
冀北山地不同坡向白桦枝叶异速生长关系研究
2024-01-20 02:46:10许竹锐王世虎高青青于小萌徐学华
西北林学院学报 2024年1期
许竹锐,王世虎,高青青,于小萌,徐学华
(河北农业大学 林学院,河北 保定 071000)
植物的枝叶大小关系是植物生活史策略研究中不可或缺的一部分,尤其是最具有活力的当年生枝条,对于解释植物生理生态变化来说更为重要[1]。当年生枝条即小枝作为木本植物的独立构件单元,在营养物质、水和碳水化合物方面相比老枝具有更大的流动性[2]。小枝主要由叶和茎组成,叶由叶片和叶柄组成,是树枝的一个基本组成部分[3]。叶片是光合作用和碳水化合物合成的主要场所,而叶柄是一个悬臂结构,支持和供应叶片,此外还发挥着支持静态重力和抵抗外部动态张力的作用[4]。植物的枝叶大小关系通常采用异速生长模型来解释,异速生长是指生物体某2个性状的相对生长速率不同的现象[5-6],认为植物器官关系可用幂函数Y1=βY2α表示,当α=1时,模型反映的是等速生长关系;当α≠1时,模型反映的是异速生长关系[7]。异速生长模型与枝叶关系相结合,有助于量化物种间变异差异和更好理解植物性状经济谱的投资与收益策略[8]。
植物当年生枝叶异速生长研究通常集中在叶与枝的大小权衡和叶大小与数量权衡等方面,已有多名学者开展研究,Westoby等[9]以悉尼33个木本植物的小枝为研究对象,发现小枝的横截面积与其所支持的叶面积为异速关系。Wang等[10]以不同海拔常绿阔叶植物为研究对象,结果表明随着海拔的升高,叶质量与小枝质量之间呈等速生长关系。叶大小-数量是植物小枝内部资源分配的一个重要权衡关系,其权衡关系受到物种本身的遗传特性以及所处环境的影响[11]。Yang等[12]发现温带阔叶树种其叶大小与出叶强度之间呈负等速生长关系。孙俊等[8]对不同海拔的竹种研究时发现,其毛竹叶大小与出叶强度之间为负等速生长关系,且不随海拔发生变化;而肿节少穗竹在低、高海拔呈负异速生长,在中海拔呈负等速生长关系。由此看出,植物枝叶生长关系在不同的环境条件表现不同,小枝之间的权衡关系在不同环境条件下如何变化还有待进一步探索。
白桦(Betulaplatyphylla),为桦木科(Betulaceae)桦木属(Betula)落叶乔木,喜光,耐严寒,耐瘠薄,生长较快,适应性强,分布甚广,是天然次生林生态系统演替过程的先锋树种,对于维持区域生态平衡具有重要的意义[13-14]。目前已有学者围绕植物当年生小枝异速生长关系进行了研究,但大多是针对林内多个物种,而很少对生境梯度下的单一物种生长关系进行研究[15]。尤其不同坡向白桦小枝性状间的异速生长关系的变化规律尚不明确。