
许氏平鲉幼鱼对人工鱼礁模型不同阴影情况的行为响应❋
2024-01-11 07:03唐衍力林嘉政于浩林王欣欣
中国海洋大学学报(自然科学版) 2024年1期
唐衍力, 林嘉政, 王 欣, 于浩林, 王欣欣
(1. 中国海洋大学水产学院, 山东 青岛 266003; 2. 山东省渔业发展和资源养护总站, 山东 烟台 264003; 3. 中国科学院海洋研究所, 山东 青岛 266003)
人工鱼礁是海洋牧场建设中的基础生态工程,与天然礁石相比,其内部中空结构能够产生更大的阴影效应。研究表明鱼类能够使用视网膜和视网膜外光感受器来感知光照强度的变化[1-2],鱼类在光照强度较高的环境中的捕食效率和存活率较低,低光照强度的环境有助于提高幼鱼的存活率并减少种内捕食现象[3-4]。尽管鱼类对光照强度有一定的适应能力,但较弱的光照强度仍然是多数鱼种在幼鱼时期的适宜生活环境之一[5]。因此,许多学者认为,人工鱼礁诱集鱼类的重要因素之一是其阴影效应能够满足负趋光性的岩礁鱼类对于生存环境的需求,并利用室内行为学实验进行了验证[6-10]。目前,国内许多学者基于室内行为学实验方法探究了不同结构的人工鱼礁聚鱼效果,吴静等[11]探究了礁体的内外部结构对人工鱼礁诱集牙鲆(Paralichthysolivaceus)效果的影响,发现顶部无开孔内部无支架的立方体型模型礁诱集效果最好;唐衍力等[12]通过水槽实验研究了表面开孔对不同形状人工鱼礁诱集短蛸(Octopusocellatus)效果的影响,发现3种有孔模型礁诱集效果均优于无孔模型礁;周艳波等[13-15]通过室内模拟实验研究了多种实际投放的人工鱼礁对几种岩礁性鱼类的诱集效果,发现有效空间大且遮蔽效果好的人工鱼礁模型对花尾胡椒鲷(Plectorhinchuscinctus)和褐菖鲉(Sebastiscusmarmoratus)的诱集效果最好,而黑棘鲷(Sparusmacrocephlus)则偏好于框架结构礁体;李磊等[16-17]研究了3种不同外形和内部构造的人工鱼礁对黑棘鲷、中国花鲈(Lateolabraxmaculatus)和大黄鱼(Larimichthyscrocea)的诱集效果,认为鱼礁的有效内部空间和遮蔽空间都会对诱集效果产生影响。总体上,对人工鱼礁诱鱼效果的室内实验主要集中在计划或已投放礁型的效果验证和比较,尚无针对人工鱼礁阴影效应的专门研究报道。
