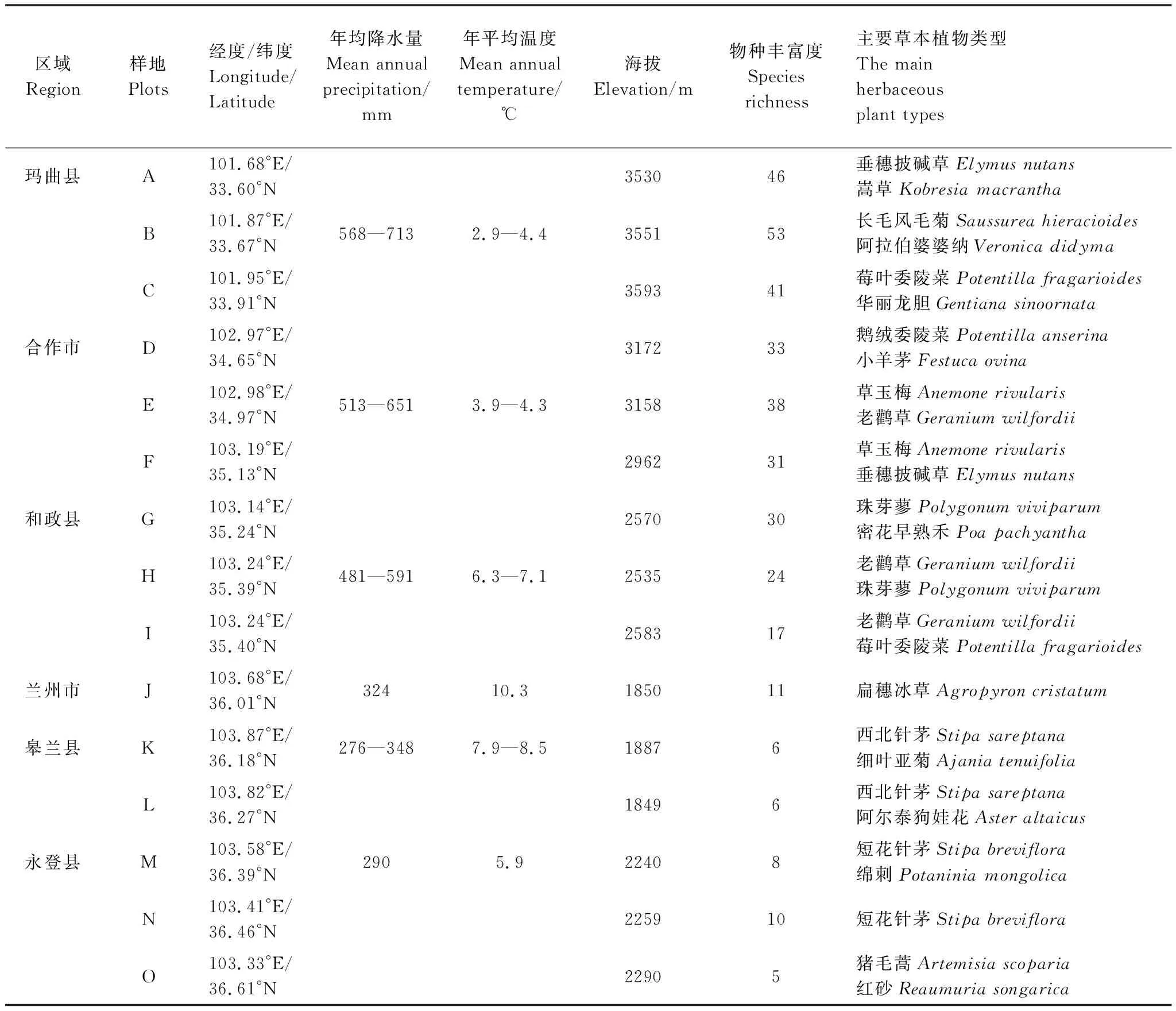
表1 采样点沿纬度梯度的分布情况Table 1 Distribution of sampling points along the latitudinal gradient
用烘干法测定土壤含水量,用pH计(ST3100,上海仪电科学仪器股份有限公司,上海)和电导仪(DDS-801A,上海仪电科学仪器股份有限公司,上海)分别测量土壤pH和电导率。土壤全氮含量采用微量凯氏法测定,土壤有机碳含量采用重铬酸钾容量法测定,土壤全磷含量采用钼锑抗比色法测定[19]。
降水量和气温数据由中国气象科学数据共享服务网平台提供(http://data.cma.cn/),质量可靠。根据 PérezHarguindeguy 等[20]的植物功能性状采集手册,选取代表群落物种叶性状和茎干性状的共 7类关键功能性状(表 2)。选择样品中的 10 个完全成熟个体来测定植物高度,选用精度为0.02 mm的游标卡尺测量叶片厚度,使用扫描仪(CanoScan LIDE 110,北京,中国) 测量了每个物种,并用 Image J 软件计算叶面积,比叶面积,将扫描后的叶片放入烘箱,烘干至恒重后称量叶片干重,计算叶干物质含量;叶片碳含量、叶片磷含量和叶片氮含量的测定同土壤[19]。
1.3 群落物种多样性和谱系多样性的计算
群落物种多样性采用物种丰富度指数(S)、Shannon 多样性指数(H)、物种均匀度(E)[21]来进行表示,谱系α多样性采用Faith′s PD 指数量化,多样性指数H和均匀性指数E的具体计算公式分别如下:
H=-∑PilnPi
(1)
E=H/lnS
(2)
式中,Pi为第i个物种的相对重要值,S表示样本内物种总数。
1.4 谱系树构建
通过调查研究样地中共出现了104个物种,在TPL(The Plant List)网站(http://www.the plant list.org/)对物种定名进行规范。然后将校正后的物种按科属种的格式输入到Phylomatic-awk软件中,基于Zanne 2014谱系数据库生成具有进化信息的系统发育树框架,用软件FigTree1.4.4构建系统发育树。
1.5 群落谱系结构和功能性状结构
采用净亲缘关系指数(Net relatedness index,NRI)和平均成对性状距离(Mean pairwise trait distance,PW)来表示群落系统发育结构和功能性状结构。NRI 是将群落内的物种两两间系统发育平均距离( Mean pairwise nodal distance,MPD)与零模型随机化之后的结果进行比较得到的。(S.E.SPW)是将平均成对性状距离与功能性状聚类树末端的物种随机置换999 次获得的零模型比较所产生的标准化距离[22]。计算公式分别为:
(3)
(4)
式中,MPD表示物种平均系统发育距离,MPDsample和MPDrandsample分别代表实际观测值和随机模拟的零群落的平均谱系距离,mean(MPDrandsample)代表零群落的MPD的平均值,sd(MPDrandsample)为随机模拟下群落的标准差;性状中各值含义同理。若NRI>0,说明物种在系统发育结构上聚集,NRI<0,说明物种在系统发育结构上发散,NRI=0,说明系统发育结构随机;S.E.SPW>0,代表物种功能性状结构聚集,S.E.SPW<0,代表物种功能性状结构发散,S.E.SPW=0,代表物种功能性状结构随机。
1.6 系统发育信号检测
本研究采用Brownian motion 进化模型的K值检验功能性状的系统发育信号[23]。若K>1,则表明物种的功能性状比布朗运动模型预测的要近,显示存在系统发育信号,功能性状表现出系统发育保守性。若K=1,则表明功能性状和系统发育结构没有关系;若K<1,则表示系统发育相近的物种,它们的功能性状比布朗运动模型预测的要远,群落系统发育结构和功能性状结构不一致。功能性状系统发育信号的显著性采用与零模型比较的方式来衡量,若是实际值大于零模型值的次数达到950次以上(P<0.05),就认为功能性状表现出显著的系统发育信号。