坡向作为重要的地形因子之一,改变了不同坡向间的温度和光照,这些变化是否会影响白桦小枝的生物量分配,是否会对白桦枝叶性状的异速生长关系产生影响,本研究以冀北山地天然次生林中的优势物种白桦为对象,分析白桦小枝性状随坡向变化的规律,旨在揭示坡向对白桦小枝生长的影响,从而阐明白桦在不同坡向条件下的生态策略和适应性,为进一步探讨白桦对该天然次生林稳定性的维持机制与结构和功能的影响提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于河北省承德市围场县五道沟林场(41°52′01″-42°11′08″N,116°49′20″-116°59′04″E),与内蒙古自治区相邻。海拔1 000~1 880 m。属半湿润半干旱大陆性季风气候,年均气温-1.4 ℃~4.7 ℃,年均降水量380~560 mm,土壤以褐色土和棕壤土为主。植被类型是以白桦、黑桦(B.dahurica)和蒙古栎(Quercusmongolica)为主的天然次生林,山杏(Prunussibirica)和绣线菊(Spiraeasalicifolia)等组成天然灌丛,还有华北落叶松(Larixgmelinii)人工林和油松(Pinustabuliformis)人工林。白桦为该研究区的代表树种,分布较为广泛,主要分布在半阳坡、半阴坡和阴坡等,生长状况良好,发挥着重要的生态功能。
1.2 样地设置与样品采集
在对研究区天然次生林充分调查的基础上,于2021年7-8月,选取海拔、坡度等条件基本一致的半阳坡、半阴坡、阴坡的白桦(表1)。每个坡向设置3个20 m×30 m的样地,对样地内的白桦进行调查。计算其平均胸径,在每个样地内选择胸径大小与平均胸径基本一致的3株,使用高枝剪在白桦的4个方向剪取小枝,每株树木选取6个没有病虫害的当年生末端小枝,将其带回实验进行测定。
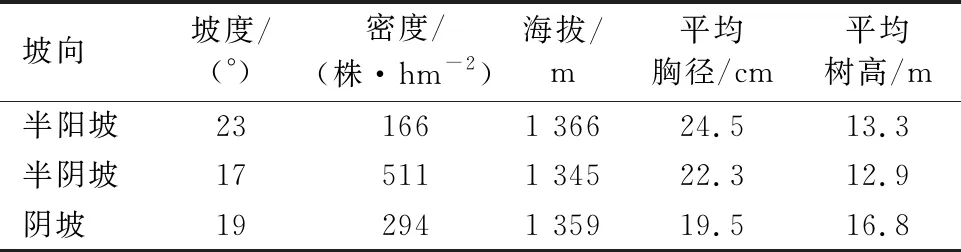
表1 不同坡向白桦样地基本概况Table 1 Basic situation of B.platyphylla sample plots in different slope directions
1.3 枝叶性状指标测定
当年生末端小枝分为叶片、叶柄和茎3个部分,对其进行分离,记录小枝上叶片数量。首先使用扫描仪 EXPRESSION 10000XL对叶片进行扫描,获得叶面积。随后样品放入65 ℃的烘箱中烘干至恒重,称其每个部位的干质量。小枝干质量、出叶强度、单叶片干质量、单叶片面积和单叶柄干质量,由以下公式所得:
小枝干质量=总叶片干质量+总叶柄干质量+茎干质量
(1)
