
Aviary measurements of dominance and affiliation between members of mixed-species birds flocks in southern China
2024-01-22 08:25:00JichongChenEstelleMeuxCiyunLiAiwuJingEbenGoodle
Avian Research 2023年4期
Jichong Chen, Estelle Meux,b, Ciyun Li, Aiwu Jing,*, Eben Goodle
aGuangxi Key Laboratory for Forest Ecology and Conservation,College of Forestry,Guangxi University,Nanning,530004,China
bDivision of Ecology and Biodiversity,School of Biological Science,The University of Hong Kong,Hong Kong,China
cDepartment of Health and Environmental Science,Xi’an Jiaotong-Liverpool University,Suzhou,215123,China
Keywords:Dominance hierarchy Keystone species Mixed-species animal groups Sociality
ABSTRACTIn mutualistic interactions, all parties are usually considered to benefit; yet there may be asymmetries in mutualisms where some partners/individuals benefit more than others.Such is thought to be the case in mixedspecies flocks of birds, where following species are thought to benefit more than leading species, and leading species may not be able to escape the association if they are subordinate to other species.We measured dominance and affiliation patterns of a mixed-species flock system of southern China in an aviary where these variables could be measured in a standardized way.In eight wild-caught flocks, the leading species, David’s Fulvetta(Alcippe davidi), was usually among the more subordinate flock members (ranking 9 of 13 species, with the the most dominant species ranked number one, and dominance measured by normalized David’s scores).Dominance was strongly influenced by body mass, and not by bill length or the number of individuals.Female fulvettas in particular tended to be among the most subordinate individuals in the flock.There was evidence of a negative relationship between a species pair’s affiliation, measured as the percentage of all observations in which these two species were found perching together, and their difference in ranking in the dominance hierarchy, particularly when some ground species, which are not likely to remain in flocks long term, were removed from the analysis.Species pairs that had more pronounced differences in their dominance rankings were less likely to be affiliated, which is consistent with the idea that subordinate species may avoid dominants.David’s Fulvetta is a strong information provider, so other species are likely benefitted in this relationship.Our data suggest one reason that fulvettas stay in flocks is that they may be subordinate and therefore have little control over who associates with them.
1.Introduction
Mutualisms, interactions between species in which all the different partners are benefitted, have been increasingly recognized to be common, and are essential for ecological functioning of ecosystems (Janzen,1985; Bruno et al., 2003; Bronstein, 2015).In mutualisms, benefits such as nutrition, energy, protection and transport are exchanged between interacting species.Yet because mutualisms are composed of individual organisms that are trying to maximize their survival and reproduction,there is often a drive for reduced investment, in which one organism tries to pay a minimal cost in order to obtain a greater benefit from its partner (Bronstein, 2001).These interactions can be shaped by the traits of the different individuals/species involved.For example, some species might be more dominant than others, where dominance is defined as repeated aggressive interactions among members of a social group that result in stable asymmetric relationships between individuals, so that higher-ranking animals consistently defeat lower-ranking animals (Lea et al., 2014; Holekamp and Strauss, 2016; Hobson, 2020).Dominant individuals or species may be able to attain resources before other mutualistic partners and therefore have higher body condition (Krams et al., 2020a; Coppinger et al., 2023) or they may punish their partners if they try to cheat them (Bshary and Grutter, 2005; Mills and Cote, 2010).Thus, studies of dominance are important to understand the maintenance of mutualistic social systems (Coppinger et al., 2023).
Mixed-species groups of animals are a type of mutualism found in many taxa, including invertebrates, but common in vertebrates, and most frequently found in birds (Morse, 1977; Goodale et al., 2017).Focusing on mixed-species bird flocks, it seems that all species may not be benefitted equally in such flocks, with leading species, also known as“nuclear species”, benefitting less than following species.Leaders tend to be conspicuous and noisy and therefore easy to follow (Hutto, 1994),and they are often gregarious in that they have many individuals per flock (Goodale and Beauchamp, 2010).Due to these behavioral traits and because they have an audience of conspecifics to communicate with,leaders often transmit a lot of information, information that can be“eavesdropped” on and used by other species (Goodale et al., 2010).In general then, followers benefit more than leaders, since they use the information provided by the leaders and do not have to seek the information themselves.Indeed, several studies that have measured foraging benefits to flocks have found leaders to benefit less often than followers(Dolby and Grubb, 1998; Hino, 1998; Sridhar et al., 2009).This raises the question of why leaders remain in mixed-species flocks: perhaps the costs imposed by the followers are not very great, or perhaps they are unable to get away from the followers, especially if they are subordinate to them (Goodale et al., 2017).Yet there have been few studies that investigate dominance in mixed-species group situations.This is particularly true of laboratory studies, in which resources can be standardized and thus the relative dominance of different individuals can be accurately measured under controlled conditions (but see Pierce and Grubb, 1981; Waite and Grubb, 1988; Dolby and Grubb, 1998).
