
核桃林下环境因子对滇黄精光合、产量及品质的影响
2024-01-20 02:46:26肖良俊武建林肖靖秀
西北林学院学报 2024年1期
肖良俊,武建林,肖靖秀
(1.云南省林业和草原科学院,云南 昆明 650201;2.云南农业大学,云南 昆明 650201)
核桃(Juglansspp.)是云南省“十四五”“绿色食品牌”重点打造的坚果产业之一,滇黄精(Polygonatumkingianum)为云南省地道药材。在核桃种植的坝区和平缓山地拥有发展林下中药材的巨大空间。滇黄精为百合科(Liliaceae)黄精属(Polygonatum)多年生药食两用草本植物,其块茎主要含浸出物、黄精多糖、黄精皂苷等物质,具有补气、养阴、润肺等作用[1]。近年随着野生资源不断减少,发展人工种植是保护滇黄精野生资源和产业持续发展的必由之路。核桃为喜阳树种,而滇黄精喜荫,适宜于核桃林下半野生化栽培。
林下光环境因子包括光照强度、温度和湿度等,它影响林下植物的光合作用、生长发育和生理生化等[2-4]。不同树龄和林木冠层结构对林下的空气和温湿度产生较大的影响,且不同季节产生的影响也不同[5]。林药复合系统作为一种特殊的栽培模式,其林下作物处于遮荫环境,光照对作物生长发育中有明显的限制作用[6]。林木冠层的光拦截作用改变了入射光质量或降低林下作物的光合有效辐射,从而导致林下作物光合作用降低、产量减少的现象普遍存在[7]。王若伦等[7]在“核桃-冬凌草复合系统内光合有效辐射对冬凌草生理生长的影响”中发现,PAR和冬凌草Pn、Tr、株高、产量以及甲素含量均显著相关,说明树冠遮荫会降低复合系统内PAR,影响冬凌草的光合作用,进而影响冬凌草的品质[8]。欧亚丽等[9]、贺安娜等[10]研究发现,遮荫条件可以降低直射光对黄精的光抑制强度,降低黄精的蒸腾速率,提高其水分利用率,黄精净光合速率日变化曲线由露天栽培下的双峰曲线变为单峰曲线,遮荫条件下的黄精光合速率变化呈单峰曲线。
在林药复合系统中开展植物光能竞争及光合生理特性方面的研究有助于了解和掌握植物的生存策略及植物对环境资源的利用效率,同时对于林药复合系统中树种的选择和配置、种间互作与生产力的关系,以及林药复合系统的可持续经营具有十分重要的意义。本研究以不同树龄核桃林下滇黄精为对象,测定其林下环境因子、滇黄精光合指标、产量和品质,并对试验结果进行相关性和通径分析。揭示不同树龄核桃林下环境因子及滇黄精光能利用特征,明确其对滇黄精生长、产量和品质的影响,为核桃滇黄精合理间作提供依据。
1 材料与方法
1.1 试验地概况
试验地点为云南林业和草业科学院核桃种质资源圃,位于昆明市盘龙区昆明树木园内(25°08′38″N,102°44′35″E),属北纬低纬度亚热带高原山地季风气候,海拔2 050 m,年平均气温14.7 ℃,绝对最低温-7.8 ℃,绝对最高温31.5 ℃,≥10 ℃的积温4 799.3 ℃,平均霜期85 d,年降水量1 100 mm,相对湿度72%,全年日照时数>2 249.3 h。土壤pH 5.49、有机质19.65 g·kg-1、全N 1.92 g·kg-1、水解N 48.62 mg·kg-1、全P 3.32 g·kg-1、全K 8.44 g·kg-1、速效K 53.83 mg·kg-1。