出叶强度=小枝上叶片数量/小枝干质量
(2)
单叶片干质量=总叶片干质量/小枝上叶片数量
(3)
单叶片面积=总叶片面积/小枝上叶片数量
(4)
单叶柄干质量=总叶柄干质量/小枝上叶片数量
(5)
1.4 数据处理
使用SPSS 19.0对数据进行单因素方差分析和多重比较,分析其在不同坡向间的差异。枝叶关系采用异速生长方程Y1=βY2α,该方程经过对数转换后,可线性化为lgY1=lgβ+αlgY2,lgβ为异速生长常数,即线性关系的截距;α为异速生长指数,即线性关系的斜率,当异速生长指数α与1或-1差异显著时,表明两性状间为异速生长关系;差异不显著时,为等速生长关系[7]。采用R 4.12 "smatr"包中标准化主轴回归分析(standardized major axis estimation,SMA)的方法计算异速生长指数和常数,并对异速生长指数进行异质性检验,且在同质时计算共同斜率。
2 结果与分析
2.1 坡向对白桦小枝性状的影响
白桦在不同坡向间茎干质量为0.114 1~0.176 8 g、总叶片干质量为0.348 2~0.456 7 g,总叶柄干质量为0.028 4~0.037 4 g,其中茎干质量在半阳坡和阴坡显著大于半阴坡,总叶片干质量和总叶柄干质量在半阳坡显著高于阴坡,但与半阴坡无显著差异,且半阴坡与阴坡也无显著差异(表2);小枝干质量、单叶片干质量和单叶柄干质量在坡向间分别为0.528 1~0.670 9、0.058 8~0.082 9 g和0.004 7~0.006 7 g,其在坡向间表现为半阳坡高于半阴坡和阴坡,而半阴坡和阴坡无显著差异;白桦总叶面积、单叶片面积、叶数在坡向间没有显著差异;出叶强度则表现为阴坡和半阴坡显著高于半阳坡的规律。
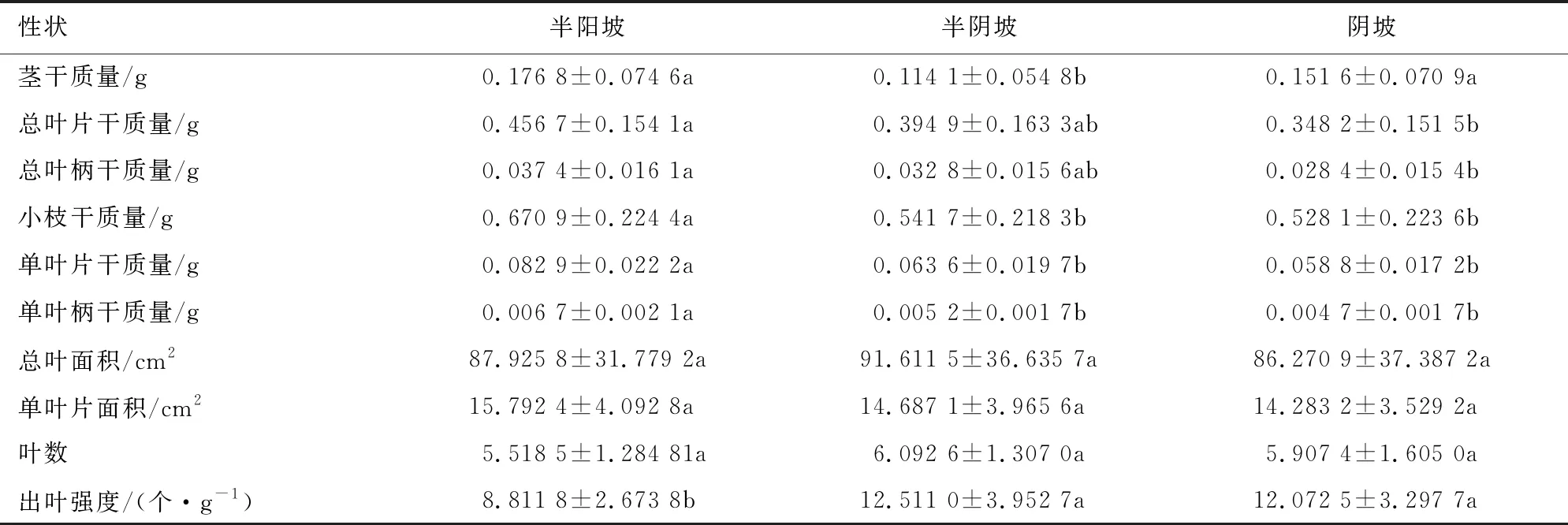
表2 白桦枝叶性状Table 2 Twig and leaf traits of B.platyphylla
2.2 坡向对白桦小枝生长的影响
整体上,茎干质量与小枝干质量、总叶片干质量与小枝干质量、总叶柄干质量与小枝干质量、总叶片干质量与总叶柄干质量、总叶片面积与总叶柄干质量均呈显著正相关关系。在小枝水平上,茎干质量与小枝干质量、总叶片干质量与小枝干质量、总叶片面积与小枝干质量的异速生长指数与1.0无显著差异,呈等速生长关系(表3),且在不同坡向间拥有共同异速生长指数,分别为1.12、1.04、1.04(图1A、1B、1D);总叶柄干质量与小枝干质量的异速生长指数与1.0有显著差异,呈异速生长,异速生长指数从半阳坡到阴坡分别为1.30、1.12、1.27(表3),且在不同坡向具有共同斜率1.22(图1C)。

