香蕉MaMC6 的克隆及原核表达分析
2024-01-12 12:41滕梦鑫徐亚何静汪奇乔飞李敬阳李新国
生物技术通报 2023年12期
滕梦鑫 徐亚 何静 汪奇 乔飞 李敬阳 李新国
(1. 热带作物生物育种全国重点实验室 海南大学热带农林学院,海口570228;2. 中国热带农业科学院热带作物品种资源研究所,海口571101)
程序性细胞死亡(programmed cell death, PCD)在植物的生长发育中发挥着重要的作用,是植物抵抗生物和非生物胁迫的基本机制[1-2]。多项研究表明,动物和植物细胞[Ca2+]cyt的升高是诱导PCD发生的必要条件。Ca2+可通过增加细胞内钙水平导致液泡破裂[3];直接激活钙依赖的核酸内切酶,使DNA 发生降解[4-5];影响线粒体通透性转变,促进线粒体释放出细胞色素c,激活下游类caspase 3 等方式参与PCD[6]。
Metacaspases(MCs)属于半胱氨酸蛋白酶类[7],在植物PCD 过程中起着关键的调控作用。根据空间结构域和氨基酸序列的相似性,植物 MCs 分为两个亚类:I 型MCs 的N 端前结构域包含富含脯氨酸的重复基序,并且在植物成员中还有一个锌指基序。II 型MCs 不含有这样的前结构域[8-9]。但MCs 缺乏caspases 活性,并且不负责在植物细胞死亡期间检测到的类caspase 活性[10],另有研究表明MCs 的表达可能导致下游具有类caspase 活性的蛋白酶激活[11],启动PCD 的发生。MCs 在拟南芥中研究较多,AtMC1、AtMC2、AtMC8和AtMC9等可正向或反向调控PCD 的发生[12-14],且多个成员受生物和非生物胁迫诱导表达[15-17]。水稻OsMCs家族成员在响应非生物和生物胁迫下也有不同的表达模式,其中OsMC1和OsMC7受盐胁迫诱导上调表达[18-19]。
香蕉产业已成为热带高效农业的支柱,海南省因自然资源条件适宜,是中国香蕉主产区之一[20-21]。香蕉对环境的变化极其敏感[22],由于生产上施肥不当和海水倒灌导致的土壤盐碱化,以及发展海水灌溉农业的需要[23],使得对香蕉耐盐的研究变得十分重要。在前期研究中我们发现,不同苗龄和组织部位对胁迫的响应存在特异性[24-25],而悬浮细胞同质性高[26],可以对外部刺激做出快速而一致的反应,是一个对研究有价值的简单和可控的系统,因此被广泛用于大麦(Hordeum vulgareL.)、小麦(Triticum aestivumL.)、拟南芥(Arabidopsis thaliana)和柑橘(Citrus reticulataBlanco)等植物对胁迫响应的生理和分子机制研究[27-30]。
