基于SSR 标记的广东黄皮种质资源遗传多样性分析及分子身份证构建
2024-01-12 12:41陆育生彭程常晓晓邱继水陈喆陈慧琼
生物技术通报 2023年12期
陆育生 彭程 常晓晓 邱继水 陈喆 陈慧琼
(广东省农业科学院果树研究所 农业农村部南亚热带果树生物学与遗传资源利用重点实验室 广东省热带亚热带果树研究重点实验室,广州 510640)
黄皮(Clausena lansium(Lour.)Skeels)属芸香科(Rutaceae)柑橘亚科(Aurantioideae)黄皮属(ClausenaBurm. f.),是多年生小乔木、热带亚热带常绿果树[1],原产于我国南部,主要分布在广东、广西、福建、海南、云南以及我国台湾等地[2]。黄皮果实外观独特,风味酸甜可口,更具有止咳化痰、消暑健胃、抗氧化、抑肿瘤、防老年痴呆等功效[3-6]。近年来,荔枝(Litchi chinensisSonn.)、龙眼(Dimocarpus longanLour.)等大宗水果价格持续低迷,黄皮的售价相对喜人,给果农带来了较高的经济收益,栽培面积持续扩大,已成为华南地区农业供给侧结构性改革和实施乡村振兴战略的重要经济树种。广东省是中国最重要的黄皮产区之一,种植面积和产量均居全国首位,其中,仅云浮市郁南县栽培面积就超过1 万hm2,占全国总面积一半以上,并建成我国首个黄皮现代农业产业园,初步形成了涵盖种植、加工、科技、流通、销售、文旅、餐饮的全产业链条,所产果品成功走出国门,远销北美、中东等国家(http://dara.gd.gov.cn/zfjy/content/post_3979099.html)。
但是黄皮作为小宗果树,育种工作长期未受到足够重视,导致目前主栽品种单一且口感不尽如人意,例如栽培面积最大的无核黄皮虽然可食率高,但风味偏酸,无法满足消费者对高品质水果的需求,而从城甜黄皮虽然口感偏甜,但缺乏风味且易于引起腻感,同样对消费者的消费热情产生了不利影响。近年来,伴随着消费升级,消费者对高品质水果的需求日益高涨,黄皮品种的改良与创新已刻不容缓。广东黄皮栽培历史悠久,加之民间习惯屋前房后实生种植,经过长期的自然变异和人工选择,形成了丰富的种质资源和各具特色的地方品种,可为黄皮遗传改良和新品种培育提供广阔的遗传基础。