图1 白桦小枝异速生长关系Fig.1 Allometry relationship of twig of B.platyphylla

表3 不同坡向白桦枝叶的SMA分析Table 3 Relationship of twig and leaf analyzed by SMA in different aspects
在叶水平上,总叶片干质量、总叶片面积与总叶柄干质量在不同坡向间的共同异速生长指数分别为0.85和0.86(图2A、2B);其中半阴坡总叶片干质量与总叶柄干质量的异速生长指数与1.0无显著差异,呈等速生长,而半阳坡和阴坡异速生长指数与1.0有显著差异,呈异速生长(表3);阴坡总叶片面积与总叶柄干质量的异速生长指数与1.0无显著差异,呈等速生长,而半阴坡和半阳坡呈异速生长(表3)。
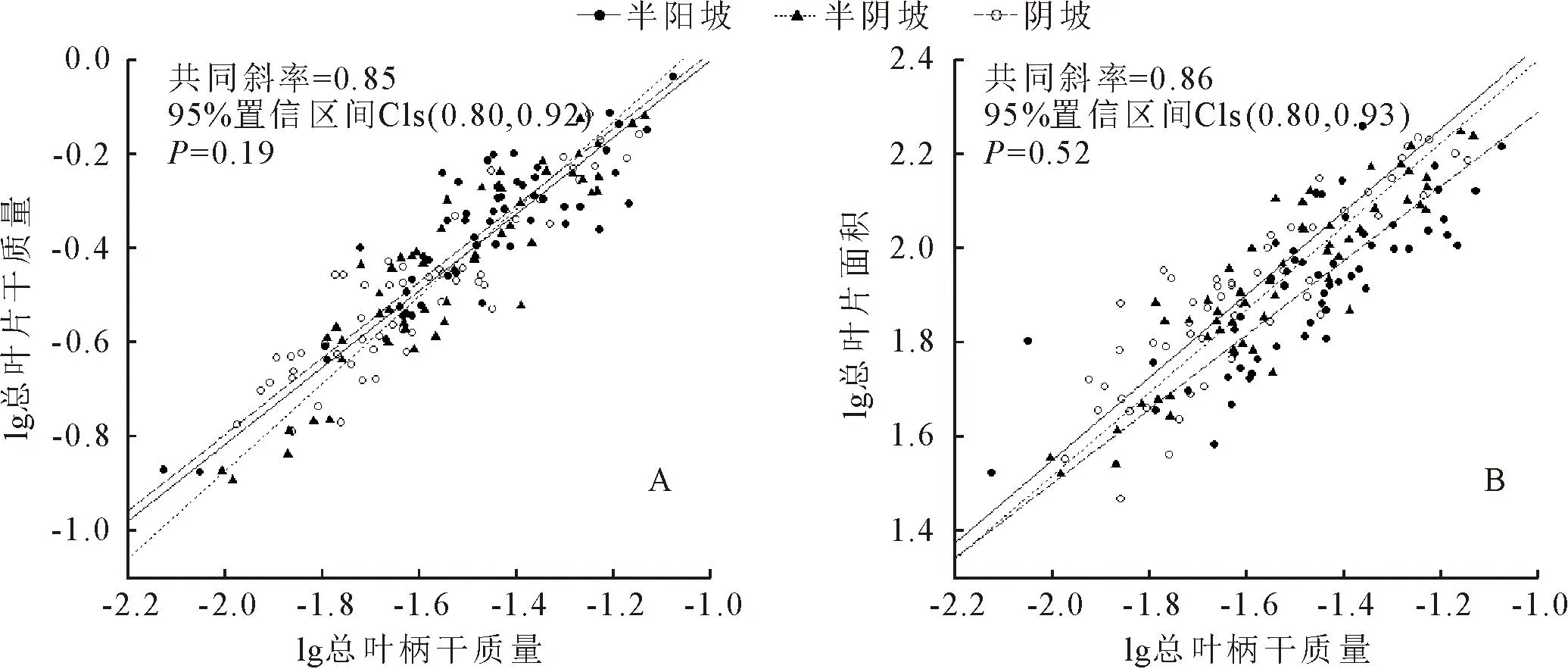
图2 白桦叶异速生长关系Fig.2 Allometry relationship of leaf of B.platyphylla
2.3 坡向对白桦叶片大小与数量的影响
白桦单叶干质量与出叶强度的异速生长指数在半阳坡、半阴坡、阴坡分别为-1.07、-1.07、-1.00,与-1.0无显著差异(表3),且在不同坡向间有共同异速生长指数-1.05(图3A),表明二者之间存在负等速生长关系且不随坡向发生改变,但其阴坡异速生长常数小于半阴坡和半阳坡,即在共同斜率下,出叶强度一定时,其阴坡的单叶干质量较其他坡向要低。半阳坡、半阴坡、阴坡白桦单叶片面积与出叶强度的异速生长指数分别为-0.89、-0.91、-0.85,与-1.0无显著差异(表3),且在不同坡向间有共同异速生长指数-0.89(图3B),表明二者之间存在负等速生长关系,且不随坡向发生改变,但其阴坡和半阴坡的异速生长常数大于半阳坡;即在共同斜率下,出叶强度一定时,其阴坡和半阴坡的单叶片面积大于其他坡向。
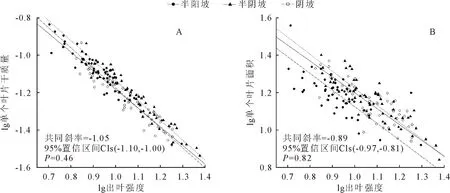
图3 白桦叶大小-数量在不同坡向的权衡Fig.3 Trade-off relationship of the leaf size-number of B.platyphylla in different aspect
3 讨论
3.1 坡向对白桦小枝性状的影响