许氏平鲉(Sebastesschlegelii)俗称黑鲪,隶属鲉形目(Scorpaeniformes)鲉科(Scorpaenidae)平鲉属(Sebastes),为冷温性近海底层卵胎生鱼类,是中国黄渤海区重要的岩礁性经济鱼种,具有适应力强、生长快、营养价值高等特点[18]。近年来由于捕捞强度过大,环境污染加剧,许氏平鲉资源量急剧减少[19]。因此,我国开展了较大规模的许氏平鲉增殖放流行动[20]。Brennan等[21]在增殖放流过程中幼鱼存活率的研究中指出,放流后幼鱼的存活率受到捕食者捕食的影响较大,这造成幼鱼较高的死亡率。因此在对现有的放流技术进行改进的同时[18],对人工鱼礁结构进行优化设计,可以增强其对放流苗种的诱集保护效果,从而降低幼鱼的流散率和死亡率。本研究通过室内水槽实验,探究了许氏平鲉幼鱼对人工鱼礁模型不同阴影情况下的行为响应,以期为今后人工鱼礁的设计和建设提供参考。
1 材料与方法
1.1 实验生物
实验用鱼为烟台开发区天源水产有限公司培育用于增殖放流的许氏平鲉幼鱼(体长(5.06±0.09) cm,体质量(3.22±0.16) g),所选个体体观正常、生长良好。将200尾实验鱼置于养殖水槽内暂养1周,暂养用水为砂滤海水,水温为(17.36±0.65) ℃,盐度为31.04±0.04,pH为7.88±0.07,溶氧保持在7 mg/L以上。暂养期间正常投喂饲料和充氧,每次实验前随机取30尾实验鱼进行实验。
1.2 实验设备与布设
实验于2021年10—11月在公司的实验车间进行,实验环境封闭,无外部光照。实验水槽为圆桶形,高86 cm,直径182 cm。水槽底部划分为5个同心圆,同心圆之间的外径间距为14 cm,半径由小到大分别为14、28、42、56、70 cm。整个实验水槽可分为Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ共6个区域(见图1),区域Ⅰ阴影部分为鱼礁模型放置区域。水槽正上方安装有1个LED灯作为实验光源,同时在LED灯旁安装普联牌摄像机1台(型号为TL-IPC44AN),用于实验期间全程摄像。实验海水与暂养海水水质相同,每次实验结束后清理粪便,每天换水1次。为避免饲料及充气石形成的局部水流对鱼类分布造成影响,实验期间不投喂饲料、不充氧,同时用黑布将水槽四周至光源区域包围,尽可能减少外部环境的干扰。
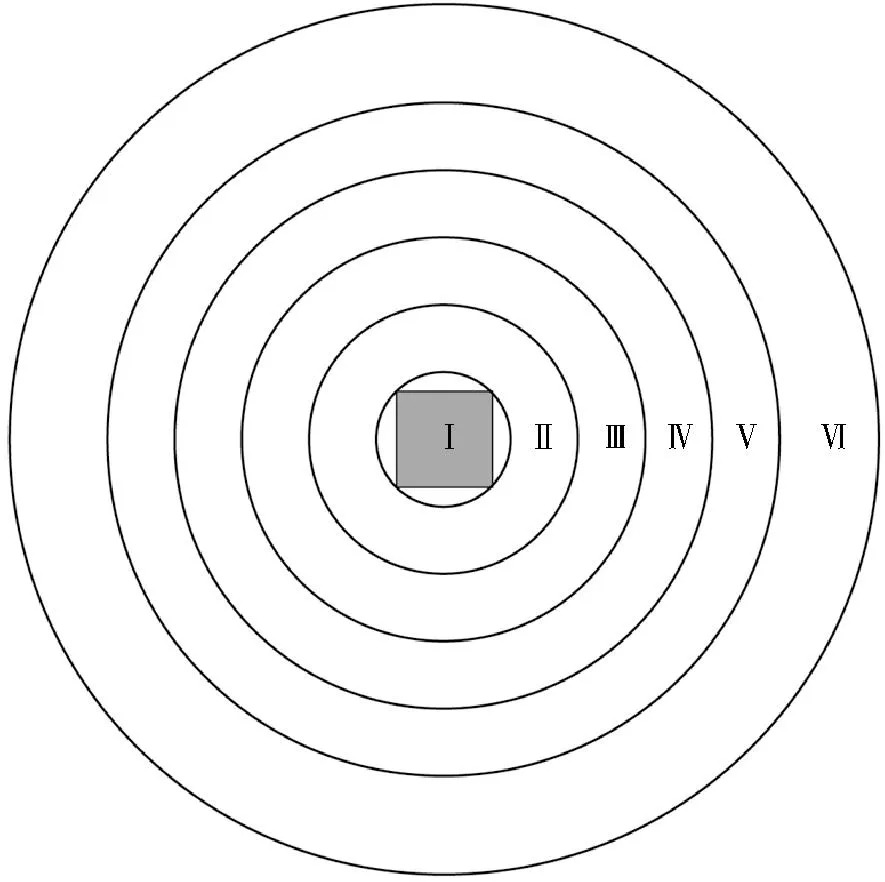
(阴影区域为鱼礁模型放置区。The reef model placement area was shown in shade.)图1 区域划分示意图Fig.1 Schematic diagram of area division
1.3 实验设计