Several types of factors may underlie the dominance hierarchy of organisms, and particularly that of birds.In the most common case,dominance is determined by intrinsic factors such as body size, fighting ability, personality traits, or other attributes that directly affect the ability to win fights (Chase and Seitz, 2011; Bush et al., 2016; Holekamp and Strauss, 2016).Within a species, other factors can shape dominance,even in the absence of individual differences in specific attributes (Chase et al., 2002; Hsu et al., 2006; Yasuda et al., 2014), such as winner–loser effects, in which victorious individuals subsequently behave more aggressively, whereas losers behave more submissively (Dugatkin and Reeve, 2014; Benelli et al., 2015; Laskowski et al., 2016), or the number of individuals in a group affecting the probability of winning intergroup encounters (Majolo et al., 2020).Dominance patterns can also be complicated if they are not linear, but rather intransitive (A dominates B, B dominates C, but C dominates A) (Zamudio and Sinervo, 2000).For birds, however, interspecific hierarchies have been found to have few intransitive relationships, and to be largely based on body size (Martin and Ghalambor, 2014; Miller et al., 2017; Leighton et al., 2022).As beaks can be used as weapons, beak length can predict dominance once body size is accounted for (Leighton et al., 2022), but there is little evidence that group size influences interspecific dominance (Shelley et al.,2004).
In this study, we moved wild-caught mixed-species flocks of southern China into aviary conditions and measured the relative dominance of the different individuals.In particular, we wanted to understand the relative dominance of the leading species of this system, David’s Fulvetta(Alcippe davidi), a gregarious species that is known to produce a lot of vocal information about predators (Jiang et al., 2020).We asked if species differed in their dominance, and if so, what factors seemed to be important to predicting dominance.We hypothesized that body size would be the most critical factor, with beak length a potentially important secondary factor, and that David’s Fulvettas would be subordinate to other members of the flock community because they are small birds, even though there are usually more fulvetta individuals than other species.In addition, we wondered whether the sex of fulvettas would influence their dominance ranking, and so we tested the individuals of this monomorphic species to determine their sex.Finally,we hypothesized that species that are different in dominance from each other would be less likely to be found affiliated together because that relationship would be threatening for the subordinate birds.We therefore measured both dominance (in interactions for access to food,measured both within and between species) and affiliation (the amount different species pairs were found on the same perch, measured between species only).
2.Methods
2.1.Study system
We studied mixed-species bird flocks of southwestern Guangxi Zhuang Autonomous Region, China.This region of southwestern Guangxi consists of limestone karst forest and has a southern subtropical monsoon climate, with annual temperature that varies between 13.8◦C(winter) and 28.1◦C (summer), and precipitation of 1200 mm annually(Qin et al., 2013).Karst forest does not have a very high canopy (20 m at most, often 10 m), and in general the flocks include understory, midstory and canopy species, and are similar to flock systems that have been described in northern Guangxi and Yunnan (Zhou et al., 2019), and Guangdong (Zhang et al., 2013, 2020).They are relatively small systems(averaging 4.6 ±1.4 species, 10.8 ±5.0 individuals; Jiang et al., 2020),much smaller than more diverse systems in the tall rainforests of Hainan(Zou et al., 2011) and Taiwan (Chen and Hsieh, 2002).