1.2 试验材料
核桃品种为“漾濞泡核桃”,是云南早期无性系优良品种。滇黄精品种为“林韵1号”优良无性系。
1.3 试验样地设置
在核桃-滇黄精复合经营区内分别选择立地条件一致的6、11、20 a核桃林地,核桃树株行距为5 m×6 m,选择9株长势良好、株型均匀的核桃树,每个树龄3棵组成1个样地即1个处理,重复3次,每样地面积为100 m2,CK为只种滇黄精、无核桃树的裸露地块。2020年12月种植滇黄精块茎,块茎大小均匀,质量12 g左右,滇黄精株行距为25 cm×30 cm,各试验处理的管理措施均一致。
1.4 试验方法
于2021年5-9月每月中旬各测定1次。选择晴朗无云的天气,利用Li-6400便携式光合测试仪在核桃树冠下东西南北4个方向的内部(距离核桃主干10 cm)、中部和外缘随机选择3株,每株选取2片中上部位同一位置、成熟无病虫害的叶片,分别在滇黄精生育期5月(伸长期)、6月(开花期)、7月(结实期)、9月(果熟期)进行光合日变化及林下环境因子测定。按相同方法对CK地块内、中、外部的滇黄精进行测定。8:00-18:00每隔2 h测定1次,共计测量6次,时间段分别为8:00、10:00、12:00、14:00、16:00、18:00。测定指标包括:净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)、滇黄精叶片温度(Tleaf)、光合有效辐射(PAR)、相对湿度(RH)和大气CO2浓度,对每片叶测定5次,测定数据取平均值[11]。
11月中旬,在滇黄精植物完全倒苗后,将每处理的滇黄精块茎挖出称重,计算产量,每处理随机选取5株,洗净块茎晾干,称鲜重后切片,60 ℃烘干后称量干重,根据公式(干重/鲜重)×100%计算折干率,利用粉碎机将其磨碎过3号筛(50目),按照蒽酮-硫酸法进行多糖含量测定[12]。
1.5 数据处理
使用Excel 2010进行数据整理及统计分析,使用SPSS 21.0进行方差分析和相关性分析、通径分析、冗余分析,使用Excel 2010制图。
2 结果与分析
2.1 不同树龄核桃树林下环境因子
由表1可知,5-9月,叶温均表现为CK>6 a>11 a>20 a,且均存在显著差异;空气CO2浓度均无显著差异。空气相对湿度表现为:5月,20 a显著高于11 a,11 a显著高于6 a和CK,且6 a和CK无显著差异;6月,20 a和11 a显著高于6 a,6 a显著高于CK;7月,20 a显著高于11 a,11 a显著高于6 a,6 a显著高于CK;9月,20 a和11 a显著高于CK,与6 a无显著差异,且6 a和CK无显著差异。光合有效辐射表现为:5月和6月CK显著高于6 a,6 a显著高于11 a和20 a,11 a和20 a无显著差异;6月,20 a显著低于11 a,11 a显著低于6 a,6 a显著低于CK;7月,CK显著高于6 a和11 a,6 a和11 a显著高于20 a,且6 a和11 a无显著差异。

