一株耐低温异养硝化-好氧反硝化菌的分离鉴定及其脱氮特性
2024-01-12 12:41董怡华王凌潇任涵雪陈峰
生物技术通报 2023年12期
董怡华 王凌潇 任涵雪 陈峰
(1. 沈阳大学区域污染环境生态修复教育部重点实验室,沈阳 110044;2. 沈阳大学环境学院,沈阳 110044)
随着社会经济的快速发展,工业、农业和生活污水中氮含量急剧增加,过量的氮排放导致水体富营养化,进而引起水质恶化,严重破坏水环境生态系统,威胁人类健康[1]。因此,高效去除水中的氮已成为环境污染治理领域的研究热点。生物脱氮技术因其简单高效、成本低廉和无二次污染等优点而受到广泛关注[2]。传统的生物脱氮技术包括自养硝化和厌氧反硝化两个过程[3]。通常,硝化是自养硝化菌在好氧条件下将氨氮(NH4+‑N)转化为亚硝态氮(NO2-‑N)或硝态氮(NO3-‑N)的过程[4]。而反硝化则是通过异养反硝化菌在厌氧条件下将NO2-‑N或NO3-‑N 转化为氮气得以实现[5]。由于好氧硝化菌和厌氧反硝化菌的生长环境条件和代谢途径不同,导致硝化和反硝化作用不能同步进行,这不仅增加了投资成本和能耗,还影响了脱氮效率[6]。
20 世纪80 年代,一株好氧反硝化细菌——泛养硫球菌(Thiosphaera pantotropha)的首次发现,打破了反硝化作用只能在厌氧条件下进行的传统观念,为实现同步硝化反硝化工艺奠定了重要的微生物学基础[7]。此后,在众多微生物物种中不断分离出了多株异养硝化-好氧反硝化(heterotrophic nitrification and aerobic denitrification, HN‑AD)菌,如Microbacteriumsp. SFA 13[8]、Alcaligenes faecalisC16[9]、Pseudomonas aeruginosaP‑1[10]、Acinetobac‑ter juniiYB[11]等。这些功能菌不仅可以利用各种有机碳作为能源和电子供体,将NH4+‑N 转化为NO3-‑N 或NO2-‑N,还可以在好氧条件下逐步将NO3-‑N或NO2-‑N 还原为气态氮,从而实现同步硝化和反硝化作用[12]。此外,好氧反硝化过程中产生的碱还可以平衡硝化作用产生的酸,从而降低调节pH 的额外成本[13]。与传统自养硝化菌和厌氧反硝化菌相比,HN‑AD 菌具有诸多优势,包括更快的生长速率和脱氮速率,更低的操作成本和空间占用,以及在氮转化过程中更少的中间产物残留量等[14]。然而,文献中报道的大多数HN‑AD 菌都是中温细菌,其细胞生长和脱氮的最佳温度为25-37℃。当温度低于15℃时,这类功能菌的生长和代谢活性就会受到严重抑制[15-16]。寒冷地区冬季的地表水和大部分生活污水温度均在15℃以下,甚至低于5℃。在这种情况下,HN‑AD 菌对实际含氮废水的处理效果较差。分离筛选高效耐低温HN‑AD 菌是解决上述问题的一种有效方法。然而目前其相关研究尚处于起步阶段,只有少数HN‑AD 细菌在低温条件下能够有效去除氮,如Arthrobacter arilaitensisY‑10[17]和Acinetobacter calco‑aceticusTY1[18]。此外,这类功能菌在低温条件下的脱氮特性和氮去除途径仍不明晰。
本研究从寒冷地区冬季河水底泥中分离、筛选得到一株耐低温HN‑AD 菌株,通过菌落及细胞形态特征观察和16S rRNA 基因序列分析对其进行了菌种鉴定。在10℃条件下考察了以NH4+‑N、NO3-‑N 或NO2-‑N 为唯一氮源,以及NH4+‑N 和NO3-‑N 为混合氮源时,该菌株的异养硝化、好氧反硝化和同步硝化反硝化性能。最后,通过PCR 扩增了硝化和反硝化特异酶的相关功能基因,并由此推测了该菌株在低温条件下可能的氮代谢途径。本研究为耐低温异养硝化-好氧反硝化细菌在含氮废水净化中的实际应用提供了一种新的菌种资源,为低温废水生物脱氮处理奠定了理论基础和技术支持。
1 材料与方法
1.1 材料
1.1.1 样品来源 本研究所用样品取自冬季辽宁省沈阳市南部污水处理厂二沉池活性污泥和北运河河水底泥沉积物,并置于4℃冰箱中保存。
1.1.2 培养基组成 牛肉膏蛋白胨培养基(g)[19]:牛肉膏3.0,蛋白胨10.0,NaCl 5.0,蒸馏水 1 L,pH 调节至7.0-7.2。在培养基中添加2%琼脂粉即为对应的固体培养基。该培养基用于细菌的培养、菌落的形态观察和菌株保存。