生物量分配是现代生态学的核心概念之一,为不同植物的生长策略提供了基础,不同器官之间的生物量分配即意味着功能的权衡[16-17]。其中枝条生物量分配是植物净碳获取、周转的重要驱动因素。茎具有养分运输、贮存、叶片支撑、扩展生长空间等多重功能。叶片是光合作用的场所,进行物质和能量交换以及叶柄支持和供应叶片。影响着生物量分配的因素有很多,如光照、温度、降雨、土壤养分等[18-19]。坡向作为重要地形因子之一,可以起到再分配不同生境温度和光照的作用,从而间接影响植物生物量的分配格局[20]。本研究表明,半阳坡小枝茎干质量、总叶片干质量、总叶柄干质量最大,这可能是因为半阳坡的光照较为充足,有助于白桦的光合作用,固定CO2合成有机物质,促进干物质的积累[21]。
3.2 坡向对小枝和叶异速生长的影响
在小枝水平上,茎与小枝的异速生长指数与1.0无显著差异,呈等速生长,但其异速生长指数随着半阳坡到阴坡的改变呈现上升趋势,表明随着太阳辐射量的降低,白桦在阴坡的光照资源受限,逐渐增大对茎的投资,加强养分的运输和支撑叶片,扩展其生长空间,以获得更多的光照。卢艺苗等[22]的研究表明,下冠层小枝比上冠层小枝向茎投入的资源更多。除此之外,白桦总叶片干质量与小枝干质量和总叶片面积与小枝干质量呈等速生长关系,表明叶片的生物量独立于小枝生物量,此结果与杨冬梅等[1]对常绿和落叶阔叶植物的研究结果相一致。但孙蒙柯等[23]对武夷山常绿植物研究中发现,乔木叶重与小枝重呈等速生长而灌木则为异速生长。这可能是不同生活型植物在同一生境中,林下灌木与林冠层乔木在面对光照受限的情况下所采取的生长策略不同,乔木增大茎的投资,而灌木增大叶的投资,以适应环境的改变。
枝条生物量的分配可以在2个层次上进行探索,即枝条内和叶内。叶片被普遍认为是唯一对光合碳增益有直接贡献的功能组织,而叶柄被归入物理支撑结构的范畴。因此,确定植物小枝中叶片和叶柄的生物量分配模式,对于理解小枝大小演化具有重要意义[24]。叶片面积可以反映叶子拦截光线的能力[25],前人根据“管道模型”认为,叶片面积与叶柄重呈异速生长[4],但在本研究中总叶片面积与总叶柄干质量在阴坡呈等速生长,在半阴坡和半阳坡呈异速生长,这可能是因为阴坡的光照受限,会优先将资源用以叶面积的扩展,以获得更多的光照,其异速生长指数和异速生长常数都显示半阳坡<阴坡<半阴坡,也表明阴坡和半阴坡的白桦具有更大的叶片面积,可获得更多的光照。叶片干质量可以最直观地反映叶内生物量的分配状况[1,25]。研究发现,当对多个物种进行研究时,其呈现异速生长,不随环境发生改变[1],而对环境梯度下的单一物种研究时,其既有异速关系又有等速关系[26],表明单一物种在不同的生境条件下其异速关系会发生改变。本研究表明阴坡和半阳坡总叶柄干质量与总叶片干质量呈异速关系,在半阴坡呈等速关系。这可能是因为阴坡光照受限,植物为获得更多的光照,会加大对叶柄的投资,以扩展生长空间,避开遮荫的限制。半阳坡的总叶片干质量显著大于阴坡,叶为能支撑其重的叶片,也会加大对叶柄的投资。