表1 不同树龄核桃树林下因子Table 1 Environmental factors in the understory of walnut trees of different ages
2.2 不同树龄核桃树林下滇黄精光合特征
2.2.1 净光合速率(Pn)日变化 由图1可知,整个生育期CK和不同处理的滇黄精Pn日变化在不同生育期存在差异,6、11、20 a核桃林下的滇黄精的Pn日变化在整个生育期均呈双峰曲线,第1个峰值均出现在10:00,第2个峰值出现时间有所区别,5月第2个峰值均出现在14:00;6、7月6、11 a和20 a的第2个峰值均出现在16:00;9月6 a下的滇黄精的Pn第2个峰值出现在14:00,11 a和20 a的滇黄精Pn的第2个高峰均出现16:00;不同树龄核桃林下滇黄精不同时刻Pn的差异不同,5月整体表现为11 a>6 a>20 a>CK;6月整体表现为11 a>6 a>20 a>CK;7月整体表现为6 a>CK>11 a>20 a;9月整体表现为CK>11 a>6 a>20 a;全生育期整体表现为11 a>6 a>CK>20 a。

不同字母表示同一月份不同树龄间差异显著(P<0.05)。下同。
2.2.2 气孔导度(Gs)日变化 由图2可知,6 a和11 a的滇黄精Gs在整个生育期日变化均呈双峰曲线,均在10:00达到第1个高峰,5、6月6 a于16:00达到第2个高峰,7、9月6 a的于14:00达到第2个高峰;5月11 a的于14:00达到第2个高峰,6、7月于16:00达到第2个高峰;5、7月20 a滇黄精的Gs日变化呈单峰曲线,于12:00左右达到最高,6、9月20 a滇黄精的Gs日变化均呈双峰曲线,均于10:00达到第1个高峰,于16:00达到第2个高峰;5、6、7月CK的滇黄精Gs日变化呈单峰曲线,5月于16:00左右达到最高,6月于12:00左右达到最高,7月于10:00左右达到最高,9月CK的滇黄精Gs日变化呈双峰曲线,于10:00达到第1个高峰,16:00达到第2个高峰。不同月份不同树龄核桃树下滇黄精不同时刻Gs的差异不同。5月整体表现为11 a>6 a>20 a>CK;6月整体表现为11 a>6 a>20 a>CK;7月整体表现为6 a>CK>11 a>20 a;9月整体表现为CK>6 a>11 a>20 a;全生育期整体表现为11 a≥6 a≥CK>20 a。

图2 滇黄精气孔导度日变化Fig.2 Daily variation of stomatal conductance of P.kingianum