本实验在两个条件相同的实验水槽中进行,鱼礁模型放置在实验水槽中央的区域Ⅰ(见图1)。实验开始前,分别向两个实验水槽注入35 cm深的海水,从暂养水槽中随机选取30尾鱼放入水槽,使实验鱼在水槽中适应20 min,然后开始实验,单次实验时间3 h,每组重复3次。实验中用摄像机连续记录水槽中许氏平鲉的分布情况。每次试验结束后,更换一批新的实验鱼。
为模拟人工鱼礁的典型阴影特征,本实验基于国内常用的立方体型人工鱼礁[22-23]设计制作了一种材质为ABS塑料、规格为20 cm×20 cm×20 cm的正方体人工鱼礁模型(见图2(a)),鱼礁模型四周有开孔率为25%的圆孔,顶板可更换以便形成不同阴影(见图2(b))。鱼礁模型投放前均放置于水桶中用海水浸泡3 d。A组实验目的是探究不同阴影面积的鱼礁对实验鱼的诱集效果。实验工况A1—A4组的顶板开孔率分别为0%、12.5%、25%、50%;对照工况A5组不放置顶板、开孔率为100%(见表1)。B组实验目的是探究不放置鱼礁模型、悬挂1个开孔顶板时不同阴影面积对实验鱼的诱集效果,B组的开孔率、实验阴影面积和形状同A组。实验工况B1—B4组的顶板开孔率分别为0%、12.5%、25%、50%;对照工况B5组不放置鱼礁模型、不悬挂顶板。C组实验目的是探究阴影面积一样但开孔数量不同的鱼礁对实验鱼的诱集效果。实验工况C1—C4组的顶板开孔率均为25%、开孔数量分别为1、4、6、9个;对照组与B5组的实验工况相同,实验水槽中不放置鱼礁模型、不悬挂顶板。
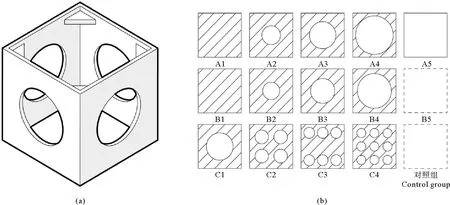
((a)鱼礁框架模型示意图;(b)鱼礁顶板示意图。(a) Schematic diagram of reef frame model; (b) Schematic diagram of the top reef panel.)图2 鱼礁模型示意图Fig.2 Schematic diagram of reef module
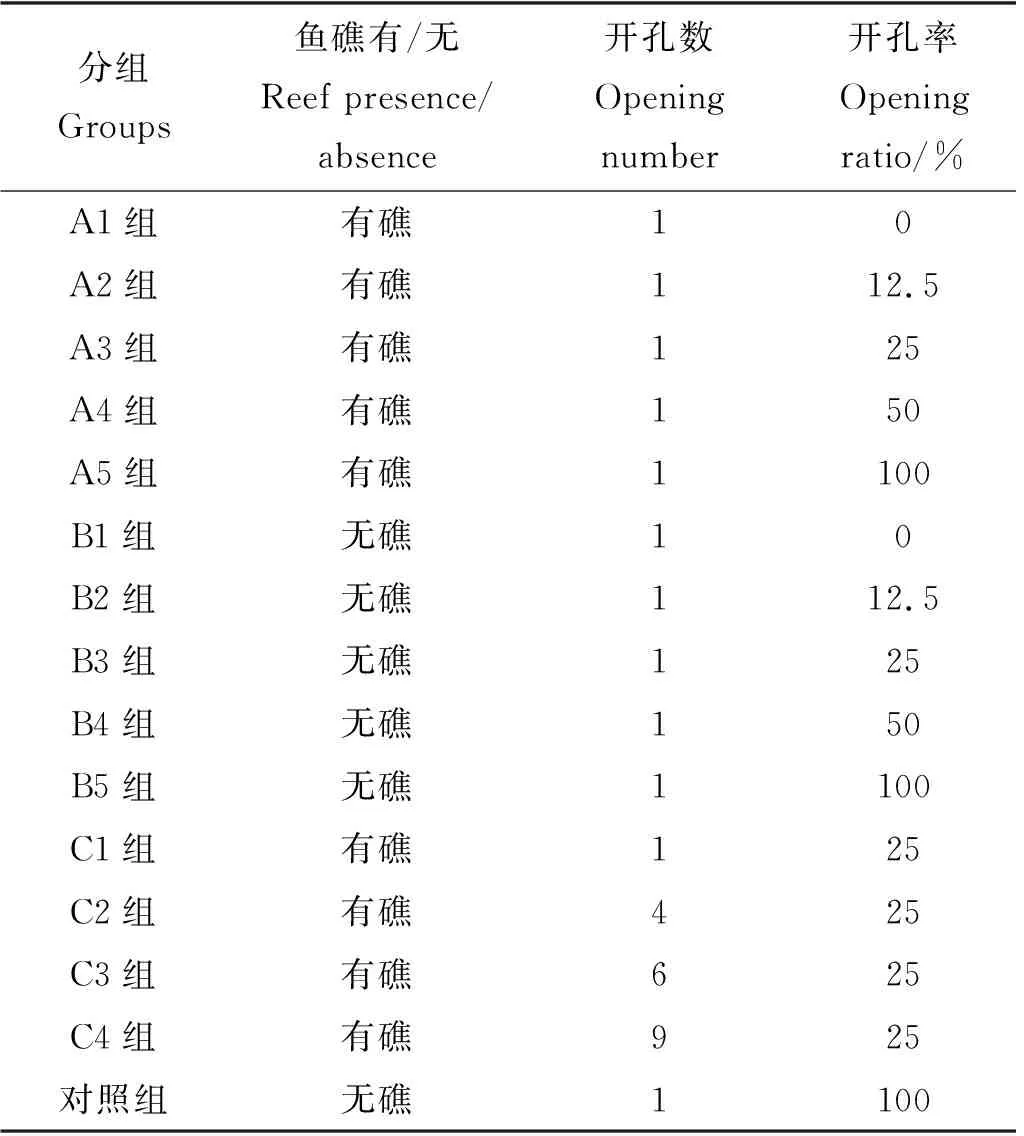
表1 实验设计Table 1 Experimental design
1.4 分析方法
使用FFmpeg软件将实验录像每隔2 min生成1张截图,每次实验总计生成90张截图。使用Engauge Digitizer软件标注每张截图中实验鱼的位置坐标,统计实验鱼与水槽中心的距离数据。通过IBM SPSS statistics 26.0软件进行各实验组内不同水平的单因素方差分析(One-Way ANOVA),使用Tamhane’s T2法进行事后检验。