
植物多样性对沿江湿地土壤温室气体排放及碳组分相关性探究
——以镇江江心洲为例
2024-01-17 06:57彭睿姚佳荆水强
能源与环境 2023年6期
彭睿 姚佳 荆水强
(江苏省镇江环境监测中心 江苏镇江 212001)
0 引言
近年来,全球温室气体排放研究成为了学术界热点,这与我国早日实现碳达峰、缓解温室效应的目标息息相关。土壤生态系统是温室气体主要排放来源之一[1],研究土壤碳组分与温室气体排放具有十分重要的意义,同时与土壤生态系统最为密切的就是分布在土壤表层的各类植物群落,植物群落的多样性对土壤的性质和功能起到十分重要的作用。湿地作为“地球之肾”,在自然生态系统中起到调节水分循环、维持生物多样性、净化污染物等作用。而湿地植物是维持湿地生态系统功能的重要因素[2],也是构建湿地“碳汇”能力的一个重要组成部分。本文通过对镇江市一江中小岛区域以植被群落特征为依据,选择不同的样地进行植物多样性调查,同时对样地内的土壤温室气体排放通量及碳组分进行测定,以研究两者间的相互关系。
1 研究区域概况
本项目研究区域位于江苏省镇江市丹徒区江心洲,属亚热带湿润气候,年平均气温15.6 ℃,年均降水量约1 088.2 mm。江心洲四面环江,岛内植物多样性十分丰富。长江直流贯穿小岛,形成岛内主干河流。由于该区域居民居住地十分分散,天然生长的植物与人为种植的作物在河流两侧均有分布(图1),因此,本文主要以该主干河流两侧天然植被群落景观与人为种植群落景观展开研究。

图1 调查区域及周边情况
2 研究方法
2.1 植物多样性调查
综合考虑江心洲主干河道两岸植被群落情况和分布特征,选择4 类不同的植被群落景观[乔木区(Site 1)、灌木区(Site 2)、草本区(Site 3)与种植区(Site 4)],一共13 个5 m×5 m样方进行植物多样性调研与环境(土壤与水)样品采集。13 个5 m×5 m 样方的近水端边界均设置在距水面1 m 处,以保证水源地距离对样方的影响。在每个5 m×5 m 样方内沿对角线设立3 个小样方(1 m×1 m),用以进行草本植物的调查(图2)。每周调查采样1 次、研究时间跨度为5 a,样方类别如表1 所示。
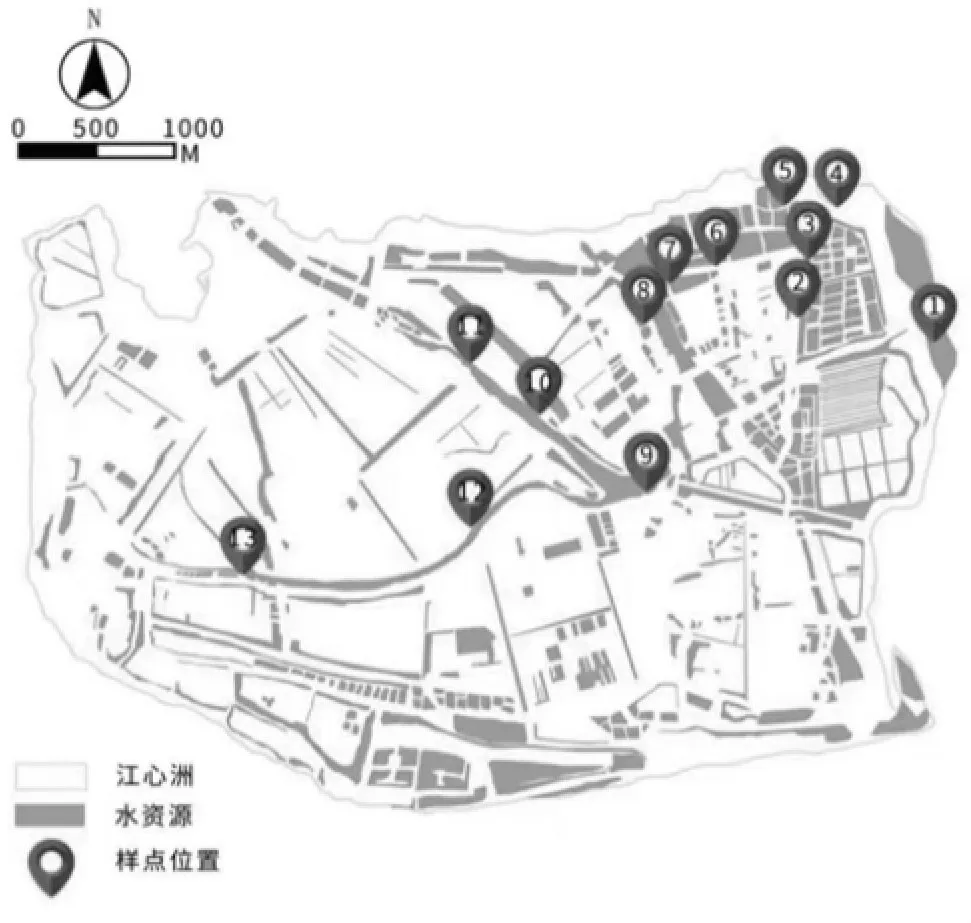
图2 样地点位设置情况