In all these forests of southern China, closely related species of fulvettas (Alcippespp.) act as flock leaders (Zou et al., 2007), with some gleaning species (like babblers and laughingthrushes in the understory,tits in the midstory, minivets and warblers in the canopy), sallying species (e.g., flycatchers, fantails and drongos), and even some nuthatches and woodpeckers following them (Goodale et al., 2015; Zhou et al., 2019).In southwestern Guangxi it is David’s Fulvetta that is the leader, being both the most frequent species in flocks (83% of flocks) and the most gregarious (3.4 individuals ± 2.3 [SD]) (Jiang et al., 2020).Although the most gregarious species in these flocks, it is much less gregarious than the leadingAlcippespecies in other systems (for example, there are 32.5 individuals of the leading species in Taiwan;Chen and Hsieh, 2002).The southwestern Guangxi flocks also tend to have few sallying (e.g., no drongos) and woodpecker species than other systems (Jiang, 2007; Jiang et al., 2020).
2.2.Flock capture
We captured mixed-species flocks in three sites: (1) Nonggang Nature Reserve, in Longzhou County, (2) Zhubu Forest, also in Longzhou County, and (3) Longmen Forest in Daxin County.Altogether we captured eight mixed-species flocks.We put out four to ten 3 m ×10 m mist nets in an area of radius ~25 m, and attempted to capture the whole flock, although because the nets were on the ground we missed most species that tend to inhabit the midstory and canopy (see Discussion section).We used playback of David’s Fulvettas to attract the flock to the area.All birds were captured in the same area within the same day, or if some part of the flock was captured in the late evening, we continued to try to capture more birds the next morning.Some ground-dwelling species (Buff-breasted BabblerPellorneum tickelli, Red-flanked BluetailTarsiger cyanurus, Siberian RubythroatCalliope calliope, Streaked Wren BabblerGypsophila brevicaudata, and White-tailed RobinMyiomela leucura) were caught in the same area as the flock; these species are unlikely to be members of the flock for a long time, although they could interact with the flock when it moves though their territory.Birds were only caught during the non-breeding season (September until March).Birds were captured with the approval of the Chongzuo Forestry Bureau.
2.3.Aviary conditions
After the capture of a mixed-species flock, the birds were transported in padded crates back to Guangxi University campus in Nanning,Guangxi.We then measured each bird (for body mass, beak length, wing length, and tail feather length), took a blood sample, placed a plastic ring on its leg so that it could be individually recognized, and introduced it to the aviary.The aviary was 7 m long × 3 m wide × 2.5 m high,constructed of metal mesh and with a large fiberglass canopy roof to provide shade and reduce the wind.The aviary was located next to a work room with a window that allowed the observers to watch the aviary and make audio recordings.All the birds were kept for approximately five days of habituation until the experiments started.An unlimited amount of mealworm and water were placed in the center of the aviary to ensure adequate food.Ten branches of diameter 2–3 cm and a length of 1 m were placed as horizontal perches for the birds at between 1.5 and 2 m in height.Including these experiments as well as subsequent experiments on the vocalizations of the birds, a mixed-species flock stayed in the aviary approximately 15 days, after which it was released at its capture site.Blood from the fulvettas was later tested to acquire information about the birds’ sex through chromosomal testing.The conditions of captivity and the experimental methods were approved by the Animal Experimental Ethical Inspection of Guangxi University(GXU2019-078).
2.4.Dominance observations
On the night of the fifth day in the aviary, food was removed at around 5:00 p.m.The next morning, mealworms were given in one bowl(3 cm diameter and 0.5 m high off the ground).This bowl was small enough that only one bird could forage from it at a time, and this situation caused the birds to make aggressive behaviors in order to monopolize the food.We let the birds forage at the bowl for 15 min a day, after whichad libitumfood was again provided to them.During the 15-min trial, the interactions at the feeding bowl were observed by the observer (JC) inside the observation room and were also videotaped by a JVC video camera (model EverioR GZ-R465), placed 0.10 m away from the feeding bowl to provide adequate resolution.
We looked for the following agonistic behaviors, first with binoculars and then in the video recordings: 1) Bite (one bird grasped the feather or the flesh of a neighbor with the beak), or Peck (one bird made a quick movement towards another with beak closed, often touching this individual), 2) Threat (one bird opened the beak widely with a movement of the head towards another individual, often with wings shaking), 3)Chase (one bird closely followed another), and 4) Displacement (one bird took the place of another bird that moved away).BORIS software(http://www.boris.unito.it/) was used to analyze the JVC camera video data, scoring how many times each bird won or lost an aggressive interaction (de Vries et al., 2006).Our objective was to collect 10–20 observations between each pair of individuals (Sanchez-Tojar et al.,2018), and we did this in approximately 6–9 days.