前期研究中我们发现MaMC6可对盐胁迫做出快速的响应[31],但该基因的表达在盐胁迫下是否起着积极的调控作用有待进一步研究。本实验从巴西蕉(MusaAAA Cavendish cv. Brazil)细胞中克隆到MaMC6基因,然后利用生物信息学和分子生物学等手段对其进行鉴定及分析。检测盐胁迫以及不同钙效应剂对MaMC6表达量的影响,转大肠杆菌验证基因功能,为丰富香蕉耐盐机制提供参考。
1 材料与方法
1.1 材料
材料来源于中国热带农业科学院热带作物品种资源研究所保存的巴西蕉悬浮细胞系,继代周期为7 d,培养条件为:温度26℃,光照时间16 h/d,转速150 r/min。
盐胁迫及钙效应剂处理:分别用100 mmol/L NaCl、100 mmol/L NaCl + 10 mmol/L CaCl2、100 mmol/L NaCl + 10 mmol/L EGTA 和100 mmol/L NaCl +5 mmol/L LaCl3处理继代后第5 天的巴西蕉细胞,并在处理后的0、2、4、8、16、24 h 进行取样。取1.8 mL 悬浮细胞于2 mL 离心管中,25℃、8 000×g离心6 min,去除培养基,液氮速冻,置于‑80℃保存备用。每组处理取3 次重复。
烟草材料为30 d 苗龄的野生型本氏烟(Nicoti‑ana benthamianaL.)。
1.2 方法
1.2.1MaMC6基因的克隆 根据MaMC6的CDS序列,设计带有NcoI‑ 和SpeI‑酶切位点的引物1302‑MaMC6‑F/R(表1),以巴西蕉细胞cDNA 为模板,用PrimeSTAR Max DNA 聚合酶扩增亚细胞定位所需的特异性目的片段,将MaMC6连接到pCAMBIA1302‑GFP 载体上;用带有NcoI‑ 和XhoI‑酶切位点的引物pET28a‑MaMC6‑F/R 扩增构建原核表达载体所需的特异性目的片段,将MaMC6连接到pET28a 载体上。连接后的质粒转化大肠杆菌感受态DH5α,随机挑取单克隆进行培养,菌液PCR 验证,阳性单克隆菌液送至上海生工公司测序鉴定。

表1 试验所用引物Table 1 Primers used in the experiment