参照汤默然等[20]有所改进的异养硝化培养基(nitrification medium, NM)(g):Na3C6H5O7·2H2O 2.0,(NH4)2SO40.5,K2HPO40.8,MgSO4·7H2O 0.4,NaCl 0.4,FeSO4·7H2O 0.04,CaCl2·H2O 0.03, 蒸馏水1 L,微量元素溶液1 mL,pH 调节至7.0-7.2。该培养基用于菌株的硝化性能研究。
参照汤默然等[20]有所改进的好氧反硝化培养基(denitrification medium, DM)(g):Na3C6H5O7·2H2O 2.5,NaNO30.64(DM‑1 培养基)或NaNO20.52(DM‑2培养基),K2HPO40.8,MgSO4·7H2O 0.4,NaCl 0.4,FeSO4·7H2O 0.04,CaCl2·H2O 0.03,蒸馏水1 L,微量元素溶液1 mL,pH 调至7.0-7.2。该培养基用于菌株的反硝化性能研究。
溴百里酚蓝(BTB)固体培养基:在DM‑1 培养基中添加0.2%溴百里酚蓝(体积分数为1%,溶于乙醇)和2%琼脂粉。
参照NM 培养基并有所改进的同步硝化反硝化培养基(simultaneous nitrification and denitrifica‑tion medium, SNDM)(g):Na3C6H5O7·2H2O 2.5,(NH4)2SO40.25,NaNO30.32,K2HPO40.8,Mg‑SO4·7H2O 0.4,NaCl 0.4,FeSO4·7H2O 0.04,Ca‑Cl2·H2O 0.03,蒸馏水1 L,微量元素溶液1 mL,pH调至7.0-7.2。该培养基用于菌株的同步硝化反硝化性能研究。
上述培养基中的微量元素液组成(g):H3BO31.5,ZnSO4·7H2O 0.06,MnSO4·H2O 0.8,CuSO4·5H2O 0.1,CoSO4·7H2O 0.01,(NH4)6Mo7O24·4H2O 0.2,KI 0.18,蒸馏水1 L。
上述所有培养基均在接种前于121℃,1.0×105Pa 条件下高温高压灭菌30 min。
1.2 方法
1.2.1 耐低温异养硝化-好氧反硝化菌的分离筛选 取5 g 活性污泥或河水底泥用无菌玻璃珠打散均匀后,转接至含有100 mL 牛肉膏蛋白胨液体培养基的250 mL 锥形瓶中,并于10℃、140 r/min 的恒温摇床中培养72 h。重复上述相同步骤3 次后获得细菌培养物。用无菌去离子水将1 mL 上述富集培养物按浓度梯度10-3、10-4、10-5和10-6倍进行稀释,吸取每种浓度下的稀释菌液100 μL 均匀涂布在BTB固体培养基上,并于10℃的恒温生化培养箱中倒置培养5 d。挑取周围变蓝且具有明显形态差异的独立菌落进行平板划线分离培养,重复多次直到在光学显微镜下确定为纯菌,然后进行斜面试管培养,并置于冰箱4℃下保存。
用接种环挑取分离的不同纯菌以相同接种量(1×106CFU/mL)分别接种于100 mL 的NM、DM‑1和DM‑2 液体培养基中,于10℃、140 r/min 的恒温振荡培养箱中培养48 h,取培养液于6 000 r/min 离心10 min 后测定上清液中NH4+‑N、NO3-‑N、NO2-‑N和TN 的质量浓度,从而对分离的纯菌进行硝化反硝化能力检测。筛选除氮效果最好且NO2-‑N 残留量最少的菌株作为后续研究对象。
1.2.2 菌株的鉴定 将所分离筛选的纯菌株接种至牛肉膏蛋白胨固体培养基上,培养3 d 后观察菌落特征。利用扫描电子显微镜(SEM, Thermo Scientific Apreo 2, USA)观察菌株细胞的个体形态,并进行革兰氏染色。