We also took data on affiliation, which was defined as two birds perched on the same 1 m long perch.We took this data opportunistically on visiting the aviary before feeding times and only recorded the species identity of the birds that shared perches and the distance between the birds (in cm, visually estimated).
2.5.Statistical analysis
David’s Score (DS) was used to analyze the dominance data.The proportion of wins by individualiin its interactions with another individualj(Pij) is the number of times thatidefeatsj(sij) divided by the total number of interactions betweeniandj(nij), i.e.Pij=sij/nij; the proportion of losses byiin its interactions withj(Pji) equals 1 -Pij(David, 1988).DS for each member,i, of a group is calculated with the formula:
(de Vries et al., 2006; Sanchez-Tojar et al., 2018), whereWrepresents the sum ofi’sPijvalues (i.e.,W=∑Pij(j=1, 2, …,N;j∕=i), andW2represents the summedWvalues of those individuals with whichiinteracted (multiplied by each individual’sPijvalues; i.e.,W2=∑WjPij(j=1, 2, …,N;j∕=i) (de Vries et al., 2006).Similarly,Lrepresents the sum ofi’sPjivalues (i.e.,L=∑Pji(j=1, 2, …,N;j∕=i), andL2represents the summedLvalues of its interactants (weighted by theirPjivalues;L2=∑LjPji(j=1, 2, …,N;j∕=i) (de Vries et al., 2006).To obtain a value standardized by group size, it is necessary to convert DS into a normalized DS (NormDS) as follows:
whereNis the number of individuals in the flock (de Vries et al., 2006).
We built mixed models to understand whether species differed in dominance from each other and what factors influenced dominance.Mixed models were constructed with the “lme4” package of the R statistical programming language.The first model asked whether species differed in dominance from each other; mixed-species flock was a random factor (i.e.which of the eight flocks was tested), and we analyzed only those species with three or more individuals.A second model was constructed to ask whether body mass and/or beak length influenced dominance, and here species and flock were random factors.A third model was built to determine whether the different sexes of David’s Fulvetta differed from each other in dominance or body mass,again with flock as a random factor.In the construction of these models,we sometimes found that the random factors did not explain any variance (i.e.the model was singular); we then removed the random factor and reran the model.
Finally, we looked at whether the difference in dominance status between species was related to affiliation.Species were ranked by their average dominance, and the difference in dominance was simply (the absolute value of) the difference in their ranked position.The metrics of affiliation were: (1) the proportion of all observations of affiliation which were composed of that species pair; (2) the average distance between the two species on the perches (in cm).Difference in dominance was related to these different metrics of affiliation using simple linear regression.The analysis was repeated using only the relationships between fulvettas and other species (here the difference in dominance[other species – fulvetta] could be either positive [fulvetta was more dominant] or negative [fulvetta was less dominant]).
In a follow-up analysis, we reran the models (specifically, the explanation of dominance by body mass and bill length, and the relationship between affiliation and dominance) without the five ground species that are unlikely to be long-term flock members.
3.Results
We captured eight mixed-species flocks, which included altogether 13 species and 55 individuals (the composition of each flock is given in Appendix Table S1).David’s Fulvettas were found in all flocks, and ranged from 1 to 4 individuals per flock (averaging 3 individuals).
3.1.Patterns of the dominance hierarchy
David’s Fulvetta was ranked 9 out of 13 species (there were eight species more dominant, and four less dominant), when using normalized David’s scores and averaging across individuals in a flock (see Table 1).The most dominant species was Streak-breasted Scimitar-Babbler(Pomatorhinus ruficollis;n= 7 individuals, 5 flocks).When the species were compared in their normalized David’s score, species differed in their dominance (F5,23.0=11.32,P<0.001; without one ground species,F4,21.40= 13.25,P< 0.001).Specifically, Streak-breasted Scimitar-Babbler was significantly more dominant than all other species (Tukey HSDP<0.004 for both analyses).
Body mass was the most important factor related to dominance.A preliminary model showed that flock explained no variance, so this random factor was removed from the model, leaving species as the only random factor.Models with two explanatory fixed factors (body massand beak length) showed that beak length did not explain further variance after body mass was accounted for (Type III Sum of Squares,F1,18.97= 1.34,P= 0.26; without ground species,F1,14.97= 0.66,P=0.43).A simplified model with just body mass showed that birds that were heavier were more dominant

Table 1The species caught in wild mixed-species bird flocks of southwestern Guangxi, and observed in the aviary.Species ordered from most to least dominant, as measured by the average normalized David’s ratio.For all metrics, we averaged among individuals of the same species within groups before averaging across groups.Standard deviations measure the variability among groups.