表1 样方类型与说明
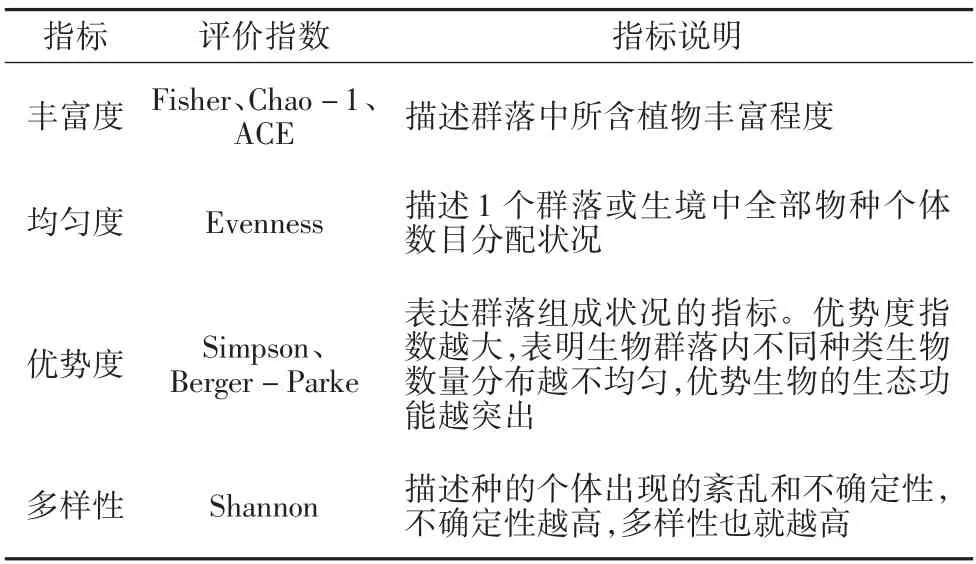
表2 植物多样性调查
植物的物种调查主要采用形态鉴定法、微观鉴定法、分子生物学法[3]3 种方法进行判别,多样性调查指标为数目、丰富度、均匀度、优势度以及多样性指数5 个维度。
2.2 土壤温室气体排放通量及碳组分调查
2.2.1 土壤温室气体排放测定
在采样日08∶00—11∶00 之间进行了温室气体的采集。将腔室放置在轴环凹槽上,旋塞密封注射器,在安装腔室后0、10、20、30 min,每个时间段用30 mL 塑料注射器收集腔室顶部空间的气体样本,将其注入集气袋中,每个集气袋收集60 mL气体。采样结束后送样至杭州水稻研究所通过气相色谱法测定二氧化碳、氧化亚氮与甲烷。
2.2.2 土壤碳组分测定
根据环境保护行业标准 《土壤环境监测技术规范》(HJ/T 166—2004),在每样方内沿对角线设立3 个小样方(1 m×1 m),在每个样方内按照对角线法,用土壤采集器采集0~20 cm 的土壤样品。每个采样点的3 个样方的土壤样品混合成为1 个混合样品,共采集12 个土壤样品。采集的土壤样品过2 mm 尼龙筛后用土壤采集布袋收集存放,带回实验室待进一步分析检测。其测量组分主要包括:土壤溶解性有机碳(DOC)、土壤总碳(TC)、土壤微生物生物量碳(MBC)、土壤总有机碳(TOC)、土壤易氧化有机碳(EOC)、重轻组有机碳(HOC/LOC)。
3 植物多样性调查结果
不同植物多样性维度条件下,调研区不同植物多样性指数呈现不同变化趋势,因此借助主成分分析统计学方法,对调研区多种维度的植物多样性指数进行整合分析,来探究调研区植物多样性是否呈显著性差异。主成分分析结果显示(图3),不同植被群落调研区域之间沿1 轴(x 轴)和2 轴(y 轴)区分明显,说明调研区植物多样性呈显著性差异(P<0.05)。主成分1 轴(x 轴)和2 轴(y 轴)对植物多样性的解释度分别为47.40%和37.59%。