3.3 坡向对叶片大小和数量的影响
研究发现,叶大小与叶数量之间存在显著的负等速关系,这与以前的研究结果相一致。而这种权衡关系有以下3种解释,首先是植物对外部环境选择压力的功能适应性的结果;其次植物内在的生物量分配限制的结果;再次是小枝大小[11]。除此之外,本研究还发现叶大小与叶数量的异速生长常数在不同坡向之间具有一定的差异。当叶大小以叶干质量表示时,其与出叶强度之间的异速生长常数在阴坡最低;当叶大小以叶面积表示时,其与出叶强度之间的异速生长常数在半阳坡最低,这表明在共同斜率下,出叶强度一定时,阴坡白桦具有较小的叶干质量,较大的叶面积,半阳坡具有大的干质量,小的叶面积。这可能是因为阴坡光照受限,投资更多资源用于扩展叶表面积,以接受更多的光照;而半阳坡光照较强和水分较少,叶片易蒸腾失水,从而加快叶片能量消耗、缩短叶片的光合碳获取时间,影响叶片寿命,因而半阳坡白桦呈现叶面积小而干质量较大的特性,便于热量和物质交换,并避免了植物组织面临水分亏缺的风险[27-28]。
4 结论
以冀北山地不同坡向白桦的当年生末端小枝为材料,结合异速生长理论,开展了白桦茎、叶、柄之间的生长关系研究,得出以下结论:半阳坡茎、叶片、叶柄干质量相对半阴坡和阴坡有0.06~0.11 g等不同程度的增长,表明半阳坡充足的阳光有利于干物质的积累。叶柄、茎、叶片在不同坡向的共同斜率为1.22、1.12和1.04,表明白桦在小枝干质量一定的情况下,会优先将资源分配给叶柄和茎,以扩展生长空间,获得更多的光照。白桦叶片大小和出叶强度在不同坡向呈负等速生长关系,但其异速生长常数表明,阴坡白桦具有叶片数量多、面积大且干质量小的性状,以适应坡向条件的变化。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29 10:10:20
南方农业(2022年23期)2022-02-14 09:45:26
思维与智慧·下半月(2021年7期)2021-08-13 16:55:24
东方少年·阅读与作文(2021年5期)2021-06-15 03:06:18
上海故事(2020年10期)2020-12-14 04:03:16
防护林科技(2019年7期)2019-08-19 00:50:58
现代农村科技(2018年11期)2018-11-15 11:27:58
河北地质(2017年2期)2017-08-16 03:17:02
课程教育研究(2017年11期)2017-04-17 07:47:23
少年文艺·少年读者文摘(2009年5期)2009-06-08 04:45:14