1.2.2 生物信息学分析 从香蕉基因组数据库(https://banana‑genome‑hub.southgreen.fr/) 中 检 索获得MaMC6 碱基序列、蛋白序列和上游2 000 bp启动子序列;使用ProtParam(https://web.expasy.org/protparam/)对蛋白理化性质进行预测;使用TMHMM(https://services.healthtech.dtu.dk/service.php?TMHMM‑2.0)进行蛋白跨膜结构预测;利用NCBI(https://www.ncbi.nlm.nih.gov/)检索MaMC6 的氨基酸同源序列,然后使用DNAMAN 进行多序列比对,MEGA‑X 构建系统进化树;通过PredictProtein(https://predictprotein.org/)进行蛋白二级结构预测,SWISS‑MODEL(https://swissmodel.expasy.org/interactive)进行蛋白三级结构预测;通过PlantCARE(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行顺式作用元件预测,并使用TBtools 对其进行可视化。
1.2.3MaMC6的表达分析 使用TIANGEN 多糖多酚植物总RNA 提取试剂盒,提取巴西蕉细胞总RNA。诺唯赞反转录试剂盒说明书,将总RNA 反转录为cDNA,反转录产物在‑20℃下保存备用。实时荧光定量PCR(RT‑qPCR),使用 QuantStudio 6 Flex实时荧光定量 PCR 仪,以香蕉A 基因组Actin为内参基因,用目标引物进行PCR 扩增(表1),对目的基因进行特异性表达分析。反应体系(10 μL),反应程序:50℃ 2 min,95℃ 10 min,95℃ 15 s,60℃1 min,95℃ 15 s,60℃ 1 min,95℃ 15 s,共40 个循环,反应结束后将结果导入Excel 软件,采用 2-ΔΔCt法计算相对表达量[25]。
1.2.4 亚细胞定位观察 采用在烟草叶片中瞬时过量表达的方法,观察MaMC6 在细胞内定位情况。从测序正确的单克隆中提取pCAMBIA1302‑MaMC6‑GFP 质粒,转化农杆菌感受态LBA4404。以转入pCAMBIA1302‑GFP 质粒的LBA4404 为对照,注射烟草叶片下表皮,黑暗培养1 d,光照培养2 d 后,制片,在激光共聚焦显微镜下观察并拍照记录。