图3 调研区植物多样性主成分分析
通过植物数目、丰富度指数、均匀度指数、优势度指数与多样性指数5 个维度,调查了镇江市丹徒区江心洲内长江贯穿主干河沿河湿地植物多样性分布情况。由调研结果可知,不同区域植物多样性差异显著。4 个调研区域植物丰富度多样性指标显示,乔木区>灌木区>草本区>种植区(P<0.05);而均匀度多样性指标显示乔木区<灌木区<草本区<种植区(P<0.05)。综合结果可知,乔灌草复杂群落垂直结构区域的植物多样性最高,而人为干扰改造区域(作物种植区)植物多样性最低。因此,推断人为景观改造是造成沿河湿地植物多样性差异的主要原因。
4 植物多样性对土壤碳组分的影响
4.1 土壤碳组分的赋存情况
镇江市江心洲调研区土壤碳组分的赋存情况如表3 所示。由表3 可知,土壤总碳(TC)含量在各研究区域无显著差异,分布范围为9.70~11.97 mg/g,其中,灌木区(Site 2)最高,草本区(Site 3)最低;土壤总有机碳(SOC)含量在各研究区域无显著差异,分布范围在8.92~10.55 mg/g,其中,种植区(Site 4)最高,草本区(Site 3)最低;土壤重组有机碳(HOC)含量在各研究区域差异显著(P<0.05),分布范围在2.79~5.75 mg/g,其中,种植区(Site 4)最高,乔木区(Site 1)最低;土壤轻组有机碳(LOC)含量在各研究区域无显著差异,分布范围在4.80~6.36 mg/g,其中,乔木区(Site 1)最高,种植区(Site 4)最低;土壤溶解性有机碳(DOC)含量在各研究区域无显著差异,分布范围在0.24~0.32 mg/g,其中,灌木区(Site 2)最高,种植区(Site 4)最低;土壤微生物生物量碳(MBC)含量在各研究区域无显著差异,分布范围在0.53~0.83 mg/g,其中,乔木区(Site 1)最高,灌木区(Site 2)最低;土壤易氧化有机碳(EOC)含量在各研究区域无显著差异,分布范围在0.91~1.75 mg/g,其中,乔木区(Site 1)最高,草本区(Site 3)最低。总体而言,不同植被群落多样性仅仅显著改变了土壤易氧化有机碳,说明在本文区域植被多样性的变化对土壤各种组分碳含量的影响较小。

表3 调研区土壤碳组分分布情况
利用主成分分析探究不同调研区与土壤碳组分的差异,主成分分析结果显示(图4),不同植被群落调研区域之间沿1 轴(x 轴)和2 轴(y 轴)区分明显,说明调研区土壤各种碳组分分布呈显著性差异(P<0.01)。主成分1 轴(x 轴)和2 轴(y轴)对土壤碳组分数据解释度分别为38%和27%。