然而,由于黄皮为小宗果树,种质资源研究基础薄弱,缺乏系统的分类鉴定,其庞大的种质资源易造成种质混乱。特别是随着黄皮产业的不断壮大,各地相互引种和种质交流频繁发生,导致“同名异物”或“同物异名”现象屡见不鲜。这不仅给黄皮种质资源的保护、管理和开发利用带来很大困难,而且也极大损害育种者和种植者的权益。因此,系统开展广东黄皮种质资源的遗传多样性和亲缘关系研究,有利于准确区分和鉴定黄皮种质,对于黄皮产业的健康发展具有重要意义。
种质资源鉴定评价的传统手段是采用形态学标记,其具有简单和直观的优点,但其易受环境及植物生长发育状态等因素的影响,而且误差大、耗时、费力,因而存在较大的局限性[7]。随着分子生物学技术的快速发展,DNA 分子标记作为新兴的标记技术,能很好地弥补形态标记的不足,已被广泛应用于植物种质资源鉴定研究。SSR(simple sequence repeat),又称微卫星DNA(microsatellites),是由1-6个核苷酸为重复单元组成的简单串联重复序列[8],重复单元数量的差异造成了种质资源间丰富的遗传多态性。作为第二代分子标记的代表,SSR 分子标记具有技术简便、多态性丰富、稳定性高、重复性好、共显性等优点[9-10],被国际植物新品种保护联盟(International Union for the Protection of New Varieties of Plants, UPOV)确定为植物新品种保护应用最广泛的分子标记体系[11]。
目前,SSR 标记已广泛应用于苹果(Malus pumilaMill.)[12-14]、梨(Pyrusspp.)[15-17]、桃(Prunus persica(L.)Batsch)[18-20]、 李(Prunus salicinaLindl.)[21]、板栗(Castanea mollissimaBlume)[22]、柑橘(Citrus reticulataBlanco)[23]、葡萄(Vitis vin‑iferaL.)[24-25]、荔枝[26]、火龙果(Hylocereus undatu‘Foo‑Lon’)[27]等果树的遗传多样性分析、核心种质筛选、遗传图谱构建以及种质鉴定等诸多领域,但关于SSR 标记技术在黄皮种质资源的多样性分析和分子身份证构建方面的研究尚未见报道。