采用Ezup 柱式细菌基因组DNA 提取试剂盒SK8255(Sangon Biotech,上海)提取菌株的DNA,并利用通用引物27F 和1492R(表1)进行16S rRNA 基因的扩增。PCR 反应体系(50 μL):模板DNA 1 μL,10×PCR Buffer 5 μL,dNTPs 4 μL,Taq Plus DNA 聚合酶1 μL,MgSO42.5 μL,引物27F 和1492R 各1 μL,ddH2O 9.5 μL。PCR 反应条件为:95℃预变性4 min;9℃变性45 s,55℃退火45 s,72℃延伸1 min,共30 个循环;72℃修复延伸10 min。PCR 产物由生工生物工程(上海)股份有限公司完成测序。通过BLAST 软件与GenBank 基因数据库中其他菌株进行16S rRNA 基因序列同源性对比,选择序列相似度较高的菌株,利用MEGA7.0 软件通过邻接法(neighbor‑joining, NJ)法构建系统发育树。

表1 PCR 引物序列表Table 1 PCR primer sequences list
1.2.3 功能基因的PCR 扩增 采用试剂盒B518255(Sangon Biotech,上海)提取菌株的DNA,对周质硝酸盐还原酶(NAP)、羟胺氧化酶(HAO)、亚硝酸盐还原酶(NIR)、一氧化氮还原酶(NOR)、一氧化二氮还原酶(NOS)的编码基因napA、hao、nirS、nirK、cnorB和nosZ分别进行PCR 扩增,PCR 特异性目标引物序列见表1。PCR 反应体系(50 μL):模板DNA 1 μL,引物F 和R 各1 μL,10×Taq Buffer 2.5 μL,dNTPs 1 μL,Taq Plus DNA 聚合酶 0.2 μL,ddH2O 25 μL。PCR 的反应条件:95℃预变性5 min;94℃变性30 s,63℃退火30 s,72℃延伸30 s,10个循环;95℃变性30 s,58℃退火30 s,72℃延伸30 s,30 个循环;72℃修复延伸10 min。PCR 产物经2%琼脂糖凝胶电泳分析。
1.2.4 菌株的异养硝化-好氧反硝化性能 以(NH4)2SO4、NaNO3和NaNO2分别作为NM、DM‑1和DM‑2 培养基中的唯一氮源,探索菌株在低温条件(10℃)下的异养硝化和好氧反硝化性能。采用(NH4)2SO4+NaNO3作为SNDM 培养基中的混合氮源,考察菌株在低温(10℃)条件下的同步硝化反硝化能力。上述培养基中的N 含量均为105 mg/L。将所筛选的菌株培养至对数生长期后,用无菌去离子水配制浓度为1×106CFU/mL 的菌悬液,以10%(体积分数)的接种量分别接种至150 mL 的NM、DM‑1、DM‑2、SNDM 无菌培养基中,在10℃、140 r/min 的恒温摇床中培养48 h。每4 h 取样测定菌体生长量(D600)、NH4+‑N、NO3-‑N、NO2-‑N 和TN 的质量浓度,并计算相应的氮去除率和氮去除速率。
1.2.5 分析方法 通过测定细菌培养液在600 nm 波长处的吸光度值(D600)表征菌体生长量。总氮(total nitrogen, TN)质量浓度是培养液摇匀后直接取样分析测定。NH4+‑N、NO3-N、NO2-‑N 和溶解性总氮(dissolved total nitrogen, DTN)是将培养液以6 000 r/min 离心10 min 后取上清液测定。上述氮指标的测定根据水和废水监测分析方法进行[27],如表2 所示。实验中对各项氮指标以及菌体生长量的测定均采用紫外-可见分光光度计(DR6000, HACH, USA)进行。

表2 水质指标及分析方法Table 2 Analysis methods for water quality indicators
上述测定结果用以下公式计算菌株的生长速率、脱氮率、细胞内生物氮和气态氮质量浓度等指标。
① 生长速率μ/h 的计算如下:
式中,D600、D'600分别是细菌培养液培养初始和终态在600 nm 波长处的吸光度值,用以表征菌体生长量,t 为培养时间,单位为h。
② 氮去除率R(%)和氮去除速率V[mg/(L·h)]的计算如下:
式中,C0和Ct分别是培养液中各种形态氮的初始质量浓度和培养t h 后的质量浓度,单位为mg/L。
③ 细胞内生物氮质量浓度CIntracellular‑N(mg/L)的计算方法为[26]:
式中,CTN和CDTN分别为培养液的TN 质量浓度及其离心后上清液DTN 质量浓度。
④ 气态氮质量浓度CGas‑N( mg/L)的计算方法:
其中,C0为氮的初始质量浓度,和分别为t h 后培养液离心上清液中NH4+‑N、NO3-‑N、NO2-‑N 的质量浓度。