1.7 数据的统计与分析
植物群落的物种以及系统发育多样性指数分别用R软件的程序包vegan、FD和picnate计算。用IBM SPSS Statistics 25对不同纬度的系统发育指数和功能性状结构指数进行单因素方差分析判断其显著性差异,所有统计分析的显著水平设定为P<0.05。使用统计软件R4.1.2中Kcalc 和multiPhylosignal 函数分析获取K值并检验系统发育信号的显著性,用RLQ(R-mode linked to Q-mode)结合第四角分析(Fourth-Corner)的方法将样方、环境变量和植物功能性状3种数据集组合成一个可描述性状-环境关联矩阵,进而确定物种丰度关联的环境梯度与物种性状之间的关系,使用R语言中的ggplot2 包作图。
2 结果
2.1 不同纬度物种多样性和谱系多样性的变化
由图2可以看出,Shannon-Wiener指数随纬度增大显著降低(P<0.05),最大值出现在低纬度地区(33.6°N),这说明在低纬度梯度上有较高的物种多样性。植物群落的物种丰富度、谱系α多样性随纬度增加有相同的变化趋势,即从低纬度到高纬度物种数显著减少(P<0.05),谱系多样性也减小,最小值均出现于高纬度梯度。Pielou均匀度指数随纬度的增加无显著变化趋势。

图2 不同纬度植物群落物种及谱系α多样性的比较(平均值±标准误差)Fig.2 Comparison of species and lineage α diversity of plant communities at different latitudes(mean±SD)
2.2 植物群落物种谱系组成
基于被子植物的分类系统APGIII对研究区104种草本植物构建的系统发育树如图3。系统发育树包括结点和进化分支两部分,其中分支节点表明历史物种的结束以及新物种的开始,各分支代表不同的亲缘关系,分支长度代表物种的进化历史。其中菊科和禾本科植物为群落优势种,在纬度33.6°N-36.7°N 的研究区域内均有分布。群落内分化时间最短的风毛菊(Saussureajaponica)和长毛风毛菊(Saussureahieracioides)大约在0.3 Ma时首先聚为一类,随后又与大刺儿菜(Cirsiumsetosum)在0.6 Ma时聚类,最终与蒲公英(Taraxacummongolicum)、火绒草(Leontopodiumleontopodioides)与细叶亚菊(Ajaniatenuifolia)等17种菊科植物在45.7 Ma时聚为一类,表明它们由共同的祖先进化而来。像甘肃马先蒿(Pediculariskansuensis)、蒙古芯芭(Cymbariamongolica)和白氏马先蒿(Pedicularispaiana)同属列当科,具有同一节点,分支长度相同,表明其进化时间相同。

图3 不同纬度植物群落系统发育树Fig.3 Phylogenetic trees of plant communities at different latitudesPotentilla anserina:鹅绒委陵菜; Potentilla fragarioides:莓叶委陵菜; Potentilla kleiniana:蛇含委陵菜; Potentilla polyphylla:多叶委陵菜; Potenilla bifurca:二裂委陵菜; Fragaria vesca:野草莓; Potentilla saundersiana:钉柱委陵菜; Potentilla fruticosa:金露梅; Sanguisorba officinalis:地榆; Spiraea salicifolia:绣线菊; Potaninia mongolica:绵刺; Gueldenstaedtia