
甘蔗转录因子DREB2 基因的克隆和生物信息学分析*
2024-01-11 09:15贾晨灏夏可灿钱庆忠黎慧敏苗贵东
科技创新与生产力 2023年12期
贾晨灏,夏可灿,2,钱庆忠,李 浩,黎慧敏,苗贵东,2
(1.兴义民族师范学院生物与化学学院,贵州 兴义 562400,2.黔西南州生物遗传资源挖掘与分子育种重点实验室,贵州 兴义 56240)
我国甘蔗区种植面积70%以上为旱坡地,甘蔗区条件逐渐由水田向高海拔、低降雨量、土壤贫瘠的旱坡地转移。干旱已成为我国甘蔗单产低、品质差的主要原因之一。如何解决干旱胁迫下甘蔗的产量问题是目前生产上面临的重要课题。甘蔗作为一种高度杂合、遗传背景复杂的无性繁殖作物,常规的抗旱选育方法盲目性大,周期长,效果也有限。随着分子辅助育种的快速发展,从分子水平改良品种抗旱性成为了可能,可以在很大程度上克服常规抗旱育种中的一些问题,从而提高育种效率,加速抗旱甘蔗品种的培育和推广。
干旱应答元件结合蛋白(Dehydration Responsive Element Binding Protein,DREB) 转录因子属于AP2/EREBP 转录因子家族,具有一个典型的AP2/EREBP 保守结构域[1],由60-70 个氨基酸组成,能够特异性结合DRE 顺式作用元件,调控与干旱、高盐、高渗、极端温度等非生物胁迫应答有关的基因表达,从而增强植物对多种胁迫的抵抗性。DREB 亚族家族具有6 个组,即A1~A6[2]。
自1997年首次从拟南芥基因中分离到编码DREB 蛋白质的CBF1 基因[3]以来,现已从拟南芥、水稻等植物中分离出DREB 类转录因子[4]。2007年,福建农林大学农业部甘蔗生理生态与遗传改良重点实验室克隆到甘蔗近缘属斑茅的DREB 基因同系物EaDREB2B,通过拟南芥诱导型启动子rd29A 驱动其在甘蔗中表达,获得了抗旱的转EaDREB2B 基因甘蔗无性系,检测结果表明该基因在转基因甘蔗中能稳定表达,并提高甘蔗的抗旱能力[5]。同时,中国热带农业科学院湛江实验站刘洋博士团队以割手密为材料,采用同源基因克隆的方法获得2 个DREB2类转录因子SsDREB2-a 和SsDREB2-f(Gen Bank登录号分别为KU963272 和KU963277)[6]。鉴于DREB 基因在植物非生物胁迫抗性过程中起着重要作用,本研究利用同源基因克隆技术从甘蔗品种中获得1 个ScDREB2 基因,比较分析获得的序列,以期为后续该基因在甘蔗抗逆育种上的开发利用和分子调控机制研究奠定理论基础和提供指导。
1 材料与方法
1.1 实验材料
以F113 甘蔗品种为材料,由国家甘蔗种质资源圃提供。
1.2 甘蔗叶片总DNA 提取