本研究所有实验重复3 次,结果用平均值±标准偏差(standard deviation, SD)表示。采用Excel 和Origin Pro 2021 软件对实验结果进行统计分析与绘制图形。
2 结果
2.1 耐低温异养硝化-好氧反硝化菌的分离筛选与鉴定
2.1.1 菌株的分离筛选 经过富集培养、划线分离和纯化,从活性污泥和河水底泥样品中分离出19 株能够在低温(10℃)条件下生长,且菌落形态不同的纯菌株。通过异养硝化-好氧反硝化能力的初步检测,从中筛选出1 株在10℃条件下脱氮效果最好且NO2-‑N 累积量最少的菌株DH‑3,培养48 h 后对NH4+‑N、NO3-‑N 和NO2-‑N 的去除率分别为99.07%、96.89%和90.29%。
2.1.2 菌株的形态学观察 菌株DH‑3 是从冬季河水底泥样品中分离得到,其在牛肉膏蛋白胨固体培养基上生长的菌落呈淡黄色不透明,边缘规则整齐,表面光滑凸起有黏性,直径约为1-3 mm(图1‑A);经革兰氏染色为革兰氏阴性菌(图1‑B),扫描电镜下观察其细胞呈长杆状,大小为(0.1-0.2)μm×(1.5-2.8)μm(图1‑C)。
2.1.3 分子生物学鉴定 通过PCR 扩增和测序获得菌株DH‑3 的部分16S rRNA 基因序列(1 428 bp),测序结果在GenBank 中的登录号为OQ152538。与NCBI 数据库中已有基因序列进行BLAST 同源性对比,采用MEGA7.0 软件通过邻接法(neighbor‑joining, NJ)法构建系统发育树(图2),结果表明菌株DH‑3 与维氏假单胞菌(Pseudomonas veronii)的基因序列相似性达99%以上,其中与Pseudomonas veroniistrain CIP 104663(NR 028706.1)基因片段的亲缘性最高。
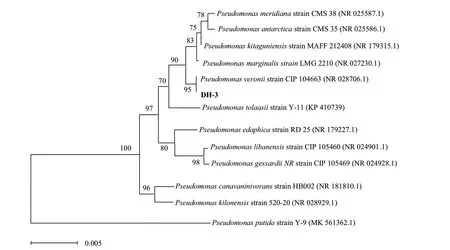
图2 基于16S rRNA 基因序列同源性构建的菌株DH-3 系统发育树Fig. 2 Phylogenetic tree of strain DH-3 based on 16S rRNA homologous sequences
综上,结合菌株DH‑3 的菌落特征、细胞形态学特征以及16S rRNA 基因序列分析结果,初步鉴定该菌株为维氏假单胞菌(Pseudomonas veronii),命名为P.veroniiDH‑3。
2.2 菌株P. veronii DH‑3的低温异养硝化性能
在去除碳源(Na3C6H5O7·2H2O)的NM 培养基中进行探索试验,培养48 h 后,培养液的D600为0,表明碳源的缺乏严重抑制了菌株DH‑3 的生长,由此证明菌株DH‑3 是一株异养菌。以(NH4)2SO4作为NM 培养基中的唯一氮源,考察10℃条件下菌株P.veroniiDH‑3 的异养硝化性能。脱氮过程中菌体生长量和各种氮指标的质量浓度变化如图3 所示。
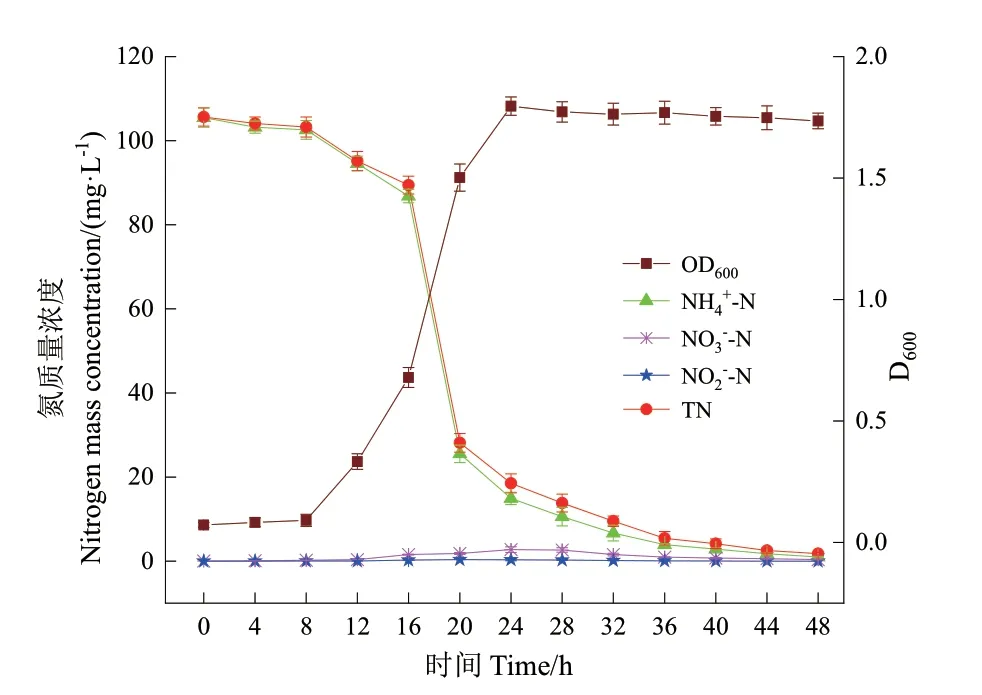
图3 以NH4+-N 为唯一氮源时菌株DH-3 的生长及异养硝化特性Fig. 3 Growth and heterotrophic nitrification performance of strain DH-3 with NH4+-N as the sole nitrogen source
经过8 h 的生长停滞期后,菌株P.veroniiDH‑3迅速生长繁殖并进入对数生长期,24 h 时D600达到最大值1.796,随后进入稳定生长期,D600变化不大。NH4+‑N 的去除率随菌株的生长繁殖而升高。好氧培养48 h 后,NH4+‑N 浓度从初始的105.5 mg/L 下降至0.98 mg/L,去除率为99.07%。培养48 h 后,TN 质量浓度从105.6 mg/L 显著下降到1.78 mg/L,去除率为98.31%。在整个脱氮过程中没有检测到中间产物NO2-‑N 的累积,而NO3-‑N 在培养24 h 出现最大累积浓度值2.74 mg/L,48 h 后降至0.37 mg/L。