1.2.5MaMC6 的诱导表达及在大肠杆菌BL21 中的抗性分析 从测序正确的单克隆中提取pET28a‑MaMC6质粒,转化大肠杆菌感受态BL21。分别将含有重组质粒pET28a‑MaMC6和空载质粒pET28a的大肠杆菌BL21 使用IPTG 诱导表达,再将诱导后的菌液按梯度稀释为1、10-1、10-2、10-3、10-4的浓度。稀释后的菌液各取4 μL 滴在含有800 mmol/L NaCl 和800 mmol/L 甘露醇的LB 固体培养基上培养,置于37℃培养进行盐胁迫和渗透胁迫试验,另一组置于50℃培养进行高温胁迫试验,以正常培养为对照组,12 h 后观察重组菌和对照菌生长存活情况并拍照。
2 结果
2.1 MaMC6的克隆
MaMC6由963 个碱基组成,凝胶电泳结果如下图1 所示。扩增出接近1 000 bp 的特异性条带,与预期大小相近。

图1 MaMC6 凝胶电泳图Fig. 1 MaMC6 gel electrophoresis image
2.2 MaMC6生物信息学分析
MaMC6 由320 个氨基酸组成,分子质量约为34.21 kD,等电点为5.99,不稳定系数为35.43 属于稳定蛋白,脂肪系数为85.97,亲水平均系数为‑0.051属于亲水蛋白,不含有跨膜结构。
序列多重比较结果显示MaMC6 与其他几种植物氨基酸序列有较高的相似性,保守结构域与拟南芥metacaspase‑9 一致含有一个caspase‑like domain,同属于II 型metacaspase(图2)。

图2 MaMC6 与其他植物氨基酸序列多重对比Fig. 2 Multiple comparison of amino acid sequences between MaMC6 and other plants
对包含MaMC6 在内的20 个同源氨基酸序列进行对比,使用MEGA‑X 构建系统进化树。图3 所示,Mametacaspase‑9 与Mbmetacaspase‑9 有高度的同源性,其次是Zometacaspase‑9‑like 和OsZIFL1(Zinc Induced Facilitator‑Like 1),与Lsmetacaspase‑9 同源性较低。

图3 MaMC6 与其他植物的系统进化树分析Fig. 3 Phylogenetic tree analysis of MaMC6 and other plants
对MaMC6 蛋白二级结构进行预测,MaMC6 含有28.12%的α-螺旋、11.25%的β-折叠和60.62%的无规则卷曲。三级结构预测如图4 所示。
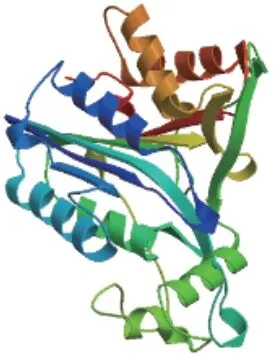
图4 MaMC6 蛋白三级结构预测Fig. 4 Prediction of the 3-three-dimensional structure of MaMC6 protein