2.2.3 胞间CO2浓度(Ci)日变化 由图3可知,整个生育期CK和不同处理的滇黄精Ci日变化呈现先下降后上升的低谷趋势。5月,6 a和11 a林下的滇黄精Ci最低点均出现在16:00,20 a和CK的滇黄精Ci的最低点均出现在14:00;6月,6、20 a和CK的滇黄精胞Ci最低点均出现在14:00,11 a滇黄精Ci的最低点均出现在16:00。7月,11、20 a和CK的滇黄精Ci最低点均出现在14:00,6 a下的滇黄精Ci最低点均出现在16:00;9月,6、11、20 a和CK的滇黄精Ci最低点均出现在16:00。不同月份不同核桃树龄林下滇黄精不同时刻Ci的差异不同。5月整体表现为11 a>6 a>20 a>CK;6月整体表现为20 a>6 a>11 a>CK;7月整体表现为20 a>6 a>11 a>CK;9月整体表现为20>6 a>CK>11 a。全生育期整体表现为CK≥6 a≥20 a>11 a。
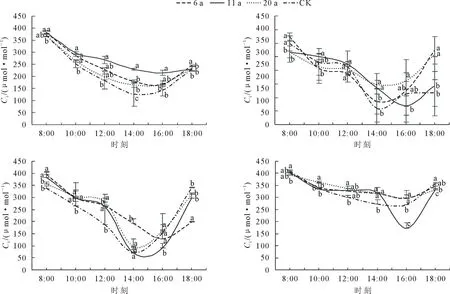
图3 滇黄精胞间CO2浓度日变化Fig.3 Daily variation of intercellular CO2 concentration of P.kingianum
2.2.4 蒸腾速率(Tr)日变化 由图4可知,6 a下的滇黄精Tr日变化在5、6、9月均呈双峰曲线,均在10:00达到第1个高峰,5月于16:00达到第2个高峰,6月于12:00达到第1个高峰,9月于14:00达到第2个高峰;在7月,6 a下的滇黄精Tr日变化呈单峰曲线,于14:00达到最大值。11 a下的滇黄精Tr日变化在5、6、7月和9月均呈双峰曲线,10:00达到第1个高峰,5月和6月于16:00达到第2个高峰,7月和9月于14:00达到第2个高峰。5、7月和9月,20 a林下的滇黄精Tr日变化呈单峰曲线,最大值出现时间分别为12:00、14:00和10:00;20 a林下的滇黄精Tr日变化在6月呈双峰曲线,于12:00达到第1个高峰,在16:00达到第2个高峰。5月CK的滇黄精Tr日变化呈双峰曲线,在10:00达到第1个高峰,在16:00达到第2个高峰,6月和7月CK的滇黄精Tr日变化呈单峰曲线,最大值出现时间分别为12:00和10:00;CK的滇黄精Tr日变化在9月呈双峰曲线,于10:00达到第1个高峰,于16:00达到第2个高峰。不同月份不同树龄核桃树下滇黄精不同时刻Tr的差异不同。5月整体表现为11 a>6 a>20 a>CK;6月整体表现为CK>11 a>20 a>6 a;7月整体表现为CK>11 a>6 a>20;9月整体表现为CK>6 a>11 a>20。全生育期整体表现为CK>11 a>6 a>20 a。

图4 滇黄精蒸腾速率日变化Fig.4 Daily variation of transpiration rate of P.kingianum