本研究以农业农村部广州黄皮种质资源圃保存的84 份广东黄皮种质为材料,利用项目团队前期开发的12 对SSR 引物开展遗传多样性分析和分子身份证构建,旨在揭示广东黄皮种质资源的遗传多样性水平及亲缘关系,从而为黄皮种质资源的合理保护和高效利用提供参考依据。
1 材料与方法
1.1 材料
参试的黄皮种质资源共84 份,包括已审定的选育品种、具有一定种植规模的农家品种以及从全省各地收集的地方种质(实生单株)(表1),以上所有种质资源均保存于广东省农业科学院果树研究所农业农村部广州黄皮种质资源圃内。每份种质材料选取新鲜健康的嫩叶1 g,液氮速冻后于‑80℃冰箱保存备用。
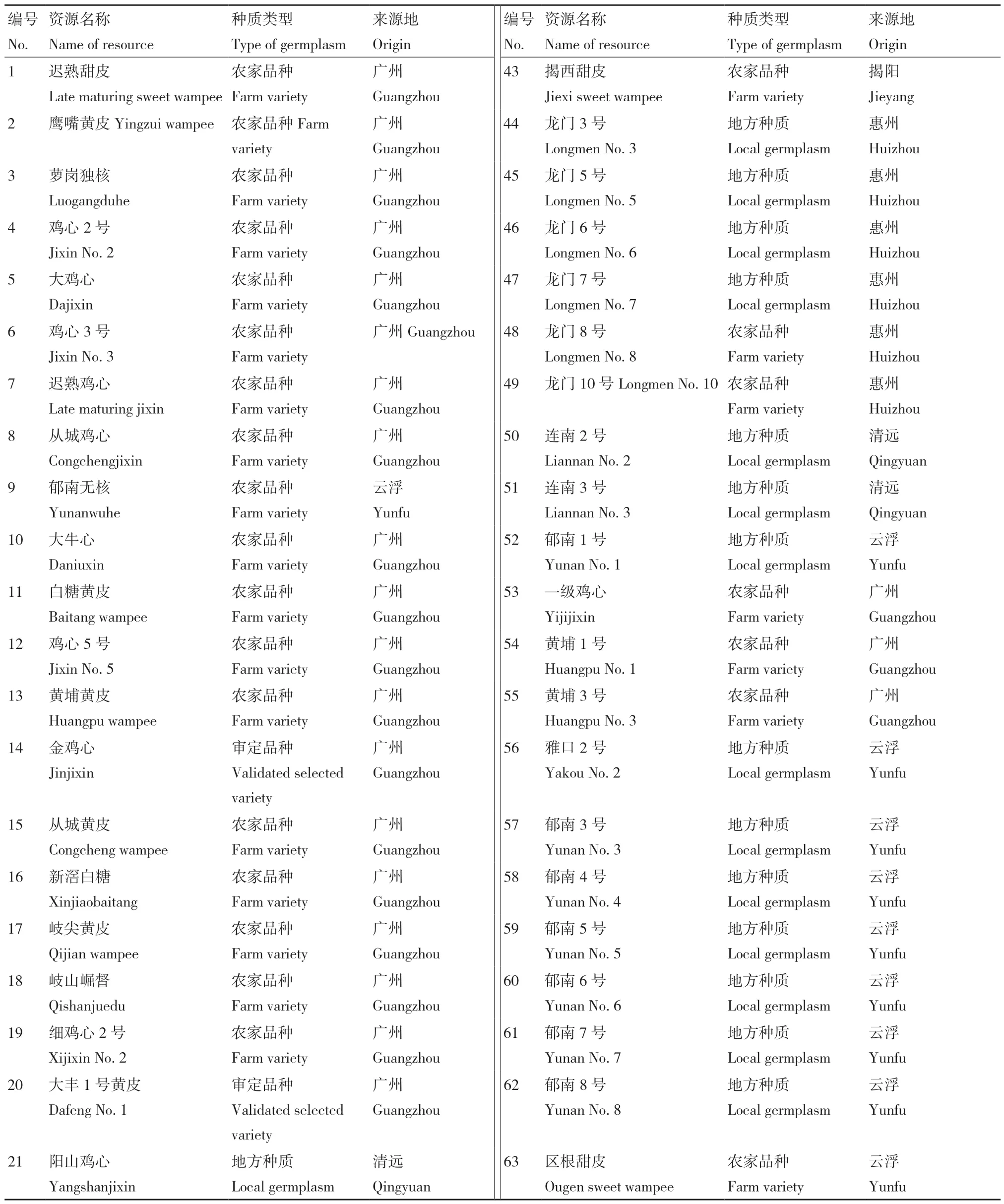
表1 84 份供试广东黄皮种质资源Table 1 84 germplasm resources of wampee in Guangdong province
1.2 方法
1.2.1 基因组DNA 提取与检测 采用高效植物基因组DNA 提取试剂盒(DP350,天根生化科技有限公司,中国北京)提取黄皮叶片基因组DNA,所得产物用1%琼脂糖凝胶电泳和Nanodrop 2000 微量紫外分光光度计(Thermo Scientific, USA)进行DNA 质量和浓度检测,然后用灭菌ddH2O 将各样品DNA浓度均稀释至20 ng/μL 备用。
1.2.2 PCR 反应及产物的毛细管电泳检测 试验所用的SSR 引物为项目团队前期从黄皮转录组序列开发[28]获得的12 对多态性引物(表2)。所有引物由生工生物工程(上海)股份有限公司合成,在上游引物的5'端加1 个荧光素标记(carboxyfluorescein,FAM)。PCR 扩增反应体系20.0 μL,包括2× Taq PCR mix 10.0 μL、5 μmol/L 正、反向引物各1.0 μL、20 ng/μL DNA 模板1.0 μL 和ddH2O 7.0 μL。扩增程序为94℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 60 s,35 个循环;72℃ 10 min,4℃保存。