verna:米口袋; Medicago falcata:黄花苜蓿; Medicago sativa:紫苜蓿; Oxytropis kansuensis:甘肃棘豆; Thermopsis fabacea:黄华; Lespedeza bicolor:胡枝子; Caragana tibetica:毛刺锦鸡儿; Caragana liouana:中间锦鸡儿; Ranunculus tanguticus:高原毛茛; Trollius farreri:矮金莲; Anemone trullifolia:条叶银莲花; Delphinium grandiflorum:展毛翠雀; Thalictrum aquilegifolium:唐松草; Anemone rivularis:小花草玉梅; Gymnaconitum gymnandrum:露蕊乌头; Stellaria media:繁缕; Sagina japonica:漆姑草; Leontopodium leontopodioides:火绒草; Ligularia sibirica:橐吾; Picris hieracioides:毛莲菜; Anaphalis lactea:乳白香青; Ajania tenuifolia:细叶亚菊; Taraxacum mongolicum:蒲公英; Saussurea japonica:风毛菊; Saussurea hieracioides:长毛风毛菊; Aster alpinus:高山紫菀; Aster altaicus:阿尔泰狗娃花; Saussurea frondosa:狭翼风毛菊; Ajania achilleoides:蓍状亚菊; Tripolium vulgare:铁杆蒿; Cirsium setosum:大刺儿菜; Artemisia capillaris:茵陈蒿; Artemisia scoparia:猪毛蒿; Asterothamnus alyssoides:紫菀木; Kobresia humilis:矮生嵩草; Kobresia macrantha:嵩草; Imperata cylindrica:白茅; Brachypodium sylvaticum:短柄草; Leymus secalinus:赖草; Poa poophagorum:波伐早熟禾; Elymus dahuricus:披碱草; Elymus nutans:垂穗披碱草; Koeleria cristata:洽草; Festuca ovina:羊茅; Poa pachyantha:密花早熟禾; Stipa sareptana:西北针茅; Setaria viridis:狗尾草; Stipa grandis:大针茅; Agropyron cristatum:扁穗冰草; Stipa breviflora:短花针茅; Cleistogenes songorica:无芒隐子草; Mentha canadensis:野薄荷; Leonurus japonicus:艾草; Salvia japonica:鼠尾草; Lavandula angustifolia:薰衣草; Scutellaria baicalensis:黄芩; Pedicularis paiana:白氏马先蒿; Pedicularis kansuensis:甘肃马先蒿; Cymbaria mongolica:蒙古芯芭; Gentianopsis barbata:扁蕾; Gentiana macrophylla:秦艽; Gentiana sinoornata:华丽龙胆; Halenia corniculata:花锚; Gentiana abaensis:阿坝龙胆; Swertia diluta:北方獐牙菜; Bupleurum chinense:柴胡; Saposhnikovia divaricata:防风; Euphorbia esula:乳浆大戟; Rumex acetosa:酸模; Polygonum