叶片总DNA 的提取采用天根植物基因组DNA提取试剂盒(Plant Genomic DNA kit)。
1.3 ScDREB2 基因克隆
参考姚艳丽等[6]在割手密上设计的两对特异引物,以DNA 为模板,采用巢式PCR 进行扩增。PCR 经1%琼脂糖凝胶电泳检测,采用Easy Pure Quick Gel Extraction Kit 回收,使用pEASY- Blunt Zero Cloning Kit 将回收的扩增产物连接、转化到Trans1-T1 感受态细胞中,阳性克隆经菌落PCR 鉴定后,再转入含氨苄青霉素的培养基中37 ℃培养12 h,至少挑取40 份克隆,送生工生物工程(上海) 股份有限公司测序。
1.4 生物信息学分析
使用ExPASy 在线平台的ProtParam tool 在线预测甘蔗DREB2 蛋白质的等电点和相对分子质量,ProtScale 进行疏水性/亲水性预测,SignaIP 5.0 软件预测信号肽、NetNGlyc-1.0 及YinOYang-1.2 进行N-糖基化位点预测和O-糖基化位点预测、NetPhos3.1 进行磷酸化位点预测,运用SOPMA 在线服务器进行蛋白二级结构分析,SWISS-MODEL进行蛋白三级结构分析,采用SMART 在线预测网站进行结构域与功能区分析。使用DNAstar 软件包中的MagAlign 程序进行同源性比对,使用MEGA 11.0 软件构建系统进化树。
2 结果与分析
2.1 ScDREB2 基因的克隆及序列分析
ScDREB2 基因PCR 扩增琼脂糖凝胶电泳检测见图1。从甘蔗中扩增得到的ScDREB2 基因,序列长度为1537 bp,通过NCBI 网站的ORF Finder 程序进行分析,测得其中包括789 bp 的开放阅读框(ORF),整个ORF 编码262 个氨基酸。
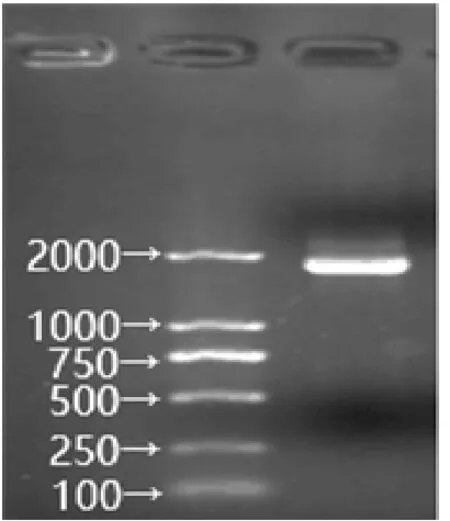
图1 ScDREB2 基因PCR 扩增琼脂糖凝胶电泳检测
利用BLAST 程序发现所得目的基因同源序列与参考序列(KU963272 和KU963277) 覆盖率为100%,相似性达97.73% (KU963272) 和95.93%(KU963277),经序列比对分析发现,该序列含有2 个外显子和1 个内含子。
2.2 甘蔗ScDREB2 基因氨基酸序列同源进化分析
采用MEGA 11.0 软件,将甘蔗DREB2 转录因子氨基酸序列与甜根子草、高粱等11 种植物的DREB2 蛋白氨基酸序列进行同源遗传进化分析,见图2。由图2 可知,甘蔗与甜根子草被聚在同一亚类,因此,它们之间具有较高的同源性和较近的亲缘关系。此进化树与传统形态学分类基本一致。进化分析表明,ScDREB2 基因编码蛋白为DREB2蛋白为AP2/EREBP 类DREB 亚族家族A-2 组转录因子。图3 为甘蔗DREB2 蛋白氨基酸序列。