PCR 产物稀释10 倍,取1 μL 加入96 孔板中,再加入甲酰胺和分子量内标GeneScan 500 LIZ(Thermo Fisher Scientific,USA)混合液(9.5∶0.5,体积分数)9 μL,95℃ 5 min,然后迅速冰上冷却10 min,3 000 r/min 离心1 min,在ABI 3730 上进行自动荧光检测。

表2 供试SSR 引物信息Table 2 Information of SSR primers used in this study
1.2.3 数据处理与分析 电泳结束后,利用软件GeneMapper 4.0 对原始数据进行分析,获得每对引物在不同样品上的扩增片段;采用Powermarker 3.25[29]和Popgene 32[30]计算等位基因数(Na)、有效等位基因数(Ne)、观测杂合度(Ho)、期望杂合度(He)、多态性信息量指数(PIC)和Shannon信息指数(I);使用Power Marker 3.25 计算各资源间的Nei's 遗传距离,随后基于非加权分组算术平均 法(unweighted pair‑group method with arithmetic means, UPGMA)在FigTree 1.4.3 中构建聚类树;使用Structure 2.3.4[31]基于贝叶斯聚类方法分析黄皮种质资源的群体结构,设不作数迭代(length of burnin period)开始时的马尔科夫链蒙特卡洛(MCMC)为10 000 次,不作数迭代后的MCMC(mumber of MCMC reps burnin)为100 000 次,采用混合祖先模型(admixture model)和等位变异发生频率相关模型(allele frequency correlated model),群体数目(K)设为1-10,对每个K值模拟运算10 次,运算结果以Zip 格式进行压缩,提交给在线软件Structure harvester(http://taylor0.biology.ucla.edu/struct_harvest/),以判断群体的Clusters 数(即最可能的K值)。