2.3 不同树龄核桃树林下滇黄精产量和品质
由图5可知,滇黄精的产量差异表现为11 a>6 a>20 a>CK,处理间差异显著,20 a比CK提高了216%,6 a比20 a提高了87.6%,11 a比6 a提高了33.4%。滇黄精的块茎多糖含量差异表现为11 a的显著高于6、20 a和CK,且6、20 a和CK间均无显著差异,11 a比20 a和CK提高了14.5%。
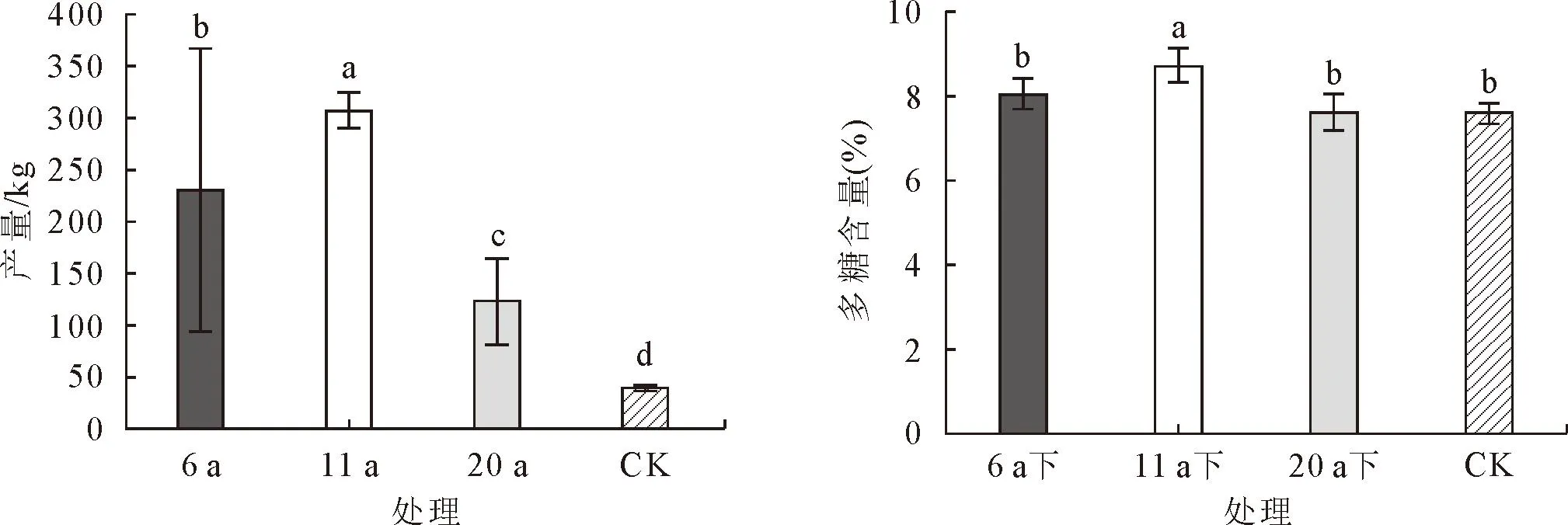
图5 不同树龄核桃树下滇黄精产量、块茎折干率和多糖含量差异Fig.5 Differences in the yield,tuber drying rate and polysaccharide content of P.kingianum unde different age walnut plantations
2.4 林下环境因子对滇黄精光合特性的影响
由表2可知,5月滇黄精Pn与叶温、空气相对湿度呈显著正相关,相关系数分别为0.707、0.652,与光合有效辐射呈极显著负相关;滇黄精Gs和Tr与光合有效辐射均呈极显著负相关,相关系数为0.840、0.861。6月滇黄精光合参数与环境因子均为显著相关关系。7月滇黄精Pn与叶温呈显著正相关,相关系数为0.687,和空气相对湿度、光合有效辐射呈显著正相关,相关系数为0.611、0.712;滇黄精Gs和Tr与空气CO2浓度均呈显著负相关,相关系数为0.594、0.680,滇黄精Ci与光合有效辐射呈显著负相关,相关系数为0.660。9月滇黄精Pn与叶温、光合有效辐射均呈极显著正相关,相关系数为0.870、0.954,与空气相对湿度呈显著负相关,相关系数为0.669;滇黄精Gs与叶温、光合有效辐射均呈显著正相关,相关系数为0.820、0.810,与空气相对湿度呈极显著负相关,相关系数为0.751;滇黄精Tr与叶温、光合有效辐射均呈极显著正相关,相关系数为0.942、0.917。在云南一般年份5月为旱季,6月中下旬进入雨季,旱季天气晴朗光合有效辐射系数大,较大的光合有效辐射抑制滇黄精的净光合速率,即显负相关。7、9月进入雨季光合有效辐射系较小有利于滇黄精净光合速率,即显正相关。

表2 林下环境因子与滇黄精光合指标间的相关性Table 2 Correlation between environmental factors in the forest understory and photosynthetic parameters of P.kingianum
2.5 林下滇黄精光合特性对其产量品质的影响