表3 的氮平衡分析结果表明,约有53.9%的初始NH4+‑N 通过硝化作用转化为含氮气体(如N2O 或N2),而约44.8%的初始NH4+‑N 通过同化作用转化为细胞内生物氮,这与之前得到的NH4+‑N 去除率与菌株DH‑3细胞生长密切相关的结果是一致的。因此,菌株P.veroniiDH‑3 的异养硝化过程主要是在好氧条件下通过同化和异化作用将NH4+‑N 转化为细胞内生物氮和含氮气体。
2.3 菌株P. veronii DH‑3的低温好氧反硝化性能
以NO3-‑N 和NO2-‑N 分别作为DM‑1 和DM‑2培养基中的唯一氮源,考察菌株P.veroniiDH‑3 在10℃下的生长和好氧反硝化性能,结果如图4 和图5 所示。
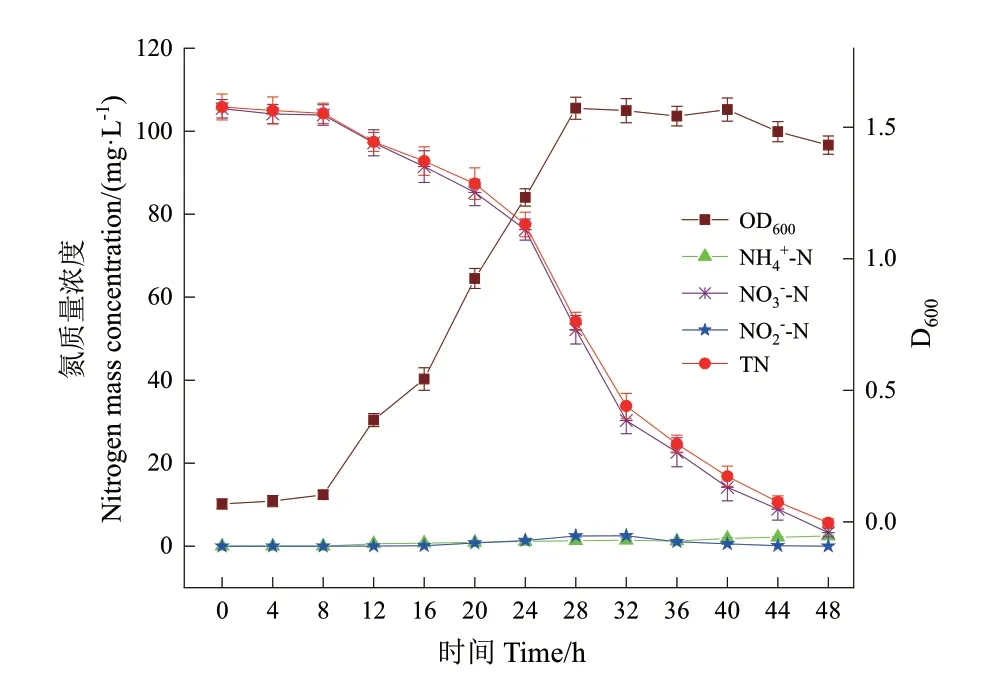
图4 NO3--N 为唯一氮源时菌株DH-3 的生长及好氧反硝化特性Fig. 4 Growth and aerobic denitrification performance of strain DH-3 with NO3--N as the sole nitrogen source
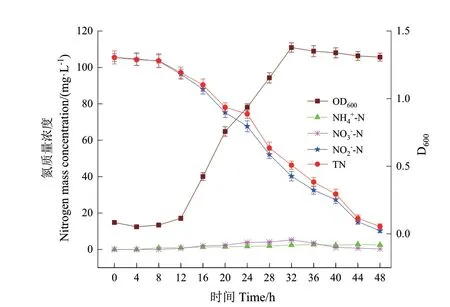
图5 NO2--N 为唯一氮源时菌株DH-3 的生长及好氧反硝化特性Fig. 5 Growth and aerobic denitrification performance of strain DH-3 with NO2--N as sole nitrogen source
由图4 可知,以NO3-‑N 为唯一氮源时,菌株P.veroniiDH‑3 在好氧条件下培养0-8 h,D600仅从0.068增加至0.103,此阶段为菌株的生长停滞期;随着菌株进入对数生长期(8-28 h)的快速增殖,D600达到最大值1.572;之后菌株进入稳定期;培养40 h后菌株进入衰亡期,D600逐渐下降。在对数生长期,NO3-‑N 浓度从105.43 mg/L 迅速下降至52.18 mg/L,48 h 后NO3-‑N 的去除率为96.89%。在氮转化过程中积累了少量的NO2-‑N,32 h 达到最大累积浓度2.51 mg/L,48 h 后被完全去除。
由图5 可知,以NO2-‑N 为唯一氮源时,菌株DH‑3 经历了12 h 的生长停滞期以适应NO2-‑N 的细胞毒性。随后菌株迅速生长并进入对数生长期,培养32 h 时D600达到最大值1.378,之后逐渐降低进入衰亡期。培养48 h 后,NO2-‑N 质量浓度从105.53 mg/L 下降到10.24 mg/L,去除率为90.29%。培养过程中检测到在32 h 出现NO3-‑N 最大累积浓度5.27 mg/L,之后逐渐降低至0.25 mg/L。菌株DH‑3 以NO3-‑N 和NO2-‑N 分别为唯一氮源时对TN 的去除率为94.69%和88.03%,低于以NH4+‑N 为唯一氮源时的去除率(98.31%)。表3 的氮平衡分析结果表明,分别有52.6%的初始NO3-‑N和50.4%的初始NO2-‑N 通过反硝化作用转化为含氮气体,相应的细胞内生物氮含量分别为(42.76 ±2.07)mg/L 和(38.58 ± 2.31)mg/L,气体和细胞内生物氮的转化率均低于NH4+‑N 为唯一氮源时的转化率,说明NH4+‑N 更容易被菌株DH‑3 所利用,同时表明该菌株的异养硝化能力强于好氧反硝化能力。
2.4 菌株P. veronii DH‑3的同步硝化反硝化特性
以NH4+‑N 和NO3-‑N 作为混合氮源,研究了菌株P. veroniiDH‑3 在10℃下的同步硝化反硝化性能,结果如图6 所示。
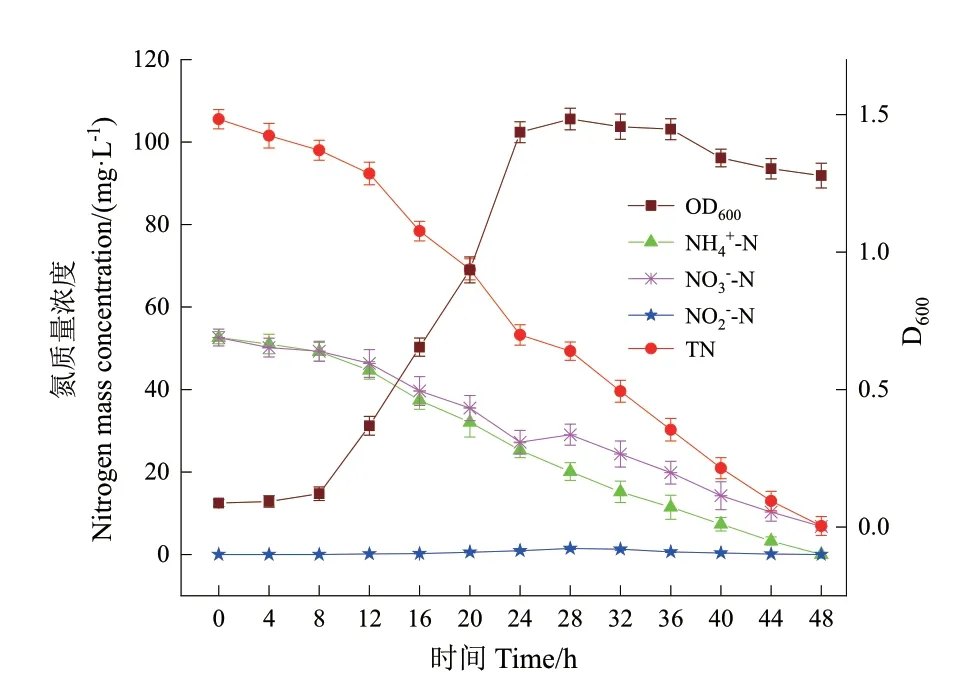
图6 NH4+-N 和NO3--N 为混合氮源时菌株DH-3 的生长及脱氮特性Fig. 6 Growth and SND performance of strain NR-5 with NH4+-N and NO3--N as mixed nitrogen source