1.2.4 分子身份证构建 根据12 对引物在各样品中扩增的带型,按分子量将条带从小到大排序,每一种带型用个位阿拉伯数字1-9 编码表示,带型数超过9 的用小写英文字母a-z 依次表示,将每个样品在12 对引物上的扩增带型数据串联起来,即得到每个样品以12 位数字或字母表示的分子身份证。
2 结果
2.1 广东黄皮种质资源SSR位点多态性分析
利用12 对SSR 引物对84 份广东黄皮种质资源进行扩增和荧光毛细管电泳检测,并计算获得各遗传多样性参数(表3)。12 个SSR 位点共检测到43 个等位基因(Na),每对引物检测到的等位基因为3-6 个,平均等位基因3.583 个。有效等位基因(Ne)为1.416-2.984,平均值为2.187。观测杂合度(Ho)和期望杂合度(He)变化范围分别为0.265(CP006)-0.675(CP010)和0.294(CP006)-0.665(CP007),平均值分别为0.497 和0.524。Shannon 信息指数(I)变化范围为0.567(CP006)-1.096(CP006),平均I为0.913。各引物的多态性信息含量(PIC)分布范围为0.291-0.642,平均PIC值为0.484,其中CP010 最大,CP006 最小。12 个SSR 位点的PIC值均大于0.25,其中6 个位点的PIC值大于0.50。
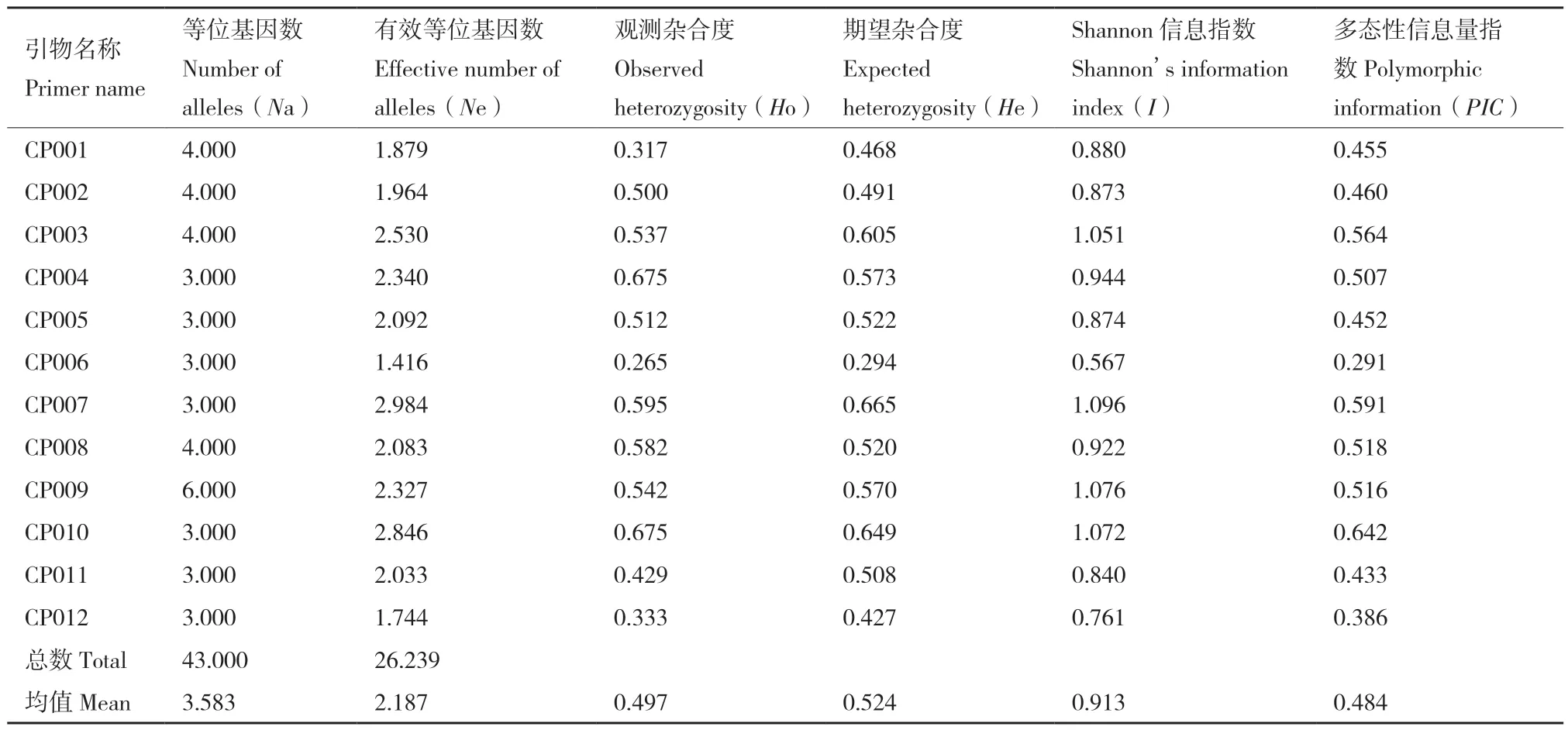
表3 12 对SSR 引物的遗传多样性Table 3 Genetic diversity at 12 pairs of SSR markers
2.2 聚类分析
根据12 对SSR 引物的扩增结果,利用Power Marker 软件对84 份广东黄皮种质资源进行基于UPGMA 方法的聚类分析。结果(图1,表4)表明,全部资源可以划分为6 个类群,第I 类群包含13 份种质,除了‘阳山鸡心’外,其他种质均来自广东西部地区的湛江和云浮。第II 类群包含8 份种质,均来自广东东部地区的梅州和潮汕。第III 类群包含11 份种质,除了‘阳山大叶’‘连南2 号’‘连南3号’‘龙门9 号’外,其他种质均来自广东西部地区的云浮和阳江。第IV 类群包含6 份种质,除了‘始兴032’外,其他种质均来自广东中部地区的惠州。