由表3可知,5-6月,滇黄精Pn与其产量均呈极显著正相关,相关系数分别为0.777、0.887,与多糖含量在5月呈显著正相关,相关系数为0.640,在6月呈极显著正相关,相关系数为0.910,其他月份与产量和多糖含量均无显著相关关系;滇黄精Gs在5月与产量和多糖含量呈极显著正相关,相关系数分别为0.860、0.792;滇黄精Ci与产量和多糖含量在5月均呈显著正相关,相关系数分别为0.745、0.859;滇黄精Tr在5月与产量和多糖含量均呈极显著正相关,相关系数分别为0.887、0.762。
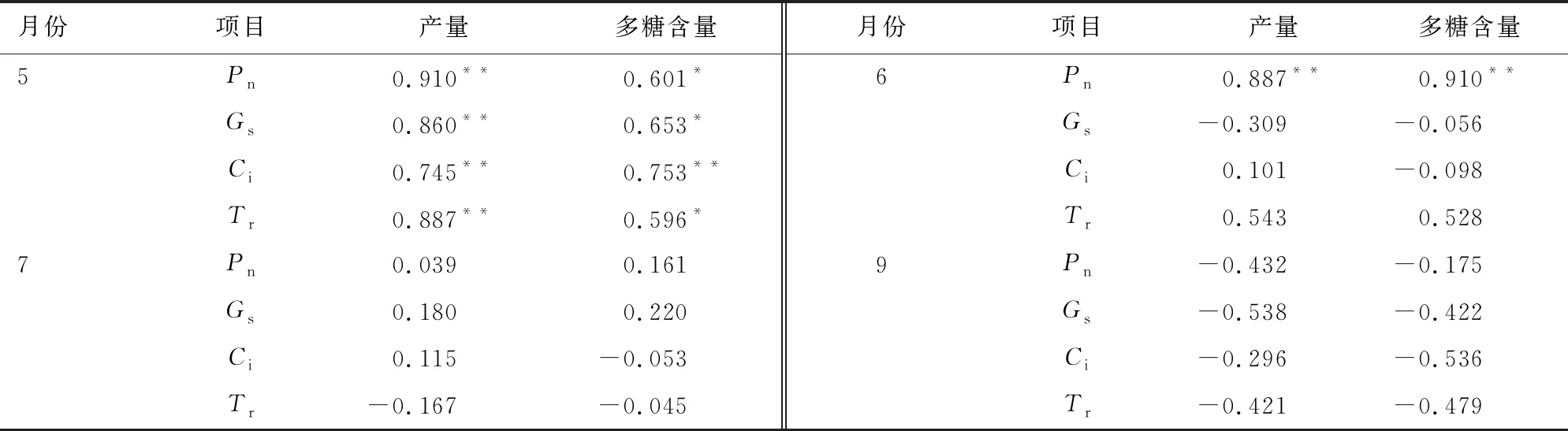
表3 滇黄精光合指标与滇黄精农艺性状、产量、品质间的相关性Table 3 Correlations between photosynthetic characteristics yield and quality of P.kingianum
对5月的滇黄精光合指标与产量进行通径分析(表4)表明,R2为0.902,光合指标可以解释滇黄精产量变化的90.2%,Pn对滇黄精的产量直接贡献最大,其次为Ci、Gs,直接贡献值均为正值,间接贡献最小。Tr的直接贡献最小,且直接贡献值为负值,间接贡献值最大,其中Pn对产量的贡献最大。决策系数表明,Pn的决策系数大于其余光合指标的决策系数,Pn对滇黄精产量起主导作用,Ci次之。
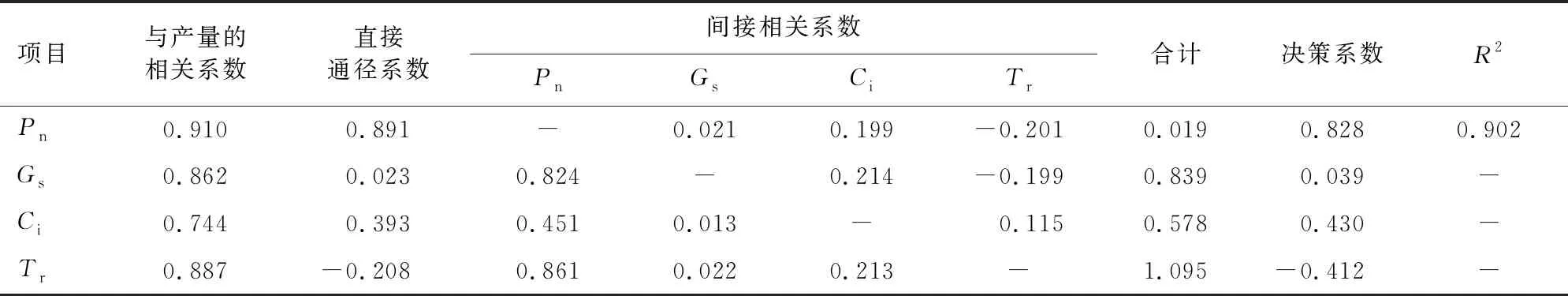
表4 滇黄精光合指标与产量的通径分析Table 4 Path analysis of photosynthetic index yield of P.kingianum
对5月的滇黄精光合指标与品质(多糖含量)进行通径分析(表5)表明,R2为0.510,光合指标仅可以解释滇黄精多糖含量变化的51.0%,还有49%的其他因子对多糖含量有影响,Tr对滇黄精的多糖含量直接贡献最大,直接贡献值为正值,其次为Gs、Ci、Pn。Pn的直接贡献最小,且直接贡献值为正值。间接贡献值最大的为Tr,其中通过Gs对产量的贡献最大。决策系数表明,Tr的决策系数大于其余光合指标的决策系数,Tr对滇黄精多糖含量起主导作用,Ci次之。
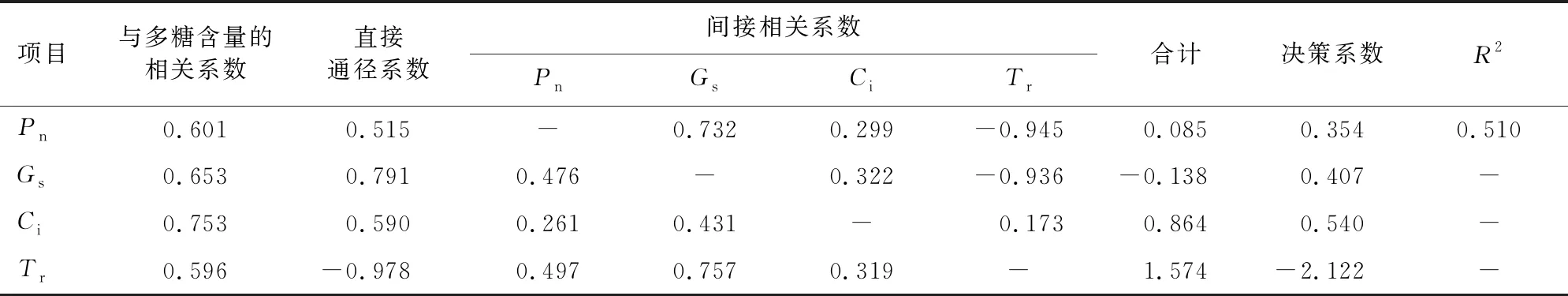
表5 滇黄精光合指标与品质的通径分析Table 5 Path analysis of photosynthetic index quality of P.kingianum
3 讨论
由于农林复合系统内的水肥资源可以通过人工操作进行及时有效的补充,因此林下环境因子被认为是农林复合系统中对间作物影响最大的因子之一。不同核桃树龄冠层结构的不同改变了林内光照(光强和光质)、湿度、温度等,进而调节林下小气候,对林下作物的光合作用速率产生影响,影响着林下作物的生长及发育[13]。对不同核桃树龄林下环境因子进行测定,结果显示不同树龄核桃林下环境因子中的光合有效辐射、叶温和空气湿度在不同月份均存在显著差异,而空气CO2浓度无差异,比较稳定。