(Fig.1;F1,17.26= 9.83,P= 0.0059,R2conditional,marginal= 0.25, 0.48;without ground species,F1,9.97= 8.43,P= 0.016,R2conditional,marginal=0.28, 0.52).Although we did not include the number of individuals in the modeling, fulvettas had the most individuals in seven of eight flocks,yet were often subordinate, demonstrating that gregarious species were not necessarily dominant.
As for dominance within the fulvettas, the preliminary model showed that the random factor of flock explained no variance.We therefore simplified to a Pearsont-test, which showed that males (3.42± 0.53) had slightly higher dominance values than females (2.86 ±0.80), with the difference being marginally significant (Pearson’st-test;t22= 1.94,P= 0.065).However, the sexes were not different in their body mass (t22=0.94,P=0.36) or beak length (t22=0.62,P=0.54).
3.2.The relationship between dominance and affiliation
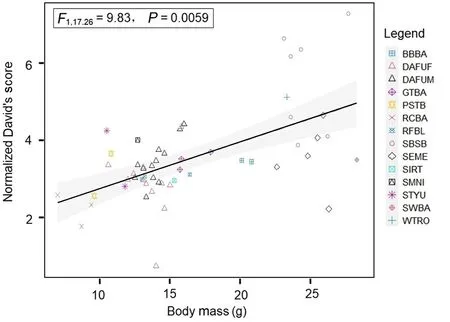
Fig.1.The relationship between dominance, as measured by the normalized David’s score, and body mass, for all birds in the experiments.Different species(see species abbreviations in Table 1) are given different colors and shapes;DAFUM = male David’s Fulvetta, and DAFUF = female David’s Fulvetta.The regression line and confidence intervals are created by a linear model, but the statistics are from a linear mixed model with species as a random factor.(For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
There was evidence of a negative relationship between a species pair’s difference in dominance status, and their affiliation, as measured by the proportion of all observations that were of the species pair (Fig.2;F1,30= 3.56;P= 0.069;R2= 0.11; without ground species,F1,15=10.28;P= 0.0059;R2= 0.41).That is, species pairs that were more separated in their dominance were less often seen together, particularly when ground species were not included in the analysis.However, the difference in dominance did not affect the closeness between the two species (F1,30=0.23,P=0.63,R2=0.01; without ground species,F1,15= 3.46,P= 0.083,R2= 0.19).When only looking at relationships between David’s Fulvetta and other species, there was no relationship between dominance and affiliation (for the proportion of observations:F1,8=1.98,P=0.20,R2=0.20; without ground species,F1,5=2.49,P= 0.18,R2= 0.33; for closeness,F1,8= 0.085,P= 0.78,R2= 0.01;without ground species,F1,5=0.92,P=0.38,R2=0.16).
4.Discussion
Through measurements in a controlled environment, we describe dominance in a mixed-species flock system in which the nuclear, leading species is relatively subordinate, ranking number 9 of 13 species (when the most dominant species is ranked number one).Here we will discuss first the factors that influenced dominance, and then how dominance may be related to the social organization of mixed-species flock systems,after acknowledging some limitations of the study.

Fig.2.Relationships between the difference in dominance between two species(i.e., the difference in “Rank” between two species in Table 1) and their affiliation in terms of the proportion of all affiliated individuals that was composed of that species pair (averaged across groups in which the species pair was found).In this analysis, all ground species were removed; the shaded region is a 95% confidence interval.
4.1.Limitations of the study
A first limitation of the study is the small sample size of mixedspecies flocks.The small number of flocks (n= 8) and the variance in composition between them resulted in four species being represented by a single individual, and there being only four species that were included in three or more flocks.Such small amounts of data may not be representative of the species involved in the system, but unfortunately our sampling, which occurred during the COVID epidemic, was very limited due to logistical constraints.