MaMC6顺式作用元件分析(图5)表明其含有较多的光响应元件,另外还含有防御和应激、玉米素代谢调控、茉莉酸甲酯、赤霉素、脱落酸等激素响应元件。其中防御和应激表明该基因可能受外界胁迫诱导表达,参与香蕉的抗逆。
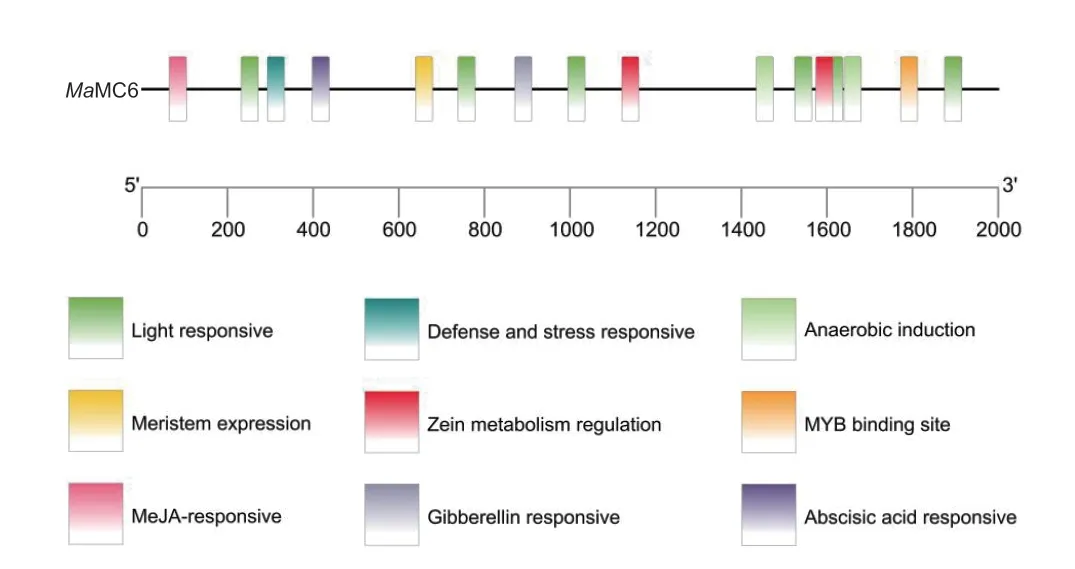
图5 MaMC6 顺式作用元件分析Fig. 5 Analysis of cis-acting elements of MaMC6
2.3 MaMC6的表达分析
2.3.1 盐胁迫下MaMC6表达分析 经100 mmol/L NaCl 处理后,以0 h 为对照组,巴西蕉MaMC6的RT‑qPCR 结果(图6)可知,在2 h 时出现显著上调表达,相对表达量为对照的7.70 倍,随后表达量开始下降,到24 h 时降至对照的31%。
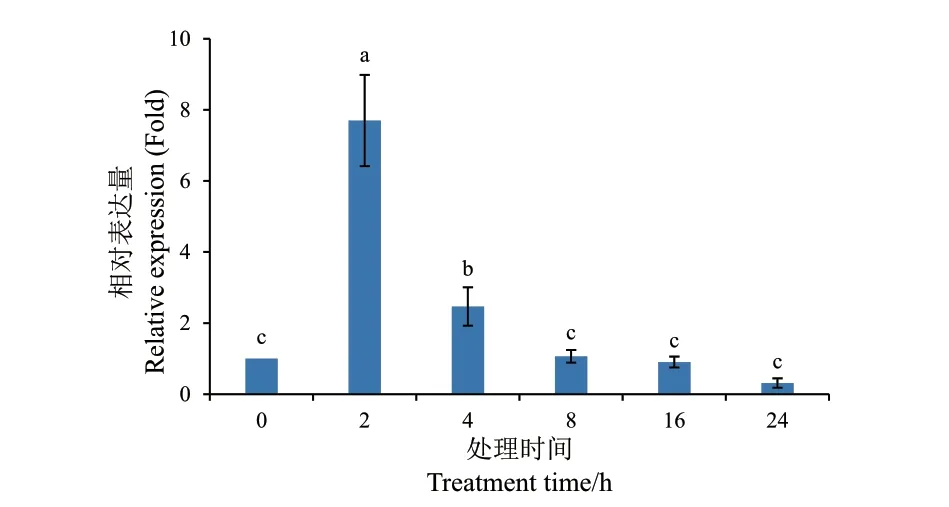
图6 MaMC6 在NaCl 处理下的基因表达Fig. 6 Gene expression of MaMC6 under NaCl treatment
2.3.2 盐胁迫下不同钙效应剂处理后MaMC6的表达分析 从图7 看出,CaCl2处理后,基因的表达受到抑制,但仍在2 h 时达到最高峰,为对照的3.99 倍。而钙离子螯合剂EGTA 和钙离子通道阻碍剂LaCl3提高了MaMC6在盐胁迫下的相对表达量,处理期间相对表达量均高于NaCl 处理组,但整体变化趋势相似,在2 h 时达到最高峰,分别为对照的9.92 和11.20 倍。由此推测,适当添加外源Ca2+可以通过抑制PCD 相关基因的表达,进而缓解由NaCl 诱导的PCD 的发生,通过EGTA 和LaCl3阻碍外源Ca2+进入胞内参与响应盐胁迫的调控则会加速PCD的发生。该基因的表达受到外源Ca2+的影响。
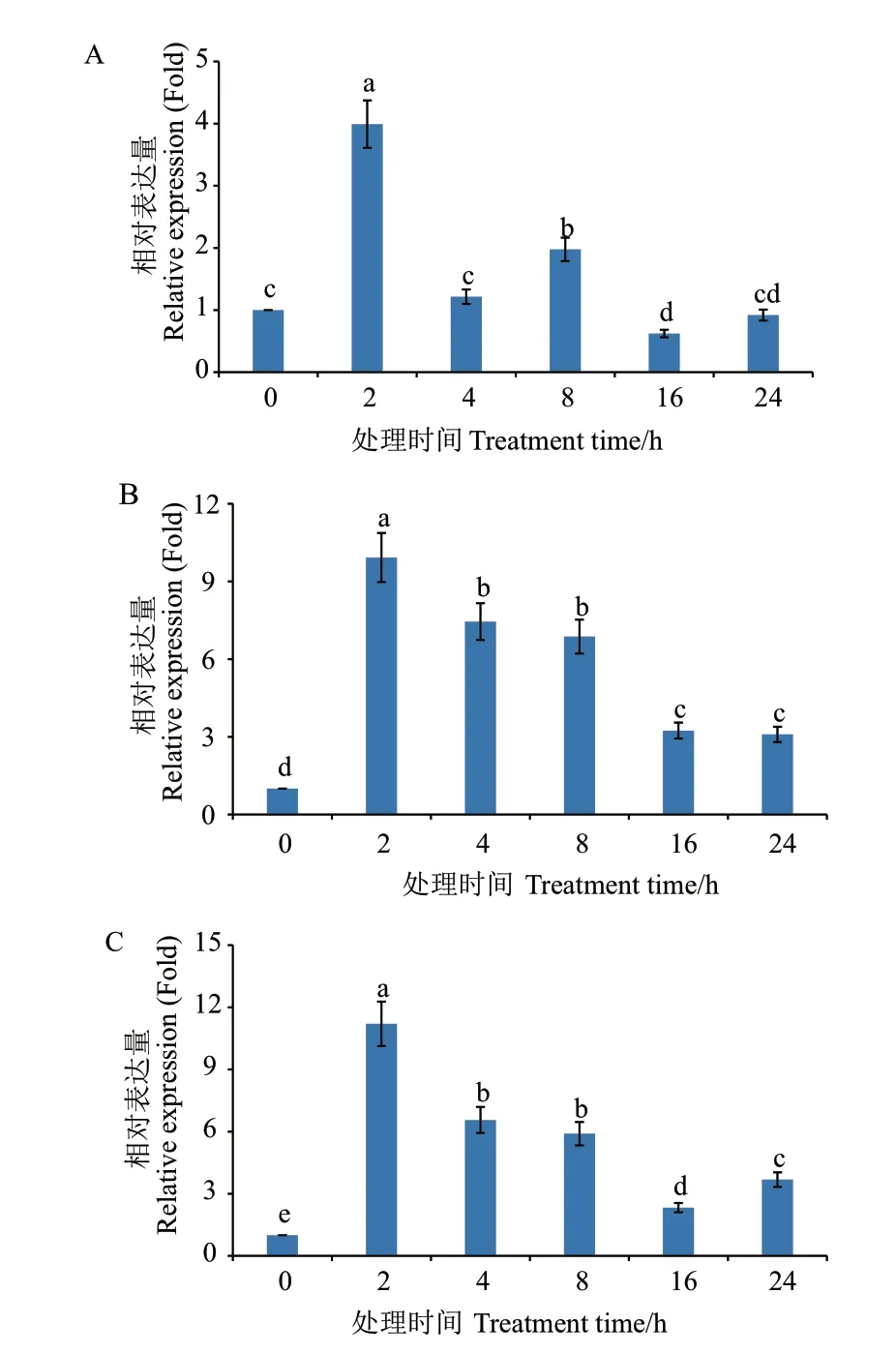
图7 不同钙效应剂对盐胁迫下MaMC6 表达的影响Fig. 7 Effects of different calcium effectors on the expression of MaMC6 under salt stress
2.4 MaMC6亚细胞定位分析
瞬时转化烟草叶片细胞后,在激光共聚焦显微镜下观察细胞核和细胞膜上有GFP 荧光信号,表明MaMC6‑GFP 融合蛋白呈现出细胞质和细胞核定位模式(图8)。
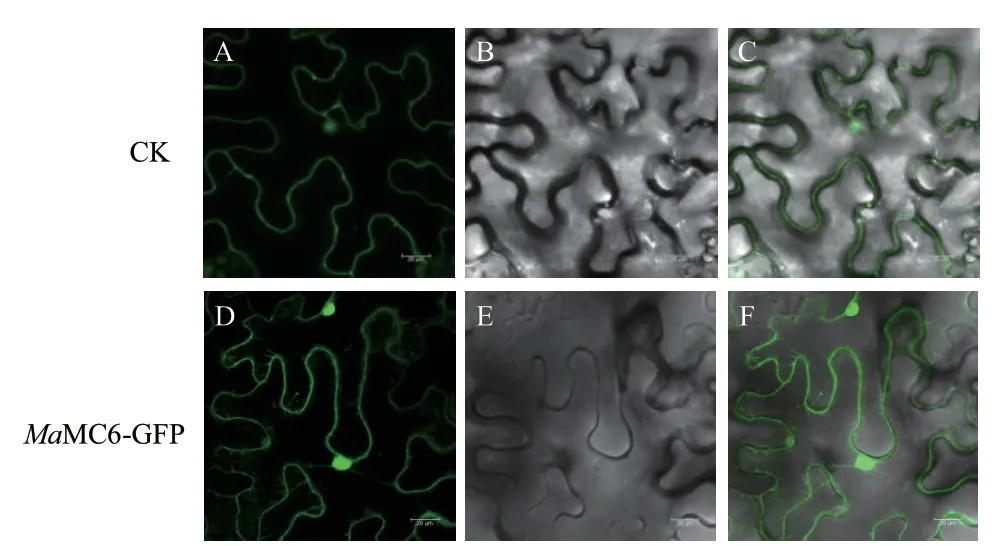