构建广义线性模型,将实验鱼在各区域的平均出现率作为因变量,各实验组(鱼礁有无、开孔数、开孔率)作为自变量,探究各实验组与出现率的相关关系。根据Linderman、Merenda、Gold[24]共同提出的计算自变量相对重要性的方法(LMG方法),通过R软件中的“relaimpo”包计算各实验组变量对平均出现率的相对贡献度[25]。LMG方法能够在考虑自变量本身和其他自变量共同影响的条件下,将多元线性回归模型的总体R2分解为每个自变量的相对贡献值[26]。
平均出现率(Average occurrence rate,RAO)指实验鱼在某一区域出现的总频数占实验鱼在各区域出现的总频数的百分比:
式中:mi表示在第i次观测时实验鱼在某一区域中出现的频数;Mi是在第i次观测时实验鱼出现在各区域的总频数。
实验中由于圆环之间的面积存在等差数列特征,面积比为1∶3∶5∶7∶9,在进行平均出现率的比较时需配平,即
式中n为区域标号,值为1、2、3、4、5。其中Ⅵ区域平均出现率配平为
构建实验鱼平均出现率与各实验组的广义线性模型
Y=α+α1x1+α2x2+…+αnxn+ε。
式中:Y为实验鱼平均出现率的对数;α为函数的截距;αi为斜率;xi为自变量;ε为误差项,与自变量无关,服从N(0,σ2)。
基于截图中实验鱼的位置坐标信息,使用ArcGIS 10.8软件的核密度分析工具绘制各实验工况下实验鱼核密度分布图。将整个实验区域除鱼礁放置区Ⅰ外,以1 cm为间隔划分网格,统计实验鱼出现频数,拟合光滑核密度函数,核密度值越高,表示出现频数越高,反之亦然[27]。
2 结果
2.1 行为描述
通过观察不同组中许氏平鲉的行为特征发现,对照组中许氏平鲉在进入实验水槽后,先向水槽四周游动,在遍历整个水槽后有些个体沿水槽壁自由游动,有些个体在水槽中心处停留。实验组中,许氏平鲉在进入实验水槽后,多数个体会游向礁体放置区域并紧贴鱼礁边缘或游动或休憩或进入礁体内部停留,少部分许氏平鲉在靠近水槽边缘的区域游动。随实验时间的延长,部分个体从礁体内部游出,同时有相同数量的许氏平鲉进入礁体,礁体内部个体数量基本保持恒定,且主要分布在明暗交替处。直到实验结束前,水槽边缘始终分布着一定数量的许氏平鲉。
2.2 对照组实验鱼在各区域的分布
对照组中,许氏平鲉在各区域平均出现率高低依次为Ⅰ>Ⅱ>Ⅲ>Ⅵ>Ⅳ>Ⅴ,各区域中不同方向上许氏平鲉出现频数差别较小(见图3和4),其中,区域Ⅰ平均出现率显著高于区域Ⅲ、Ⅳ、Ⅴ、Ⅵ (P<0.05),与区域Ⅱ之间差异不显著(P>0.05);区域Ⅱ与区域Ⅵ之间存在显著差异(P>0.05),与区域Ⅲ、Ⅳ、Ⅴ 之间差异不显著(P>0.05);区域Ⅲ、Ⅳ、Ⅴ、Ⅵ之间差异不显著(P>0.05)。