viviparum:珠芽蓼; Veronica polita:婆婆纳; Plantago asiatica:车前; Veronica didyma:阿拉伯婆婆纳; Geranium wilfordii:老鹳草; Erodium stephanianum:牻牛儿苗; Adenophora stricta:沙参; Humulus scandens:拉拉藤; Descurainia sophia:播娘蒿; Asparagus cochinchinensis:天门冬; Viola diffusa:七星莲; Viola striatella:圆叶堇菜; Viola dissecta:裂叶堇菜; Euphorbia fischeriana:狼毒; Allium sikkimense:高山韭; Lycium chinense:枸杞; Zygophyllum mucronatum:蝎虎驼蹄瓣; Incarvillea sinensis:黄花角蒿; Peganum harmala:骆驼蓬; Linum perenne:宿根亚麻; Convolvulus ammannii:银灰旋花; Reaumuria songarica:红砂; Kalidium foliatum:盐爪爪; Androsace umbellata:小花点地梅
2.3 植物叶片性状沿纬度梯度的分布规律
将叶片性状与物种组成和环境变量联系起来的RLQ分析表明(图4),轴1表征了从降水充沛、有机碳含量高的低纬度生境到位于温暖干燥的高纬度地区的偏碱性土壤的土壤养分梯度,高大的植物一般分布在温度较高的地区和相对贫瘠的土壤中,而矮小且比叶面积大的植物则出现在比较肥沃的土壤中,除了降水和温度梯度,植物也沿着土壤电导率和土壤氮磷梯度分布,如第二RLQ轴所示,叶片氮含量高的物种更容易出现在土壤电导率较低的富氮土壤中。
多变量排列检验(图5)揭示了不同纬度植物叶片性状和环境变量之间显著但微弱的整体关系(模型2,P=0.001,模型4,P=0.572)。植物高度与温度显著正相关,比叶面积与土壤含水量显著正相关。个体叶片性状与环境因子之间的这种弱双变量关联可以解释植物整体的功能聚类。

图5 FDR (False Discovery Rate)方法进行P值校正的第四角方法检验结果Fig.5 Test result of the fourth corner method for P-value correction by FDR methodP=0.05显著水平,红色表示功能性状与环境变量呈显著正相关,蓝色表示功能性状与环境变量呈显著负相关
2.4 不同纬度植物群落系统发育结构和功能性状结构
不同纬度群落系统发育和功能性状结构如图6所示,不同纬度的谱系结构均非随机(Studentt检验,P<0.05)。纬度小于35.5°N时,系统发育多样性指数NRI<0,随着纬度的增加,NRI逐渐增大直至36.0°N处变为正值,表明群落系统发育结构随着纬度增大发散程度减弱到聚集的变化趋势,在纬度36.4°N处,群落系统发育结构又变为发散状态。所有植物群落的功能结构均非随机(Studentt检验,P<0.05),在低纬度和高纬度处S.E.SPW都大于0,表现为功能聚集,在中纬度(35.2°N-35.5°N)主要表现为功能发散(S.E.SPW<0)。

图6 植物群落的系统发育和功能性状结构在不同纬度上的分布Fig.6 Phylogenetic and functional trait structures of plant communities distribution at different latitudes基于系统发育距离和成对性状距离的标准效应值的平均值±95%置信区间表征系统发育和功能性状结构,***,P<0.001;**,P<0.01;*,P<0.05
2.5 功能性状的系统发育信号