图8 MaMC6 亚细胞定位Fig. 8 Subcellular localization of MaMC6
2.5 MaMC6在BL21中对非生物胁迫的响应
从图9 可知,在LB 培养基上的对照菌和重组菌的生长情况没有明显差别。而在含有800 mmol/L NaCl 和800 mmol/L 甘露醇的LB 培养基上,菌落生长情况在菌液稀释到10-2梯度时已经出现明显差异,表达MaMC6 蛋白的重组菌的生长情况要优于对照菌。高温胁迫下培养的结果显示,pET28a‑MaMC6菌落生长情况在菌液稀释到10-2梯度时开始出现差异,此时重组菌的生长情况要优于对照菌,到10-3梯度时高温基本抑制了含有空载pET28a 质粒的对照菌的生长,而此时表达MaMC6 蛋白的菌落仍具有活力。以上结果表明,MaMC6的表达能提高大肠杆菌BL21 的耐盐、耐渗透和耐高温能力。
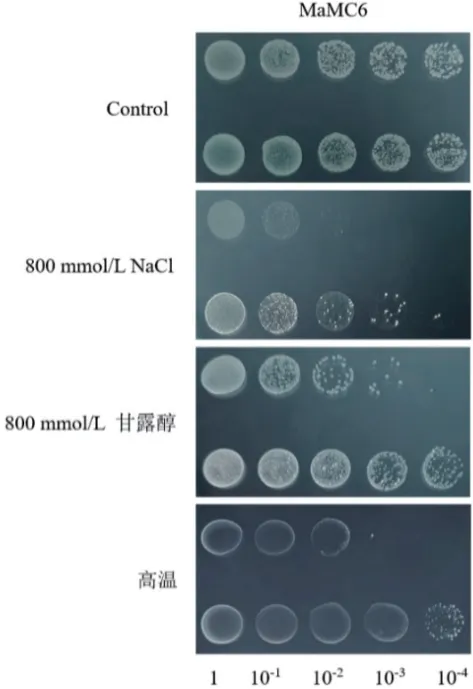
图9 MaMC6 在大肠杆菌BL21 中的功能验证Fig. 9 Functional verification of MaMC6 in Escherichia coli BL21
3 讨论
Metacaspases(MCs)属于植物类caspases 酶的半胱天冬氨酸酶类,通过激活下游具有类caspase 活性的蛋白酶,参与植物生长发育以及响应生物和非生物胁迫诱导的PCD 过程[11]。从香蕉A 基因组数据库鉴定到的MaMC6,利用生物学信息分析基因的特性。氨基酸序列多重比对以及系统进化树分析结果显示,MaMC6 属于II 型metacaspases,与唐露等[24]分析结果相似。跨膜结构预测显示MaMC6 没有跨膜结构,保守结构域分析发现只含有一个caspase‑like domain 与水稻和拟南芥一致[18]。顺式作用元件分析显示其含有防御和应激响应元件,猜测可能与香蕉的抗逆有关。
Metacaspase 在参与植物PCD 过程中的作用机制是复杂的,在拟南芥中AtMC1和AtMC2拮抗控制细胞死亡[12];过表达辣椒CaMC9,增强植株对病原体的敏感性和细胞的死亡率[32];水稻OsMCs基因经盐胁迫后的表达情况存在差异,过表达OsMC4基因有助于提高水稻对盐胁迫的耐受性[19,33]。香蕉根系响应盐胁迫的转录组学研究结果显示,外源Ca2+对盐胁迫诱导香蕉幼苗根系PCD 的调控可能与MaMCs有密切联系[34]。本实验用100 mmol/L NaCl对巴西蕉细胞进行人工模拟盐胁迫,使用RT‑qPCR分析MaMC6的表达情况。