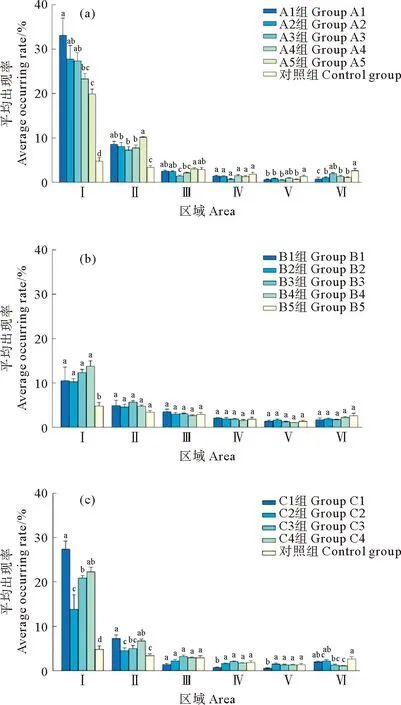
((a)A组实验;(b)B组实验;(c)C组实验。同一区域内字母不同表示各组间平均出现率差异显著(P<0.05)。(a)Group A experiment;(b)Group B experiment; (c)Group C experiment. Different letters indicate significant differences in average occurrence rate among different experimental groups for the same area(P<0.05).)图3 不同实验组的各区域平均出现率Fig.3 Average occurrence rate of each area among different experimental groups
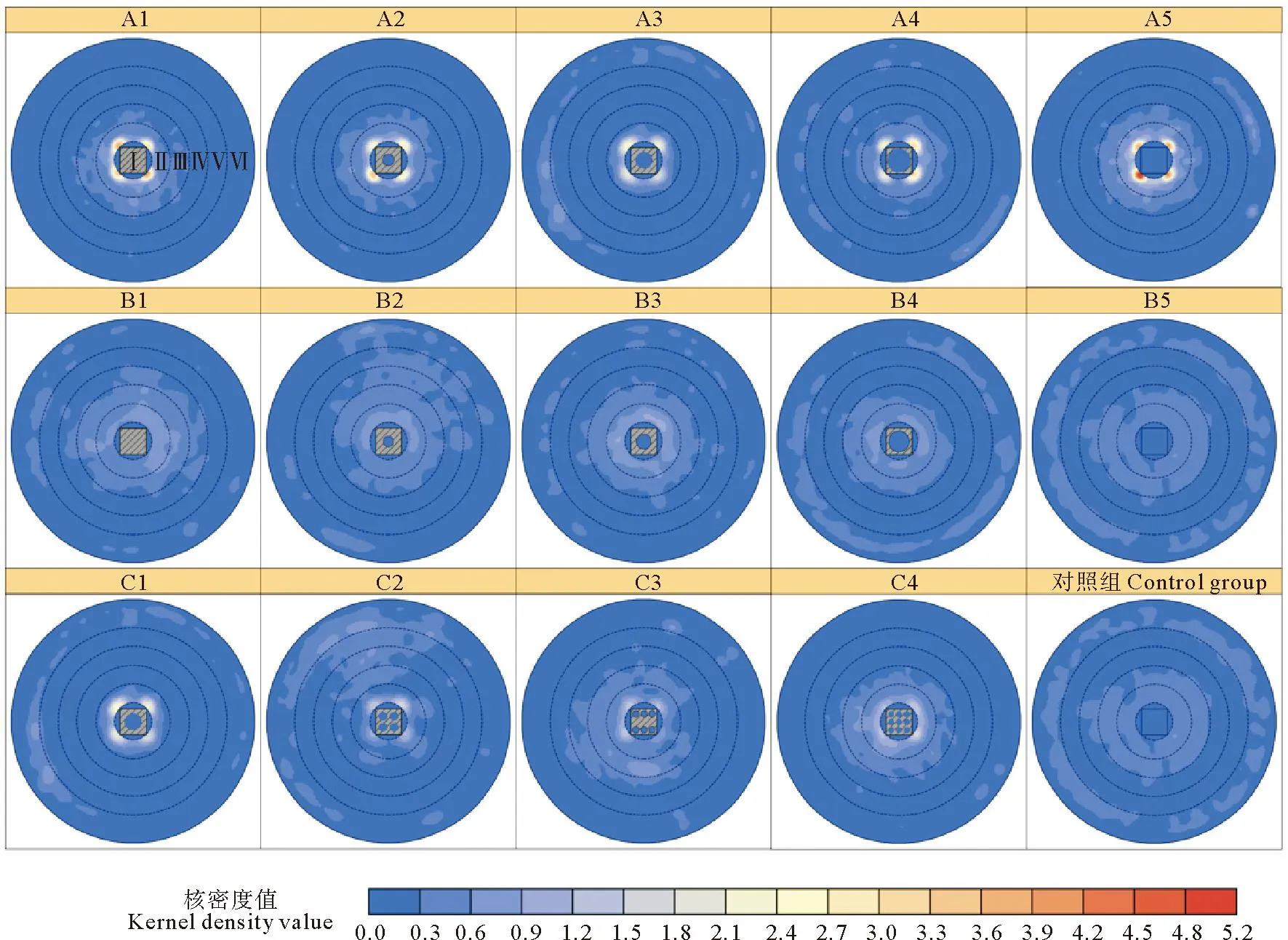
图4 不同实验组的实验鱼出现频数核密度分析Fig.4 Kernel density analysis of experimental fish occurrence frequency among different experimental groups
2.3 鱼礁模型顶板不同开孔率诱集效果
A组实验中,许氏平鲉在区域Ⅰ、Ⅱ的平均出现率显著高于对照组(见图3(a),P<0.05),区域Ⅲ、Ⅳ、Ⅴ平均出现率与对照组相近,靠近水槽壁的区域Ⅵ平均出现率低于对照组。区域Ⅰ中许氏平鲉平均出现率存在显著差异(P<0.05),随着开孔率增大,平均出现率逐渐下降;区域Ⅱ中平均出现率与区域Ⅰ相比有所下降,随着开孔率的增大呈现先下降后上升的趋势,组间差异显著(P<0.05),许氏平鲉在该区域多聚集于鱼礁模型的折角处(见图4);区域Ⅲ、Ⅳ、Ⅴ、Ⅵ组间差异不显著(P>0.05),区域Ⅲ、Ⅳ平均出现率随开孔率增大呈现先下降后上升的趋势,区域Ⅵ平均出现率随开孔率增大呈现先上升后下降的趋势,各区域不同方向上许氏平鲉分布较为分散。
2.4 悬挂不同开孔率顶板的诱集效果
B组实验中,许氏平鲉在区域Ⅰ、区域Ⅱ的平均出现率相较于A组实验有明显的下降,在各区域不同方向上分布较为分散(见图3(b)和图4)。区域Ⅰ中B1—B4组的平均出现率均显著高于B5组(P<0.05),B1—B4组之间差异不显著(P>0.05);区域Ⅱ、Ⅲ、Ⅳ、Ⅴ中各组平均出现率相较于区域Ⅰ有所下降,单个区域中各组之间平均出现率差异不显著(P>0.05);区域Ⅵ各组平均出现率相较于区域Ⅴ有所上升且B5组平均出现率高于其余4组,但各组间差异不显著(P>0.05)。