A second limitation is that the flocks that were captured are not completely representative of the flocks in nature at our study site.The birds were captured in ground-based mist nets, and therefore not all the individuals in the flocks were captured; in particular, species that forage in the midstory or canopy, such as tits, minivets, warblers and yuhinas were not captured.Even understory species could sometimes be missed(the flock with one individual David’s Fulvetta being an obvious example).Given that flocks in nature average 10.8 ± 5.0 individuals(Jiang et al., 2020) and our groups averaged 6.9 ±2.1 individuals, it is obvious that a sizeable fraction of flocks were not captured.In addition to these missing members, there may also have been some non-flock members that were captured in the same 25 m radius area as the flock.In particular, the five ground species mentioned in the methods are mostly solitary and territorial species that are unlikely to follow flocks for long periods, although they could interact with flock members when the flock comes through their territory (Goodale et al., 2020).Nevertheless, most of the species in the sample of captured birds are regular members of mixed flocks.For example, we captured five of the 11 most frequent species in the study of Jiang et al.(2020), all members of the laughing-thrushes (Leiothrichidae) or babbler (Timaliidae) families.Several of these species (e.g., Rusty-capped Babbler and Streak-breasted Scimitar-Babbler) are common species in flock systems of other parts of southern China, such as Hainan (Zou et al., 2011) and Guangdong (Zhang et al., 2013, 2020).Therefore, we think the results are fairly representative of the groups of babblers and their allies that interact in nature in flocks in the understory of the forests of the region.
An aviary experiment also, of course, provides the birds with an atypical environment, to which different individuals and different species may respond in varying ways, affecting the natural social system both within and between species.The distribution of food resources and risk of predation will be different in captivity compared to the wild, and both these factors influence flocking behavior (Terborgh, 1990; Gil et al., 2017).At the same time, in other experiments conducted with the same mixed-species flocks that focused on vocalizations, we found that the birds responded similarly to predators in the aviary as they do in the wild—for example, David’s Fulvetta was particularly vocal when elicited by predator models (Chen et al., in preparation).Others also have used aviary conditions to study dominance; for example, a recent study of ravens in captivity showed that their dominance relationships were similar to those in the wild, while at the same time the captive conditions allowed many tests of their cognitive abilities that would otherwise be impossible (Boucherie et al., 2022).While dominance interactions in the field could be influenced by the quality and quantity of the resources, by the earlier foraging success of the birds, and by the group of animals that is currently present, in the aviary we were able to always have the same individuals present, and, when measuring dominance,have the birds compete against each other for the same resources (the feeding bowl), while standardizing their foraging history (ad libitumfor much of the day).Nonetheless, we hope that future field observations of dominance in this system can be made to complement the findings of this aviary study.
4.2.Factors influencing dominance
Body mass (as a metric of body size) appeared to be the major influence on dominance, with David Fulvetta’s low ranking reflecting its status as a small bird (10.6–16.0 g from our data).Body mass is a common and powerful influence on dominance hierarchies in animal communities and in birds in particular (Shelley et al., 2004; French and Smith, 2005; Miller et al., 2017; Leighton et al., 2022).A secondary factor that can explain interspecific dominance in birds, once body size is accounted for, is beak length (Leighton et al., 2022), as beaks may be used as weapons.However, we found no evidence that beak length had an effect on dominance in this system.Future analyses might use more sophisticated measurements of beak strength, which could include beak depth as well as beak length (Soons et al., 2010).Finally, the number of individuals in groups is also known to affect intergroup conspecific aggression (Majolo et al., 2020), and could be important in birds, with numerous species being more dominant (Burger and Gochfeld, 1984;Shelley et al., 2004).However, again, we did not find evidence for this factor here, given that fulvettas were almost always the most numerous species.In mixed-species flocks, leading species that are gregarious like fulvettas, or tits in northern temperate systems (Dolby and Grubb,1998), are often subordinate to other larger, less gregarious species.
A final factor influencing dominance is sex, and we found female fulvettas tended to have lower rank than males (we did not test other species, because of their low numbers of individuals).Female David’s Fulvettas will be subordinate to most other species, whereas male David’s Fulvettas will be more similar to other species in the flock, and only subordinate to much larger birds (like Streak-breasted Scimitar Babbler,see Fig.1).The idea that some individuals of a species in a mixed-species flock will be closer in dominance to heterospecifics than others is a new realization both in empirical (Krams et al., 2020b) and theoretical(Coppinger et al., 2023) work.