本研究区不同纬度植物群落的7个功能性状的K值均小于1,说明该区域的植物群落未表现出较强的系统发育保守性。其中叶干物质含量、叶片碳含量、叶片磷含量、叶片氮含量、叶片厚度和比叶面积不具有显著性的系统发育信号,只有植物高度表现出微弱的系统发育信号且种间差异大(P<0.05)(表2、图7),说明植物功能性状与物种遗传进化关系不大,而环境因素对其种间变异有一定的影响。

表2 植物功能性状的系统发育信号Table 2 The phylogenetic signal of plants functional traits
3 讨论
3.1 群落物种多样性和谱系多样性沿纬度梯度的变化
纬度梯度上水热条件变化显著,研究物种多样性的纬度模式对于探索物种对环境动态的响应、理解群落聚集过程、预测气候变化对生物多样性的未来影响至关重要[13]。相关研究表明一般大尺度上群落物种多样性的纬度梯度格局表现为随纬度增加由赤道到高纬地区物种多样性逐渐减少的趋势[24]。本研究通过对甘肃不同纬度植物群落研究发现,Shannon-Weiner 指数,物种丰富度和谱系α多样性随纬度增加而显著降低(P<0.05)(图2),这与全球大陆生态系统的变化格局一致,也与石娇星等[25]对中国东部海岛黑松群落的研究结果符合。这主要是因为低纬度处的高寒草甸降水量丰富,光照充足,水热条件相对优越,根据“能量假说”,优越的水热条件适合更多的物种生存[26]。而较高纬度地区降水稀少且集中,蒸发量大,土壤表层盐碱累积强烈,植被受水盐双重胁迫[27],生态环境脆弱并容易受外界干扰,可维持的物种多样性也就较低(图3)。
3.2 植物叶片性状对纬度变化的响应特征
叶子是植物光合作用的重要功能器官,也是与周围环境接触最多的器官。利用叶片性状的变化来研究植物对环境的适应性已成为研究植物群落结构的重要手段之一[28]。本研究结果表明在有机质和降水量丰富的土壤中,植物叶片的氮磷含量更高(图4),这是因为植物地上部分元素特征通常与土壤养分含量有一定的相关性(图5),高寒草甸属于低纬度寒冷湿润区,微生物活性偏低,植物残体、有机质等分解速率较慢,腐殖层较厚,土壤养分条件优越,植物通过根系吸收土壤养分,又以凋落物的形式将C、N、P归还土壤,形成生态系统养分转化的有效循环过程[29]。而在黄土高原半干旱区较为贫瘠的偏碱性土壤中,比叶面积较小、叶干物质含量高的植株更为普遍,这表明植物对干旱和贫瘠环境具有高度的适应性和耐受性。植物为了维持自身新陈代谢,倾向于更好的保护内部资源,而采取减小比叶面积,增加叶厚度的生存策略以减少水分蒸腾,提高水分利用率,并且为了增强耐旱性会积累更多的干物质来抵抗不利环境[30-31]。
3.3 系统发育和功能性状结构沿纬度的变化
物种的形成是生态与进化过程共同作用的结果,通过对群落系统发育和物种功能特征的研究,有助于我们从进化历史和生态适应的角度理解群落聚集过程[32]。在本研究中植物群落的系统发育相关性随着纬度增加先增大后减小(图6),在纬度梯度上表现出明显的谱系结构(P<0.05)。低纬度处高寒草甸植物群落的系统发育结构是发散的,这是因为此处水热条件适宜,人为干扰较少,所以种间竞争排斥作用增强[9],从而驱动共存物种的生态位分化使各物种间的相似性受限以减少竞争。这与郝淑君[33]和王诗韵等[34]的研究结果一致;另一种原因是当物种间促进作用(如保育效应)有助于耐受恶劣环境时,物种间生态位重叠极大,补充了资源的各种利用方式,基于此系统发育结构也可能过度分散[35],这与luo[36]在玉龙雪山发现种间促进作用有助于植物适应极端高山气候并保持物种丰富度的研究一致。随着纬度增加植物群落系统发育结构聚集,说明在高纬度上非生物过滤在物种形成过程中占优势[6],主要原因是黄土高原干旱少雨,土壤贫瘠,而且容易受到放牧压力,植物受到强烈的环境胁迫同时生境过滤作用会选择与干旱条件相适应的物种生存,从而导致亲缘关系较近的物种聚集在一起形成谱系聚集的结构,在纬度36.4°N处,物种间系统发育相关性降低,该区域年平均温度5.9℃,降水量小于300 mm,气候干旱,生境恶劣,但是群落表现出发散的谱系结构,说明相似性限制在物种聚集过程中起重要作用[9]。有研究表明,资源匮乏条件下物种生存的核心是对地下资源(尤其是地下水)的获取[37],而对有限资源的激烈竞争必然导致相同生态位的物种难以共存。以红砂(Reaumuriasongarica)+绵刺(Potaniniamongolica)群落为例,红砂为浅根性植物,侧根较多,根系多分布在地下20-60 cm的范围内;而绵刺主根发达,可以利用地下80 cm以下的水分[38]。即亲缘关系较远的物种分别利用不同层次的资源,从而导致谱系发散。