图4 调研区土壤碳组分主成分分析
4.2 植物多样性与土壤碳组分的相关性分析
利用相关性热图探究调研区植物多样性与土壤碳组分相关性关系(图5),相关性热图结果显示,Individuals 指数与土壤碳组分均无显著相关关系;Simpson 指数与土壤碳组分均无显著相关关系;Shannon 指数与土壤碳组分均无显著相关关系;Evenness 指数与土壤微生物生物量碳(MBC)(R=-0.70,P<0.05)存在显著的负相关关系;Fisher 指数与土壤微生物生物量碳(MBC)(R=0.59,P<0.05)存在显著的正相关关系;Berger-Parker 指数与土壤碳组分均无显著相关关系(P<0.05);Chao-1指数与土壤微生物生物量碳(MBC)(R=0.66,P<0.05)存在显著的正相关关系;ACE 指数与土壤微生物生物量碳(MBC)(R=0.73,P<0.01)存在显著的正相关关系。

图5 调研区植物多样性与土壤碳组分相关性热图
冗余分析结果进一步说明植物多样性会影响土壤理化性质。主成分1 轴(x 轴)和2 轴(y 轴)对植物多样性数据解释度分别为52.84%和25.36%(图6)。土壤碳组分对植物多样性解释度结果显示,植物数目、丰富度指数、均匀度指数、优势度指数与多样性指数均会对土壤碳组分产生影响,其中丰富度指数对土壤碳组分的影响最为突出,其次为均匀度,且丰富度指数与相关性指数呈显著负相关关系。

图6 调研区植物多样性与土壤碳组分冗余分析
已有研究发现,不同利用方式下林地和天然草地在0~100 cm 土层总碳,轻组、重组、溶解性有机碳以及轻组有机碳分配比例均不同程度高于耕地[4],而其有机无机复合度则低于耕地,灌木林地和天然草地这种趋势尤为突出。土壤总碳、有机碳以及各组分有机碳之间呈极显著正相关,而无机碳则与其呈负相关,这与本文的结果相一致。同时,植物丰富度指数对土壤微生物生物量碳影响更为显著,可能是由于不同丰富度下的植被群落改变了土壤微生物群落结构,导致土壤微生物生物量碳随着植物丰富度的增加而增加,这与表3 中乔木区的微生物生物量碳最高相一致。
5 植物多样性对土壤温室气体排放的影响
5.1 调研区土壤温室气体排放的主成分分析
镇江市丹徒区江心洲调研区土壤温室气体分布情况如图7 所示。由图7 可知,二氧化碳在4 个调研区域差异显著(P<0.001),分布范围在133.12~392.81 mgC/(m2·h),其中,草本区(Site 3)最高,而灌木区(Site 2)最低;氧化亚氮在4 个调研区域差异显著(P<0.001),分布范围在0.014 7~0.149 3 mgN/(m2·h),其中,草本区(Site 3)最高,而乔木区(Site 1)最低;甲烷在4 个调研区域差异显著(P<0.001),分布范围在-0.056~0.022 mgC/(m2·h),其中,种植区(Site 4)最高,而草本区(Site 3)最低;全球变暖潜能值(GWP)在4 个调研区域差异显著(P<0.001),分布范围在136.77~431.92 mgC/(m2·h),其中,草本区(Site 3)最高,而灌木区(Site 2)最低。

图7 调研区土壤温室气体排放通量分布情况
在4 个调研样方中,对于土壤温室气体浓度最低的样方,二氧化碳(CO2)出现在种植区(Site 4),氧化亚氮(N2O)出现在草本区(Site 4),甲烷(CH4)出现在灌木区(Site 4),全球变暖潜能值(GWP)出现在乔木区(Site 4)。
利用主成分分析探究不同调研区与土壤理化性质的差异,主成分分析结果显示(图8),调研区土壤温室气体(P<0.01)分布呈显著性差异。主成分1 轴(x 轴)和2 轴(y 轴)对土壤温室气体的数据解释度分别为73%和22%。

图8 调研区土壤温室气体排放通量主成分分析
5.2 植物多样性与土壤温室气体的相关性分析
利用相关性热图探究调研区植物多样性与土壤理化性质相关性关系(图9),相关性热图结果显示,Individuals 指数与温室气体均无显著相关关系;Simpson 指数与温室气体均无显著相关关系;Shannon 指数温室气体均无显著相关关系;Evenness指数温室气体均无显著相关关系;Fisher 指数与甲烷(CH4)(R=0.75,P<0.01)存在显著的正相关关系;Berger-Parker 指数与温室气体均无显著相关关系;Chao-1 指数与甲烷(CH4)(R=0.61,P<0.05)存在显著的正相关关系;ACE 指数与甲烷(CH4)(R=0.59,P<0.05)存在显著的正相关关系。