第V 类和第VI 类为培育品种和农家品种,来源地分散在广东各地,其中,第V 类包含30 份种质,除‘番禺甜皮’‘白糖黄皮’‘萝岗独核’‘小果甜皮’‘塔下甜皮’外,其他均为鸡心黄皮类,特征是果实均为圆卵形或椭圆形、肉质细嫩、风味甜酸适中。‘潮州11 号’‘从城鸡心’‘鸡心5 号’‘揭阳鸡心’完全聚在一起,推测这4 份种质为同物异名种质。第VI 类群包含16 份种质,除‘龙门1 号’‘龙门10 号’‘龙门8 号’外,其他均为甜黄皮类,特征表现为果皮浅黄色、肉质脆嫩、风味清甜、无酸味。
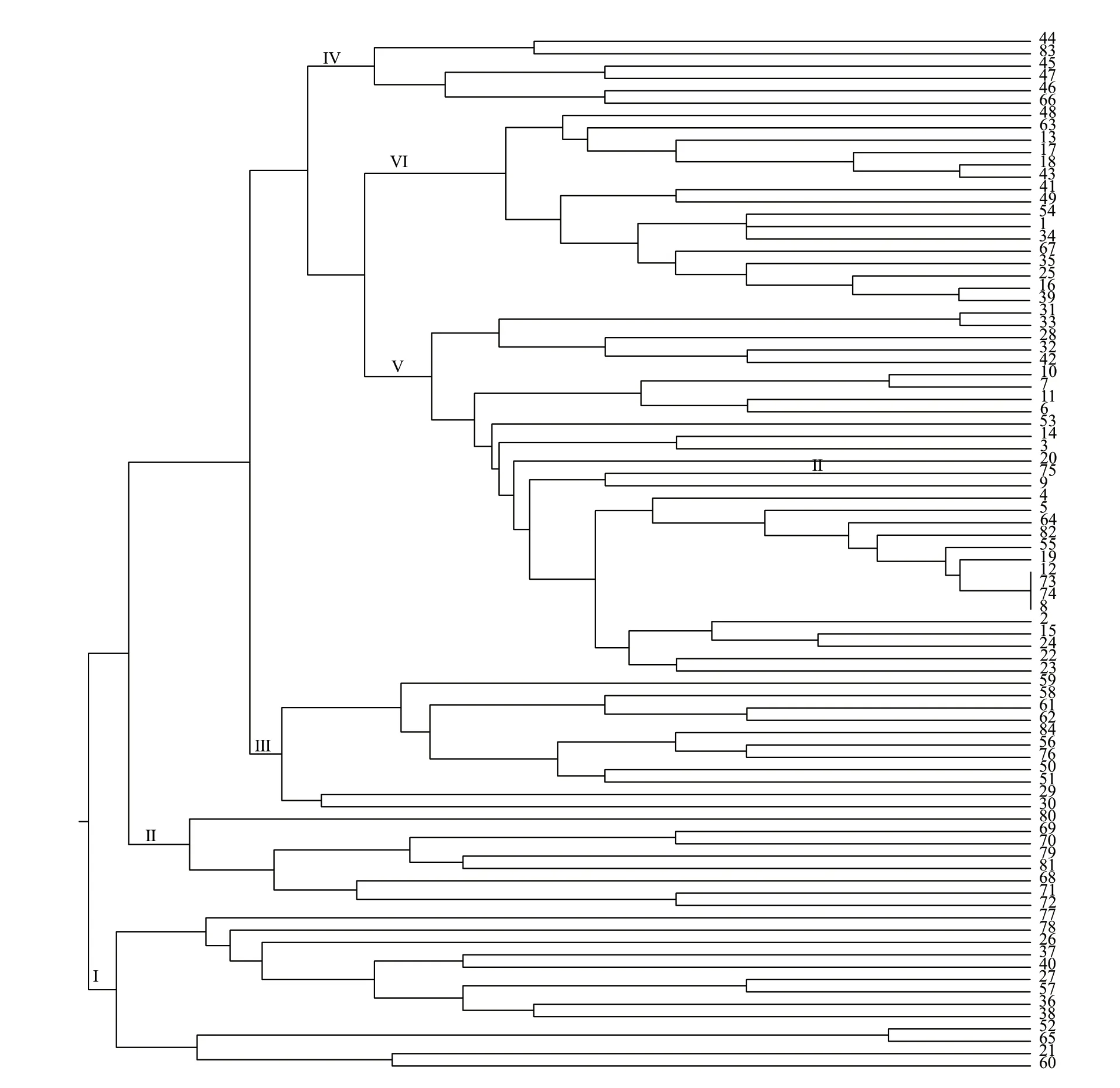
图1 基于SSR 标记的84 份种质资源聚类图Fig. 1 Cluster diagram of 84 wampee germplasm resources based on SSR markers
2.3 遗传结构分析
进一步利用structure 软件构建广东黄皮种质资源的群体居群遗传结构图,结果显示,84 份种质在K=2 时,ΔK有最大值(图2),此时84 个样本可分为2 个亚群(图3)。亚群A 包含39 份资源,除了来自云浮的‘区根甜皮’、广州的‘禄田独核’和惠州的‘龙门10 号’等农家品种外,其他种质均为来源于全省各地的地方种质;亚群B 包含45 份资源,除了地方种质‘始兴032’和‘龙门7 号’外,其他种质均为选育品种和农家品种,结果与UPGMA聚类分析结果一致。84 份材料中有79 份材料在某一类群的同系得分(Q 值)>0.6,占94.05%,表明供试的黄皮种质遗传结构较为简单,但群体间仍有少量种质混杂,存在相互的基因渗透。
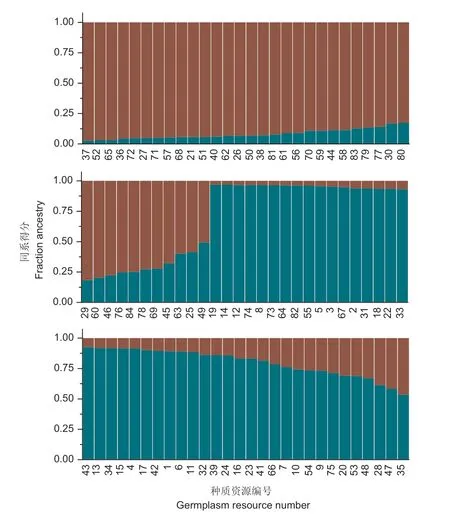
图3 K=2 时基于SSR 标记的84 份黄皮种质群体遗传结构Fig. 3 Genetic structure of 84 wampee germplasm and population at K=2 based on SSR markers
2.4 分子身份证构建
按表3 引物的顺序将所有引物在样品中扩增出来的基因型(带型)的分子量大小进行赋值排序(表5),串联排列后获得供试84 份黄皮资源的分子身份证代码(表6),在同一位置代码相同则表示基因型相同。其中,推测为同一种质的‘潮州11 号’‘从城鸡心’‘鸡心5 号’‘揭阳鸡心’具有相同的分子身份代码,其余种质均具有专属的分子身份代码。