植物群落功能多样性指数在高、低纬度上均高于恶劣环境下的随机预期,表现出功能聚集,而在中低纬度上显示为功能发散。这表明在低纬度和高纬度上环境过滤对于物种性状格局的形成起重要作用[6]。由于环境过滤作用,植物性状发生收敛进化,最终不同谱系的物种以相似的功能特性适应恶劣环境。与王寅等[39]在干旱内陆河流域的植物群落中,环境筛选在群落构建过程中发挥主导作用的研究结果一致。这些研究表明,群落倾向于沿着环境压力增加的梯度进行功能性状聚类[40]。中纬度上植物功能结构发散说明种间竞争作用占优势,该区域地处青藏高原与黄土高原交汇地带,环境条件趋于温和,当土壤肥力较高时,竞争通常会增加其重要性[41]。因此,本研究中植物群落系统发育结构和功能性状格局在纬度梯度上并不一致,就生物相互作用、非生物过滤以及环境变化和物种形成的历史过程而言,沿纬度梯度的群落聚集模式在区域之间是复杂多变的。
3.4 功能性状的系统发育信号
植物功能性状作为系统发育结构的一部分,它是区分物种生态位的主要属性,性状的变化取决于系统发育过程以及环境条件。因此,通过检验植物功能性状的系统发育信号来分析物种进化历史和环境之间的关系对于研究物种共存过程至关重要。
在本研究选取的7个植物功能性状中,只有植物高度表现出微弱的系统发育信号(P<0.05)(表2),这可能源于植物普遍需要长高和获得更高的光照环境,其他性状都没有检测出显著的系统发育信号(图7),这表明植物叶片性状主要受环境因素影响,与物种进化关系不大,亲缘关系相近的物种并没有表现出相似的性状特征,功能性状格局与系统发育结构不一致,这与王诗韵[34]、车应弟等[42]在艾比湖流域和亚高寒草甸的研究结果一致。这是因为研究区域地理环境复杂,植被类型多样,导致生境异质性增大,植物为了适应恶劣的生境,从而表现出功能性状上的一致性。以往有研究表明多数功能性状具有显著的系统发育信号[43-44],当然系统发育和功能特征的不一致模式可能源于功能特征采样的不完整,从而观察到的数据不能完全代表物种的实际生态位。
4 结论
纬度梯度上植物群落的功能性状格局与系统发育结构不一致。随着纬度增加,系统发育结构先由发散趋于聚集状态,随后在高纬度上又表现出发散的谱系结构,表明种间竞争作用减弱,环境过滤作用逐渐增强,随纬度继续升高相似性限制作用在物种聚集过程中占优势;而功能性状在中低纬度上发散,其他纬度上表现为聚集结构,说明植物群落形成过程中环境过滤作用明显。土壤含水量、土壤酸碱度和年平均气温等环境因子对植物性状格局影响较大,不同的气候条件和土壤养分特点发育了与之相适应的物种。本研究有助于了解不同气候带植物多样性的地理分布格局和潜在的生态维持机制。
致谢:感谢兰州大学高寒草甸与湿地生态系统定位研究站在试验中给予的支持和帮助。
猜你喜欢
艺术品鉴(2022年16期)2022-07-09
数学物理学报(2021年6期)2021-12-21
河池学院学报(2021年1期)2021-07-10
景德镇陶瓷(2021年1期)2021-03-24
应用数学(2020年2期)2020-06-24
数学年刊A辑(中文版)(2018年2期)2019-01-08
齐鲁周刊(2017年29期)2017-08-08
东方考古(2017年0期)2017-07-11
技术经济与管理研究(2014年11期)2014-03-11
河南科技(2014年3期)2014-02-27