2.5 鱼礁模型顶板不同开孔数诱集效果
C组实验中,区域Ⅰ中4个实验组之间平均出现率有显著差异(见图3(c),P<0.05),且都高于对照组;区域Ⅱ各组平均出现率较区域Ⅰ有所降低,C1组、C4组显著高于对照组(P<0.05),C2组、C3组与对照组差异不显著(P>0.05),许氏平鲉在该区域多聚集于鱼礁模型的折角处(见图4),但出现频数总体低于A组;区域Ⅲ、Ⅳ、Ⅴ中,C1组平均出现率较低,其余实验组平均出现率与对照组相近(P>0.05);区域Ⅵ各实验组平均出现率小于对照组,组间差异显著(P<0.05),区域Ⅲ、Ⅳ、Ⅴ、Ⅵ不同方向上许氏平鲉分布较为均匀。
2.6 各实验变量与平均出现率的相关关系及相对贡献度
广义线性模型分析结果显示开孔率只对区域Ⅰ平均出现率有极显著影响(P<0.01)且与其呈负相关关系(见表2),对其他区域的平均出现率影响不显著(P>0.05);开孔数对区域Ⅰ、Ⅱ、Ⅳ、Ⅴ的平均出现率有极显著影响(P<0.01)且与区域Ⅰ、Ⅱ的平均出现率呈负相关关系,对区域Ⅲ的平均出现率有显著影响(P<0.05),对区域Ⅵ的平均出现率影响不显著(P>0.05);是否放置鱼礁对各区域的平均出现率均有极显著影响(P<0.01),与区域Ⅲ、Ⅳ、Ⅴ、Ⅵ的平均出现率呈负相关关系。
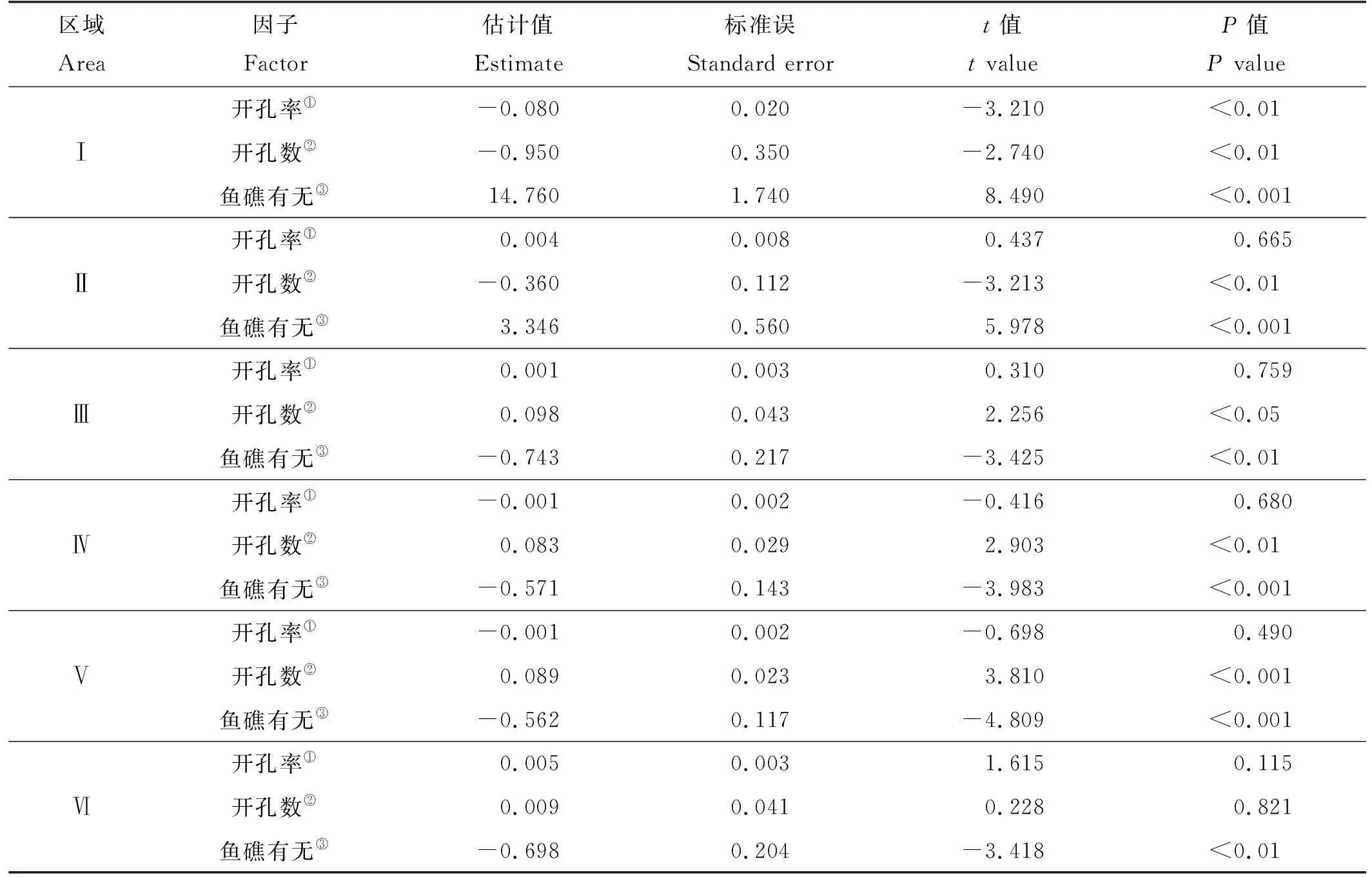
表2 各实验组的各区域实验变量对平均出现率影响的广义线性模型显著性的检验(α=0.05)Table 2 Statistics of GLM relationships between experimental variable and average occurrence rate
本文使用LMG方法将3个实验变量(鱼礁有无、开孔数、开孔率)对实验鱼在不同区域的平均出现率的影响进行量化。在6个区域中,在平均出现率的相对贡献度中是否放置鱼礁这一变量大于其他2个变量之和(见图5)。开孔率对区域Ⅰ和区域Ⅵ平均出现率的相对贡献度大于开孔数,而对区域Ⅱ、Ⅲ、Ⅳ、Ⅴ的相对贡献度小于开孔数。开孔率对区域Ⅱ、Ⅲ、Ⅳ、Ⅴ平均出现率的相对贡献度均在5%以下。
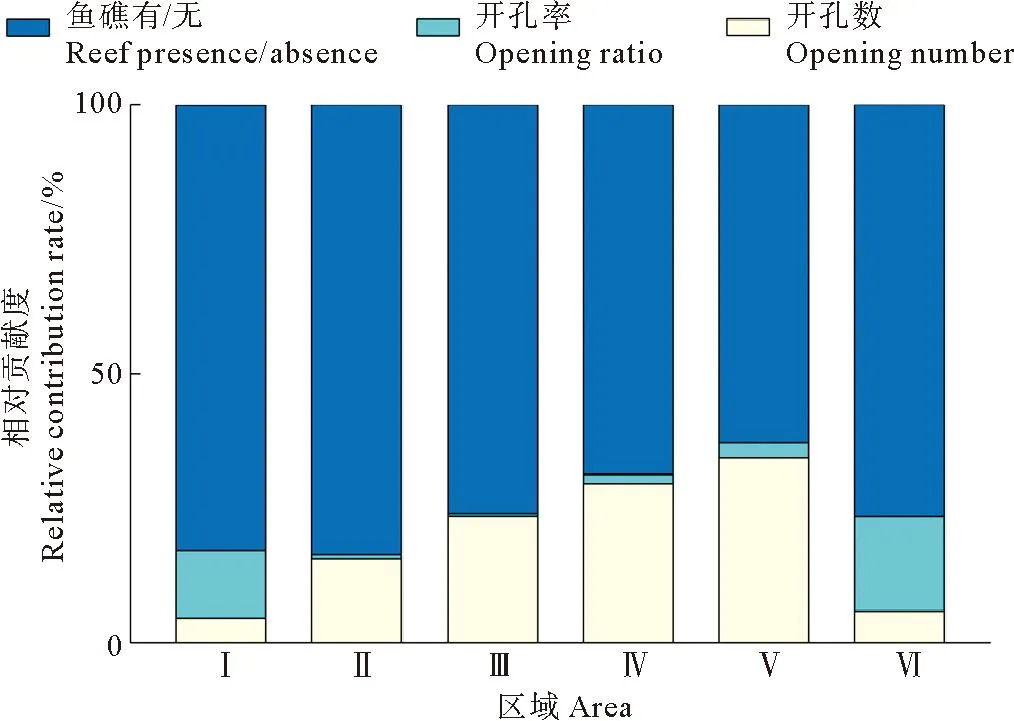
图5 实验变量对平均出现率的相对贡献度Fig.5 Relative contribution rate of experimental variables to average occurrence rate