由图6 可知,菌株DH‑3 在SNDM 培养基中于10℃条件下培养28 h 后菌体生长量达到最大值1.484,之后进入到稳定生长期;培养36 h 后菌株进入衰亡期。NH4+‑N 在48 h 内被完全去除。与NH4+‑N 浓度始终下降的趋势不同,NO3-‑N 浓度在最初的24 h 内从52.61 mg/L 下降到27.24 mg/L,然后在28 h 略微升高到29.05 mg/L,可能是该菌株在异养硝化过程中积累了少量硝酸盐所致。NO3-‑N 在48 h 内的去除率为87.09%,平均去除速率为0.95 mg/(L·h)。TN 在48 h 内的去除率为93.45%,没有检测到NO2-‑N 的积累。表3 的氮平衡分析结果表明,约有51.2%的初始NH4+‑N 和NO3-‑N 在同步硝化反硝化作用转化为含氮气体。上述结果进一步证实了P.veroniiDH‑3 是一株有效的HN‑AD 菌,能够在低温好氧条件下实现同步硝化反硝化脱氮。
2.5 菌株的功能基因检测
基因hao、napA、nirS、nirK、cnorB 和nosZ 常被认为是参与HN‑AD 过程的关键功能酶基因。在本研究中,通过PCR 扩增这些潜在基因,以期深入阐明菌株P.veroniiDH‑3 低温脱氮途径,结果如图7所示。
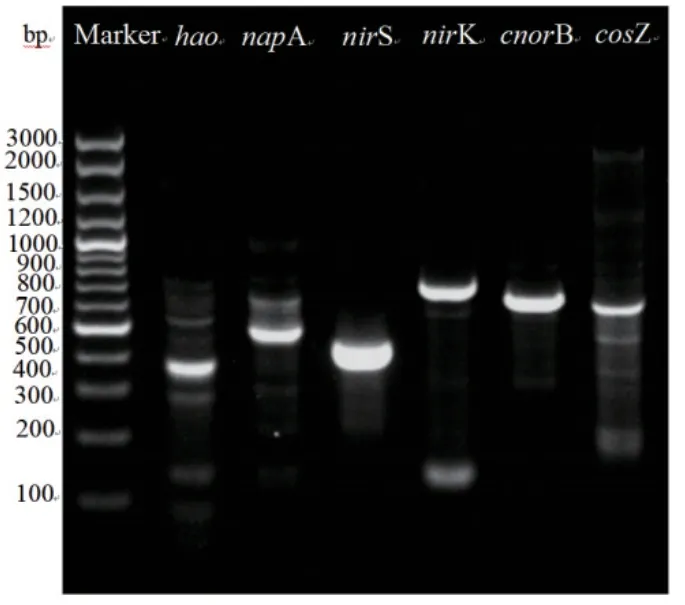
图7 菌株DH-3 功能基因的扩增Fig. 7 PCR amplification results of functional genes in strain DH-3
由图7 可知,该菌株成功扩增出hao、napA、nirS、nirK、cnorB 和nosZ 的基因产物,其对应的序列长度分别为365、452、414、642、575 和500 bp。这些功能基因产物的成功扩增表明菌株P.veroniiDH‑3 可产生参与HN‑AD 过程的关键功能酶,进一步证明该菌株具有硝化反硝化的能力。
3 讨论
低温菌是一种极端微生物,根据生长温度特性可分为嗜冷菌(Psychrophilies)和耐冷菌(Psychro‑trophs)两类[28]。其中,耐冷菌的关键特征是能够在较宽泛的温度(0-30℃)下有效代谢营养物质,产生各种酶类,促进低温水体中营养物质的转化与能量流动,维持水体环境中生物群落的稳定和生态系统平衡,因此在实际推广应用中具有显著的经济效益和环境效益[29],是目前微生物处理低温废水研究的热点。本研究从东北寒冷地区冬季河水底泥样品中分离出一株耐低温HN‑AD 菌P.veroniiDH‑3,与Pseudomonassp.中一些具有HN‑AD 能力的菌株具有较高的同源性,例如Pseudomonas tolaasiiY‑11(KP 410739)[30]和P. putidaY‑9(MK 561362.1)[6]。目前国内外关于P.veronii在低温条件下作为异养硝化-好氧反硝化菌的研究尚未见报道。
3.1 菌株P. veronii DH‑3具有较快的生长速率和良好的异养硝化性能
大多数中温HN‑AD 菌以NH4+‑N 为唯一氮源时生长停滞期较短,例如,在30℃条件下,Exiguo‑bacterium mexicanumSND‑01[31]能够快速生长,没有停滞期,而Ochrobactrum anthropicLJ81[24]的停滞期为4 h。相比之下,菌株P.veroniiDH‑3 需要较长的生长停滞期(8 h)以适应低温环境。然而,在低温条件下,菌株DH‑3 的生长停滞期短于其他耐冷HN‑AD 菌,例如Bacillus simplexH‑b 在10℃下为24 h[32],Arthrobacter arilaitensisY‑10 在15℃下为12 h[17]。此外,检测到菌株DH‑3 在培养16-20 h时出现最大生长速率0.21/h,高于一些文献报道的HN‑AD 细 菌, 如Acinetobacter sp. YS2(30℃,0.11/h)[33]和Acinetobactersp. ND7(28℃,0.12/h)[26]。上述结果表明,P.veroniiDH‑3 在10℃条件下具有较短的代际时间和较快的生长速度,这对于低温含氮废水的生物处理具有重要意义。