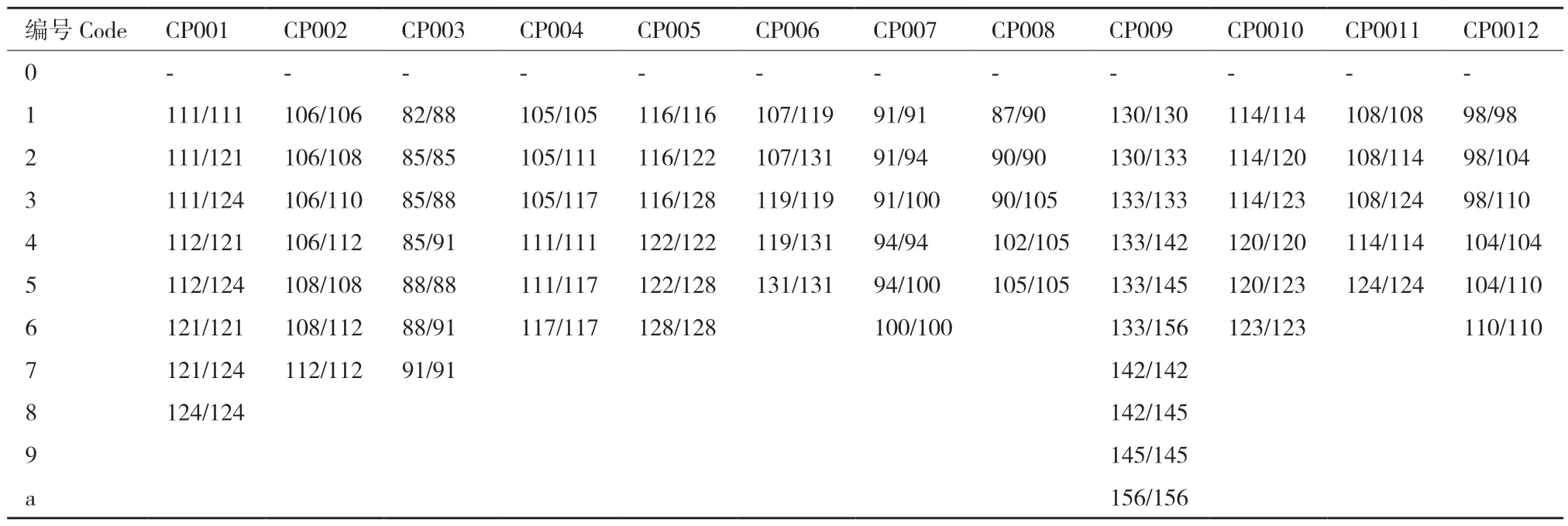
表5 12 对SSR 引物的扩增带型编号Table 5 Amplified band codes of 12 pairs SSR primers
3 讨论
遗传多样性是生命系统的基本特征,是物种适应自然和发生进化的产物,开展遗传多样性分析对种质资源的有效保护、开发和利用具有重要意义。目前,关于DNA 分子标记在黄皮种质资源遗传多样性分析中的应用仅有少数报道,刘小梅等[32]利用10 对RAPD(random amplified polymorphic DNA)标记对来自广东、广西和海南的36 份材料进行研究,结果表明上述黄皮种质间的亲缘关系较近。李开拓等[33]利用15 对ISSR(inter‑simple sequence repeat)引物对来源于福建、广东和广西的40 个黄皮品种(品系)进行遗传多样性分析,结果也表明黄皮品种(品系)间的遗传相似性较大。本研究在前期基于转录组序列的黄皮SSR 标记挖掘的基础上[28],首次利用12 对SSR 荧光引物对来源于广东的84 份黄皮种质资源进行基因分型和遗传多样性分析。多态性信息含量(PIC)和期望杂合度(He)是衡量种群遗传多样性的重要指标,PIC反映SSR 位点的变异程度,当PIC≥0.5 时表示位点具有高度多态性,当0.25 根据SSR 分型数据,采用UPGMA 方法进行聚类分析。结果表明,供试的84 份广东黄皮种质资源可分为6 个类群,第I-IV 类群为地方种质,其中,第I 和第III 类群大部分种质来自广东西部地区,第II 类群主要来自广东东部地区,第IV 类群主要来自广东中部地区;而第V 类和第VI 类则为具有一定栽培规模的培育品种和农家品种,来源分布于广东各地。可见,基于UPGMA 的聚类结果与地理分布有一定关联,但并没有严格按照材料的地理来源进行聚类。关于黄皮种质资源的分类目前尚未有统一标准,传统习惯按单一形态或园艺性状进行简单分类,如按果实风味分为甜黄皮和酸黄皮,按果实形状分为鸡心黄皮、圆果黄皮、牛心黄皮、梨形黄皮,按种子的多少分为有核黄皮和无核黄皮等[32]。