3 讨论
A组实验结果表明,人工鱼礁的内部阴影面积与许氏平鲉的平均出现率呈显著正相关关系。此结果与其他学者对许氏平鲉的研究结果一致[28-29],这可能与许氏平鲉的生活习性有关,避敌、索饵和休憩是许氏平鲉在幼鱼期的主要活动方式[30],对复杂底质类型及阴影区域偏好较强。顶板开孔率增大,鱼礁模型内部的阴影减少,可供幼鱼躲藏的空间也随之减少,这是因为岩礁性鱼类需要占据一定的栖息空间,且具备明显的领域行为[31]。由于许氏平鲉的趋礁性,即使鱼礁内个体数量接近饱和,部分许氏平鲉依然在靠近人工鱼礁模型的区域Ⅱ游动并尝试再次进入区域Ⅰ。田方等[32]通过实验发现阴影面积较大的鱼礁模型对真鲷的诱集效果较好,这与本研究结果相同。
B组实验排除了水中是否放置人工鱼礁模型这一变量。不同的阴影面积对许氏平鲉幼鱼的诱集效果无明显规律。比较A、B组实验结果,B组实验许氏平鲉在区域Ⅰ的平均出现率明显低于A组实验。水面上悬挂的鱼礁顶板能够产生与放置鱼礁时形状和面积相同的阴影,许氏平鲉由于负趋光性倾向于在顶板产生的阴影下游泳。但鱼礁顶板表面较光滑,导致灯光经由水面-顶板-水面被反射到阴影区域形成闪烁的光斑,对鱼类有一定的驱散作用[33]。有研究表明,许氏平鲉属于趋触性鱼类,具有强烈的与岩礁接触的习性[6],B组实验结果与此结论相似,由于鱼礁模型的移除,水体缺少人工鱼礁空间结构的遮蔽效果,使得B组实验许氏平鲉在区域Ⅰ的出现率较A组明显下降。
C组实验探究了鱼礁模型顶板不同开孔数的诱集效果。顶部开孔数为1时,人工鱼礁对许氏平鲉幼鱼的诱集效果最好。这可能与岩礁性鱼类对不同形状阴影的偏好有关。当开孔数为1时,鱼礁内部的边际区域可形成大面积的连续阴影,有利于体型相对较小的幼鱼在阴影区域聚集。Li等[34]在静水条件下观察了不同人工鱼礁模型对许氏平鲉的诱集效果,发现具有连续阴影的立方体型鱼礁诱集效果好于星型鱼礁、金字塔型鱼礁和圆管型鱼礁,这与本实验结果相符。而C组其余实验当中,鱼礁内部的阴影被分割成多个部分,幼鱼在阴影中稍有游动就会受到灯光刺激,因此对幼鱼的诱集效果弱于开孔数为1时。鱼礁顶板开孔数目为9时,许氏平鲉在区域Ⅰ的平均出现率与开孔数为1时差异不显著,这可能是因为顶板开孔孔径与许氏平鲉幼鱼的体高相近,且顶板的复杂度高更易于吸引趋礁性鱼类的穿梭与游泳。这与Zhang等[35]发现复杂底质类型更有利于吸引许氏平鲉的实验结果一致。