图9 调研区植物多样性与土壤温室气体排放通量相关性热图
植物群落的结构和物种多样性都会影响土壤中碳输入的数量和质量,这可能会进一步改变土壤的温室气体排放[5]。考虑到植物对土壤的基质输入是温室气体排放的重要因素,不同研究区域的土壤养分存在显著差异,可以推断,植物基质投入的变异、数量和质量的差异可能本质上影响土壤的温室气体排放。
此外,由于根系可以提供温室气体的运输通道,并通过呼吸活动对二氧化碳排放产生贡献[6]。植物根系可以向土壤释放氧气,并重新氧化替代电子受体,如硫酸盐,从而抑制甲烷的产生,因为土壤中的碳基质与微生物竞争。根系密度和结构也会引起土壤通气性、碳氮沉积和利用模式的变化。此外,具有高生物量的多样化植物群落可以通过减少土壤中的硝态氮来减少氧化亚氮的排放。
冗余分析结果进一步说明植物多样性会影响土壤温室气体。主成分1 轴(x 轴)和2 轴(y 轴)对植物多样性数据解释度分别为90.28%和0.20%(图10)。土壤温室气体对植物多样性解释度结果显示,植物数目、丰富度指数、均匀度指数、优势度指数与多样性指数均会对土壤理化性质产生影响,其中植物丰富度指数对土壤温室气体的影响最为突出。

图10 调研区植物多样性土壤温室气体排放通量冗余分析
另一方面,土壤水分、养分有效性和土壤微生物群落被认为是影响温室气体排放的非生物和生物因素。由于这些非生物因子和生物因子与植物群落密切相关[7],植物群落景观多样性的变化必然会改变土壤的温室气体排放,本文再次证实了这一点。本文观测到的不同植物群落景观区土壤水分的差异可能是引起土壤温室气体排放差异的原因之一。土壤湿度与温室气体排放之间存在很强的相关性。据报道,土壤湿度直接或间接影响土壤温室气体的排放。一方面,土壤水分可以溶解二氧化碳、影响土壤物理结构(如孔隙度和团聚体),从而改变温室气体的运输。此外,土壤水分可以通过改变土壤中氧和可溶性养分的扩散和有效性来控制土壤微生物群落的组成、结构和活性,从而改变温室气体的产生。随着土壤水分的变化,土壤氧含量也发生变化。功能性微生物(如甲烷营养细菌和反硝化细菌)与氧气条件的变化无疑会影响生态过程(如反硝化、硝化和产甲烷)和温室气体的产生。不同植物群落景观多样性区域的土壤湿度差异可以进一步揭示土壤湿度对温室气体排放的非依赖性,从而解释景观多样性区域的温室气体排放格局。此外,养分利用率,特别是无机氮,容易受到土壤水分的影响,从而进一步调控温室气体的产生。
6 结论
通过主成分分析、相关性分析与冗余分析3 个维度,探究镇江市丹徒区江心洲长江贯穿河沿河湿地植物多样性对湿地土壤碳组分及温室气体排放有如下影响:
(1)植物丰富度指数对土壤碳组分的影响最为显著,其次为均匀度指数。其中,土壤微生物生物量碳受植物多样性影响最大。
(2)种植区(Site 4)土壤温室气体排放量最低是因为用于温室气体生产的植物和基质的丰度较低,不同区域的温室气体排放通量的影响,是由于植被群落结构和物种多样性的差异所导致的。
(3)植被群落多样性的变化,不仅直接影响土壤温室气体的排放,还会通过影响土壤理化性质从而间接影响土壤温室气体的排放。
猜你喜欢
农业工程技术(2022年1期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
煤气与热力(2021年12期)2022-01-19
云南农业(2021年9期)2021-09-24
云南农业(2021年8期)2021-09-06
食品安全导刊(2021年20期)2021-08-30
成都大学学报(自然科学版)(2021年1期)2021-05-22
英语文摘(2019年2期)2019-03-30
中成药(2018年8期)2018-08-29
中成药(2018年2期)2018-05-09