基因表达分析结果显示,MaMC6在2 h 时的上调表达倍数较高。与唐露等[24]在高浓度盐胁迫(200 mmol/L NaCl)诱导香蕉组培苗MaMCs表达中的结果存在一定差异,这可能和实验材料以及胁迫强度不同有关。另外,在烟草、豌豆、番茄、大豆等植物中均发现[Ca2+]cyt会影响PCD的发生[35-38]。为验证Ca2+是否会影响MaMC6的表达,本实验在盐胁迫基础上添加不同钙效应剂,然后测定MaMC6的相对表达量。结果发现,CaCl2会降低MaMC6的表达,而EGTA 和LaCl3导致MaMC6的表达升高,这可能与Ca2+能够缓解植物盐胁迫有关。
对MaMC6进行亚细胞定位实验,MaMC6呈现出细胞质和细胞核定位模式,这与拟南芥AtMC9(Atmetecaspase‑9)亚细胞定位结果相似[39],并且MaMC6和AtMC9在进化分析中有一定的同源性。为进一步鉴定MaMC6的功能,构建重组质粒pET28a‑MaMC6,将其转入大肠杆菌中分析该基因对胁迫的耐受性发现,重组菌株在NaCl、甘露醇和高温胁迫下的生长情况均优于对照,推测MaMC6可能是香蕉抗逆的候选基因。
目前, 关于香蕉MaMCs以及Ca2+与mete‑caspase 的研究较少。本实验从巴西蕉细胞中克隆到MaMC6,初步验证了该基因可受盐胁迫诱导表达并且其表达情况还受Ca2+影响,原核表达验证了MaMC6 对盐、干旱和高温胁迫的耐受性,为香蕉metecaspase 基因家族响应非生物胁迫的分子机制研究提供了参考。
4 结论
从巴西蕉细胞中克隆到MaMC6,该基因编码320 个氨基酸,属于稳定亲水蛋白,不含有跨膜结构,为II 型metacaspase,含有光、激素等响应元件。MaMC6受盐胁迫诱导表达,且在盐胁迫下的表达受Ca2+影响。MaMC6 呈现细胞质和细胞核定位。MaMC6的表达能提高大肠杆菌的耐盐、渗透和高温的能力。
猜你喜欢
数学物理学报(2021年4期)2021-08-30
新世纪智能(数学备考)(2020年10期)2021-01-04
家教世界·V家长(2019年3期)2019-06-11
食品科学(2018年10期)2018-05-23
中成药(2017年12期)2018-01-19
小天使·三年级语数英综合(2017年11期)2017-12-05
阅读与作文(小学高年级版)(2017年10期)2017-10-11
中国交通信息化(2017年8期)2017-06-06
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01