4.3.Relationship of dominance to affiliation
We hypothesized that species pairs on the opposite end of the dominance hierarchy would not spend as much time together, because the subordinate would be threatened by the dominant.We found weak evidence in the direction of our hypothesis, as species with larger differences in dominance were less often found together.This relationship was more detectable when ground-foraging species were removed from the analysis, perhaps because the ground species did not use the perches as much as the other species, and therefore there was less data on their affiliation.We believe that this data is consistent with the idea that subordinate species will remove themselves from positions in which they are close to threatening dominant ones, and therefore also supports the idea that subordinate species could face some costs to flocking with dominant ones (Suhonen, 1993; Krams et al., 2020b).However, in the future it would be useful to make direct observations that demonstrate that subordinates move when dominants get too close.
4.4.The role of dominance on the organization of mixed-species flock systems
Our data is related to the question of how leader and follower species interact in flocks.David’s Fulvetta is a nuclear species in mixed flocks,and it is very vocal both in making alarm calls and in making mobbing calls, which other species can listen to (Jiang et al., 2020).It is possible that the benefits of participation to David’s Fulvetta in mixed-species flocks are less than the benefits to those species that follow them, as has been found in other systems (Dolby and Grubb, 1998; Hino, 1998;Sridhar et al., 2009).This then raises the question of why leaders continue to associate with flock followers.Our data provides one potential reason: leaders might simply be “trapped” and not able to escape followers if the followers are dominant to them.As mentioned above,the fulvetta flock system studied here is not unique in this respect; flock systems in North America consist of species of tits that are followed by larger nuthatches and woodpeckers, which listen to the tits and accrue benefits by reducing their own vigilance and increasing their foraging(Sullivan, 1985; Dolby and Grubb, 1998).Nevertheless, the “trapped leader” hypothesis is not the only one that could explain why leaders tend to continue being in flocks despite not benefiting a lot.Another hypothesis that also needs to be tested is that the costs of having heterospecific followers (e.g., aggression from dominant species) are not very high, or that the leaders do derive some benefits from their followers, such as a dilution of risk.We hope further work will test the“trapped leader” hypothesis and its alternatives.
In conclusion, we found that the leading species of a mixed-species flock group was among the more subordinate species, when dominance was measured under controlled conditions.Body size seemed to be more important than beak length or group size in influencing dominance.We found some evidence that dominance differences may shape the association of birds together in space.Differences in dominance between leader and followers in mixed-species flocks may influence their interactions, including the ways in which information about resources or predators are shared among flock participants.
Ethics statement
The present study complies with the current laws of China, and handling of birds was permitted by the Chongzuo Forestry Bureau(Permission No.2018 [1]) and the Animal Experimental Ethical Inspection of Guangxi University (Permission No.GXU2019-078).
Funding
This project was supported by the National Science Foundation of China, grant 31770424, and a Special Talents Recruitment Grant from Guangxi University.
Authors’contributions
Jichong Chen: Investigation, Formal analysis, Writing – Original draft preparation, Writing – Review and Editing.Estelle Meaux: Methodology, Writing- Review and Editing.Caiyun Li: Investigation.Aiwu Jiang: Supervision, Project administration, Writing – Review and Editing.Eben Goodale: Conceptualization, Funding acquisition; Formal analysis, Writing – Original draft preparation, Writing – Review and Editing.All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have influenced the work reported in this paper.
Acknowledgements
We appreciate the Chongzuo Forestry Bureau for permission to catch the birds, and the Animal Experimental Ethical Inspection of Guangxi University for permission to keep the birds in captivity and conduct the experiments.We are grateful for the help of Shilong Liu, Tianlong Zhou,and Haijun Tan in the field and Wuyuan Pan with the aviary.Two anonymous reviewers provided constructive comments that improved the manuscript.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.avrs.2023.100139.

- Avian Research的其它文章
- Home range variability and philopatry in Cinereous vultures (Aegypius monachus) breeding in Iberia
- A high level of extra-pair paternity in the Chestnut Thrush(Turdus rubrocanus)
- Comparisons of microstructure and elemental composition of eggshells among wild plover populations
- Antipredatory call behavior of lapwing species in an Afrotropical environment
- Impact of agricultural landscape structure on the patterns of bird species diversity at a regional scale
- Modelling the nesting-habitat of threatened vulture species in the caucasus:An ecosystem approach to formalising environmental factors in species distribution models