当NH4+‑N 为唯一氮源时,P.veroniiDH‑3 对NH4+‑N 的平均去除速率为2.18 mg/(L·h),高于Moraxella osloensisLT‑01[1.48 mg/(L·h),10℃][34]和Arthrobacter arilaitensisY‑10[0.4 mg/(L·h),15℃][17],甚至比某些中温菌,如Pseudomonassp.ADN‑42[1.38 mg/(L·h),30℃] 更具优势[35]。Guo 等[36]对P. indoloxydansYY‑1 转录组的分析,初步揭示了HN‑AD 细菌的耐冷机制是由参与TCA循环、氨基酸代谢和冷休克蛋白相关的一些关键酶随着温度的降低而上调所引起的,而NH4+‑N 为这些关键酶提供了额外的必要能量需求。
3.2 菌株P. veronii DH‑3具有良好的好氧反硝化性能
以NO3-‑N 为唯一氮源时,菌株P.veroniiDH‑3对NO3-‑N 平均去除速率为2.13 mg/(L·h),明显高于P. putidaY‑12[1.57 mg/(L·h),15℃][37]和Rhodococcussp. CPZ24[0.93 mg/(L·h),30℃][38]。先前的研究表明[39],反硝化过程中NO3-‑N 和NO2-‑N 存在相互转化,而本研究的氮转化过程中尽管出现少量的NO2-‑N 累积,然而48 h 后被完全去除。推测原因可能是由于NO3-‑N 在脱氮过程中被转化的同时,中间产物NO2-‑N 也被用作氮源以维持微生物所需的生物合成,从而导致NO2-‑N 浓度下降。此外,在脱氮过程中还检测到微量NH4+‑N 存在,这可能是由死亡菌体细胞发生裂解而释放出胞内氮所引起的[40]。
菌株P. veroniiDH‑3 以NO2-‑N 为唯一氮源时,培养48 h 平均去除速率为2.13 mg/(L·h),低于以NO3-‑N 为唯一氮源时的平均去除率[1.99 mg/(L·h)]。先前的研究表明,当NO2-‑N 浓度超过30 mg/L 时会阻碍微生物生长,从而抑制反硝化活性[24,30],本研究结果与之相似。然而,该菌株对NO2-‑N 的最大瞬时去除速率为4.87 mg/(L·h),仍然高于其他耐冷反硝化细菌,例如Bacillus simplexH‑b[0.18 mg/(L·h),10℃][32]和Acinetobacter calcoaceticusTY1[2.74 mg/(L·h),8℃][18]。
3.3 菌株P. veronii DH‑3具有良好的同步硝化反硝化性能
菌株P.veroniiDH‑3 具有良好的同步硝化反硝化特性。以NH4+‑N 和NO3-‑N 为混合氮源时,NO3-‑N在48 h 内的平均去除速率为0.95 mg/(L·h),低于DM‑1 培养基中的平均去除率[2.13 mg/(L·h)]。分析上述现象的原因可能是由于NH4+‑N 和NO3-‑N的共存导致它们之间出现“反馈抑制”,从而影响硝酸盐的去除[23]。此外,菌株DH‑3 在SNDM 培养基中对NO3-‑N 的平均去除速率低于NH4+‑N,再次证明该菌株优先利用NH4+‑N 而不是NO3-‑N,这个结果与Acinetobactersp. JR1[31]和Acinetobactersp.C‑13[41]特性一致,这可能是由于菌株的羟胺氧化还原酶(HAO)的活性高于硝酸盐还原酶(NR)。因此可以推断HN‑AD 细菌对氮源的利用可能与其相应的催化酶活性有关。
3.4 菌株P. veronii DH‑3的脱氮途径
生物脱氮是将氨氮和硝氮转化为含氮气体的过程,为了研究菌株P.veroniiDH‑3 的脱氮途径,选择了6 个潜在的硝化反硝化酶及基因进行PCR 扩增。hao是硝化过程中的关键酶基因,它通过编码羟胺氧化酶(HAO)催化异养硝化的中间产物NH2OH氧化为NO2[42]。从菌株P.veroniiDH‑3 中成功扩增得到hao功能基因,表明该菌株具有潜在的异养硝化能力。先前的文献报道从其他HN‑AD 菌中也扩增出了hao基因,如蜡样芽孢杆菌GS‑5(Bacillus cereusGS‑5)[43]和不动杆菌ND7(Acinetobactersp.ND7)[26]。napA 基因是在好氧条件下参与硝酸盐呼吸和反硝化作用的关键酶基因,能够转录表达周质硝酸盐还原酶(NAP)[44],该酶可以在有氧条件下将NO3-‑N 还原为NO2-‑N,因此napA 基因常被作为鉴定好氧反硝化菌的典型标志[45]。