本研究结果表明,基于SSR 的分类与传统形态分类具有一定的相关性,如第V 类群中大部分种质为鸡心黄皮类种质,其主要特征是果实均为圆卵形或椭圆形,肉质细嫩、风味甜酸适中;而第VI 类群主要为甜黄皮类,形态特征是果皮浅黄色、肉质脆嫩、风味清甜、无酸味。然而传统形态分类范围过宽,且性状评价容易受主观和环境因素干扰,难以准确反映种质资源间的亲缘进化关系;与之相比,基于SSR 的分类是从DNA 水平揭示种质资源间的差异,能更全面、准确地反映种质间的亲缘关系,其结果更加严谨、可靠。 分子身份证是在DNA 指纹图谱的基础上,采用一定编码规则对电泳图谱进行数字化处理,从而获得字符串形式表示的身份代码;其克服了指纹图谱人工比对的繁琐、低效等问题,能简单、直观地区分不同种质,被广泛应用于种质资源鉴定和品种识别。目前,基于SSR 标记技术已成功构建了葡萄[24]、苹果[35]、梨[36]、火龙果[27]、山楂[7]等多种果树种质资源的分子身份证。本研究利用12 对SSR 荧光引物对84 份黄皮种质进行扩增,获得指纹图谱,并采用基因型赋值的方式编码,得到了供试材料的分子身份证。除了推测为同一种质的‘潮州11 号’‘从城鸡心’‘鸡心5 号’‘揭阳鸡心’外,其余种质均具有唯一的分子身份证,可被有效区分,表明SSR标记技术作为一种有效的技术手段,可用于黄皮种质资源的分子身份证构建。 广东黄皮种质资源具有较为丰富的遗传多样性,基于UPGMA 方法可将供试材料分为6 个类群,与传统形态分类相比,SSR 聚类结果更严格、准确;基于12 对SSR 引物的分型结果,采用数字加英文字母的编码方式,成功构建黄皮种质资源的分子身份证。4 结论
猜你喜欢
中国果业信息(2023年9期)2023-12-17
河北果树(2020年4期)2020-11-26
小天使·三年级语数英综合(2019年9期)2019-11-09
创新作文(1-2年级)(2019年4期)2019-10-15
江苏农业科学(2019年5期)2019-09-02
小猕猴学习画刊(2019年5期)2019-06-21
西江月(2018年9期)2018-09-12
广东农业科学(2017年5期)2017-08-29
中成药(2017年6期)2017-06-13
应用海洋学学报(2014年1期)2014-11-22