核密度分析图显示,在水槽中未放置鱼礁模型的工况中,许氏平鲉在各方向上的分布较为均匀,而在放置鱼礁模型的实验工况中更倾向于出现在鱼礁模型的四个折角处。这说明礁体模型的存在能改变许氏平鲉的空间分布,杨吝等[6]认为许氏平鲉属于趋礁性鱼类,其具有强烈与岩礁接触的习性,鱼礁模型的折角容易与鱼鳍等部位发生接触,因此许氏平鲉倾向于在鱼礁模型折角附近活动。Lee等[36]在对同属岩礁性鱼类的黑棘鲷、黄鳍棘鲷(Acanthopagruslatus)的水槽实验研究中也发现,放置鱼礁模型能够明显地改变实验鱼在水槽中的空间分布,且两种实验鱼均倾向于在鱼礁模型折角处出现,这与本文所研究的许氏平鲉相似。
广义线性模型分析表明,不同实验变量对许氏平鲉分布的影响范围不同。其中,开孔率的影响范围最小,是因为改变顶板的开孔率只影响了阴影面积的大小,由于鱼类的领域行为,未能在阴影下躲避的许氏平鲉幼鱼会选择在实验水槽中游动或将阴影中的幼鱼驱逐;而改变开孔数则是在阴影面积不变的前提下改变了阴影的形状,受开孔数改变影响的区域数多于受开孔率影响的区域数,这可能是因为不同的开孔数改变了鱼礁内部的阴影环境,许氏平鲉幼鱼对阴影形状的敏感度高于对阴影面积的敏感度,这可能与幼鱼躲避大型捕食者的天性有关;是否放置鱼礁对实验水槽各区域许氏平鲉幼鱼的分布有极显著影响(P<0.01),人工鱼礁模型一方面能满足岩礁性鱼类趋触性的本能,另一方面鱼礁内相对封闭的空间对许氏平鲉产生了诱集效应,一些学者对其他岩礁性鱼类也进行了相似的室内水槽实验,结果表明模型礁的存在对实验鱼具有较强的诱集效果[15-17,37],与本研究结果一致。LMG方法分析结果与广义线性模型结果一致。由于鱼类趋触性的影响,人工鱼礁的阴影效应需要依附于鱼礁实体才能发挥作用;拥有更大阴影面积的人工鱼礁的诱集效果要好;在鱼礁总阴影面积不变的情况下,在表面制作尽量多的开孔,既能够增强对幼鱼的吸引与保护作用,也能拥有与单个开孔鱼礁相近的诱集效果。
在鱼礁表面开孔,是礁体结构设计中的常规选择,但对于开孔数量和大小的选择的参考依据较少[38]。本研究探讨了许氏平鲉对立方体型人工鱼礁不同阴影情况的行为响应,可为人工鱼礁表面开孔设计提供参考。
猜你喜欢
海洋通报(2022年5期)2022-11-30
水产养殖(2021年5期)2021-05-21
海洋通报(2020年5期)2021-01-14
时代农机(2019年8期)2019-12-27
珠江水运(2018年21期)2018-12-20
读者·校园版(2018年22期)2018-11-01
大自然探索(2018年7期)2018-09-01
保健与生活(2018年17期)2018-01-27
中国海洋大学学报(自然科学版)(2015年8期)2015-03-18
食品工业科技(2014年15期)2014-03-11