亚硝酸盐还原酶(NIR)负责将NO2-‑N 催化转化为NO[46],根据其结构和辅因子分为两类:以血红素为辅因子的同二聚体细胞色素cd1 型亚硝酸盐还原酶(cd1‑NIR)和以Cu 为辅因子的铜型亚硝酸还原酶(Cu‑NIR),这两种酶分别有由nirS 和nirK 基因编码[47]。先前研究普遍认为nirS 和nirK 基因不能在同一微生物细胞中共存[43]。然而,本研究从菌株P.veroniiDH‑3 中扩增出了nirS 基因和nirK 基因,表明这两种基因同时存在于菌株DH‑3 中,以实现亚硝酸盐的转化。同样在Ochrobactrum anthropicLJ81[25]中也观察到了两种功能基因共存的现象。此外,nirS 和nirK 的共存也表明在NO3-‑N 去除过程中,NO2-‑N 的积累可能较少,这与之前的脱氮实验中未检测出NO2-‑N 的结果一致。从菌株P.veroniiDH‑3 中还成功扩增出了表达一氧化氮还原酶(NOR)的cnorB 基因和表达一氧化二氮还原酶(NOS)的nosZ 基因,这两种关键酶分别负责将NO 转化为N2O 以及将N2O 转化为N2。cnorB 基因和nosZ 基因的成功扩增表明该菌株更有可能在不产生N2O 的情况下实现完全脱氮[35]。
基于HN‑AD 性能研究、氮平衡分析和关键酶功能基因扩增结果推测菌株P.veroniiDH‑3 的脱氮途径如图8 所示。
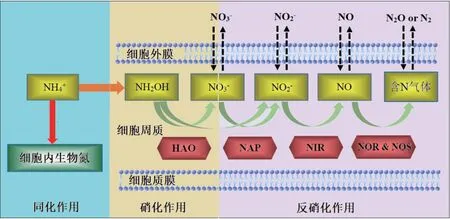
图8 菌株DH-3 的氮代谢途径Fig. 8 Proposed nitrogen metabolism pathway of strain DH-3
根据先前的研究报道,HN‑AD 菌株的脱氮主要包括以下两种代谢途径[31]:
在本研究中,菌株P.veroniiDH‑3 在低温条件下的脱氮过程遵循代谢途径I,这与大多数其他HN‑AD 细菌的代谢途径基本一致,如Microbacteriumsp. SFA13[8]、Acinetobacter sp. HA2[48]和Halomonassp. H36[49]。菌株DH‑3 可以利用无机氮化合物(如铵、亚硝酸盐和硝酸盐)作为唯一或混合氮源,通过同化作用将其转化为细胞内生物氮,以及通过HN‑AD 作用将其转化成含氮气体。该菌株异养硝化过程的第一步是利用柠檬酸钠作为碳源和电子供体,将NH4+‑N 氧化为NH2OH,然后由HAO 进一步氧化为NO2-‑N。在好氧反硝化过程中,NAP 可以将NO3-‑N 还原为NO2-‑N。随后,通过功能酶NIR、NOR 和NOS 依次将NO2-‑N 转化为NO、N2O 和N2。具有这种代谢途径的HN‑AD 细菌可以转化多种形式的氮,如NH4+‑N、NO2-‑N 和NO3-‑N,使这类细菌在环境中具有广泛的生态位和良好的环境适应性,从而表现出对废水中各种氮底物的出色代谢能力[50]。与途径I 中的细菌相比,途径II 中的细菌只代谢NH4+‑N,这阻碍了它们在实际污水处理中的应用[51]。因此,在低温和好氧条件下,良好的氮去除效能和较低的中间产物积累表明菌株P.veroniiDH‑3 是一种在净化含氮废水方面具有应用前景的脱氮菌剂。
4 结论
从河水底中分离筛选得到一株耐低温假单胞菌Pseudomonas veroniiDH‑3。10℃条件下,该菌培养48 h 后对NH4+‑N、NO2-‑N 和NO3-‑N 的去除率分别为99.07%、96.88%和90.30%,并且脱氮过程中没有检测到明显的NO2-‑N 积累。以NH4+‑N 和NO3-‑N为混合氮源时,48 h 后NH4+‑N 被完全去除,而NO3-‑N 的去除率为87.09%。该菌展现出良好的氮去除性能,推测其脱氮途径为硝化、反硝化作用和同化作用。
猜你喜欢
生物工程学报(2020年6期)2020-07-31
环境保护与循环经济(2017年7期)2018-01-22
中国酿造(2016年12期)2016-03-01
中国环境科学(2016年3期)2016-02-08
应用海洋学学报(2014年2期)2014-11-26
中国酿造(2014年9期)2014-03-11
食品工业科技(2014年9期)2014-03-11
植物营养与肥料学报(2011年6期)2011-10-24
环境工程技术学报(2011年2期)2011-10-14
海洋通报(2010年2期)2010-12-28