林下环境因子(光合有效辐射、空气湿度、温度、CO2浓度等)的差异,影响着林下植物的光合作用效率[14-15]。滇黄精为阴性植物,本研究发现环境因子对滇黄精光合影响较大,林下滇黄精的不同生育期光合特性存在差异,净光合速率日变化呈双峰曲线,胞间CO2浓度日变化呈“V”形曲线,气孔导度和胞间CO2浓度日变化曲线不规则,裸露地块种植滇黄精(CK)的净光合速率呈单峰曲线。林下环境因子与滇黄精光合指标间的相关性分析显示,不同生育期林下滇黄精光合速率的相关因子主要为光合有效辐射、叶温和空气相对湿度。
不同树龄核桃树冠幅大小和结构不同,同一月份不同树龄其林下环境因子的光合有效辐射、空气相对湿度、叶温差异显著,进而影响着林下滇黄精的净光合速率,滇黄精全生育期净光合速率整体表现为11 a>6 a>CK>20 a。试验结果表明,滇黄精的光合作用对林下环境因子的响应呈现为较强的光合有效辐射(CK)和过低的光合有效辐射(20 a),均不利于光合作用。林下滇黄精光合特性对其产量、品质的相关性和通径分析表明,5-6月(伸长期-开花期)滇黄精的净光合速率与其产量和品质显著正相关,8-10月(结实期-枯萎期)株高、地径与其产量和品质呈显著正相关,该结果与实际动态测定滇黄精产量和品质分析结果相符合。5-6月,滇黄精较高的净光合速率产生的物质均被营养生长所消耗,即株高的生长和地径的增粗,高壮的植株有利于8-10月块茎的增长和药用成分的提高,本研究发现产量与品质呈显著正相关,高产即高品质。
4 结论
综合以上研究得出,不同核桃树龄的冠层结构不同,影响着林下环境因子,且不同时刻、月份的环境因子各不相同。在核桃滇黄精间作模式中,核桃树主要通过其冠层结构来影响光合有效辐射、叶温空气相对湿度等环境因子,进而调控林下滇黄精不同生育期的净光合速率等指标,最终影响滇黄精的生长、产量和品质。本试验中11 a核桃树冠下的环境因子最利于滇黄精光合作用,利于滇黄精生长、产量最高和品质最优。在生产实践中,可根据不同作物对林下环境因子需求的差异,选择与林下环境因子相适应的间作物,或通过树冠的整形修剪来调节林下环境因子,以提高土地生产力和光能利用率。
猜你喜欢
今日农业(2021年8期)2021-07-28 05:56:04
Digital Chinese Medicine(2020年3期)2020-11-03 03:12:12
云南农业科技(2020年5期)2020-10-14 07:24:50
书香两岸(2020年3期)2020-06-29 12:33:45
新疆农业科学(2019年11期)2019-12-31 07:02:30
天然产物研究与开发(2016年11期)2016-06-15 20:29:17
卫星与网络(2016年12期)2016-02-05 09:23:22
小天使·四年级语数英综合(2015年6期)2015-07-17 18:01:11
药学与临床研究(2015年4期)2015-06-05 11:35:51
饮食科学(2009年3期)2009-03-23 10:10:06