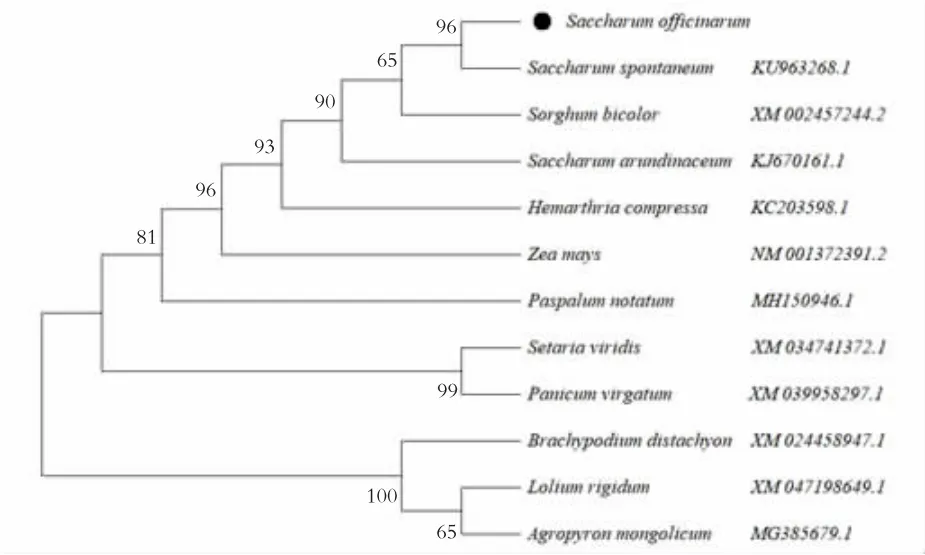
图2 甘蔗DREB2 蛋白氨基酸序列的同源遗传进化分析

图3 甘蔗DREB2 蛋白氨基酸序列
2.3 甘蔗ScDREB2 转录因子的生物信息学分析
2.3.1 甘蔗ScDREB2 转录因子的等电点和分子量预测
通过ExPASy 在线平台的ProtParam tool 在线预测甘蔗ScDREB2 蛋白质的等电点和相对分子质量,见图4。结果显示,DREB 转录因子等电点为9.25和相对分子质量为26861.97。该蛋白质由3972 个原子组成,分子式为C1238H1958N360O406S10。其不稳定指数(Ⅱ)计算为43.95,为不稳定蛋白质。
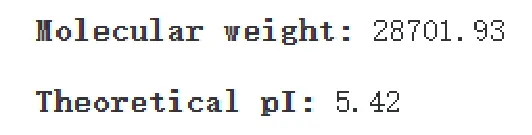
图4 甘蔗DREB2 蛋白质等电点和相对分子质量预测
2.3.2 甘蔗ScDREB2 转录因子亲水性/疏水性分析
通过ExPASy 在线平台上的ProtScale 对甘蔗ScDREB2 转录因子进行疏水性/亲水性预测,见图5。分析图5 疏水性图谱可知,多肽链的第234 位的异亮氨酸基(Ile) 具有最高的分值,为2.289,疏水性最强。第27 位的甲硫氨酸(Met),具有最低的分值,为-3.178;其次是第46 位的谷氨酸(Glu),为-3.056;以及第251 位的组氨酸(His),为-2.844。三者均为亲水氨基酸。经过计算可知,甘蔗ScDREB2 转录因子呈亲水性(Hydropathicity),为亲水性蛋白。
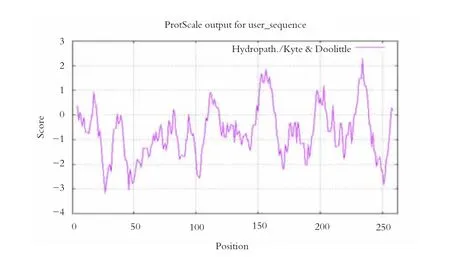
图5 甘蔗ScDREB2 转录因子的亲水性/疏水性预测
2.3.3 甘蔗ScDREB2 转录因子活性位点预测分析
利用在线服务器SignaIP 5.0 预测信号肽、NetNGlyc-1.0 和NetOGlyc-4.0 预测N-糖基化位点和O-糖基化位点、NetPhos3.1 预测磷酸化位点,结果见图6。拥有信号肽的概率为0.0022,可知DREB2 类转录因子不存在信号肽,为非分泌蛋白。甘蔗DREB2 类转录因子含有32 个O-糖基化位点,1 个N-糖基化位点,由于DREB2 蛋白质不存在信号肽,虽然预测图上出现可能存在的1 个N-糖基化位点,但是因为没有信号肽的蛋白质不太可能暴露于N-糖基化机制下,因此DREB2 蛋白质应不存在糖基化位点。
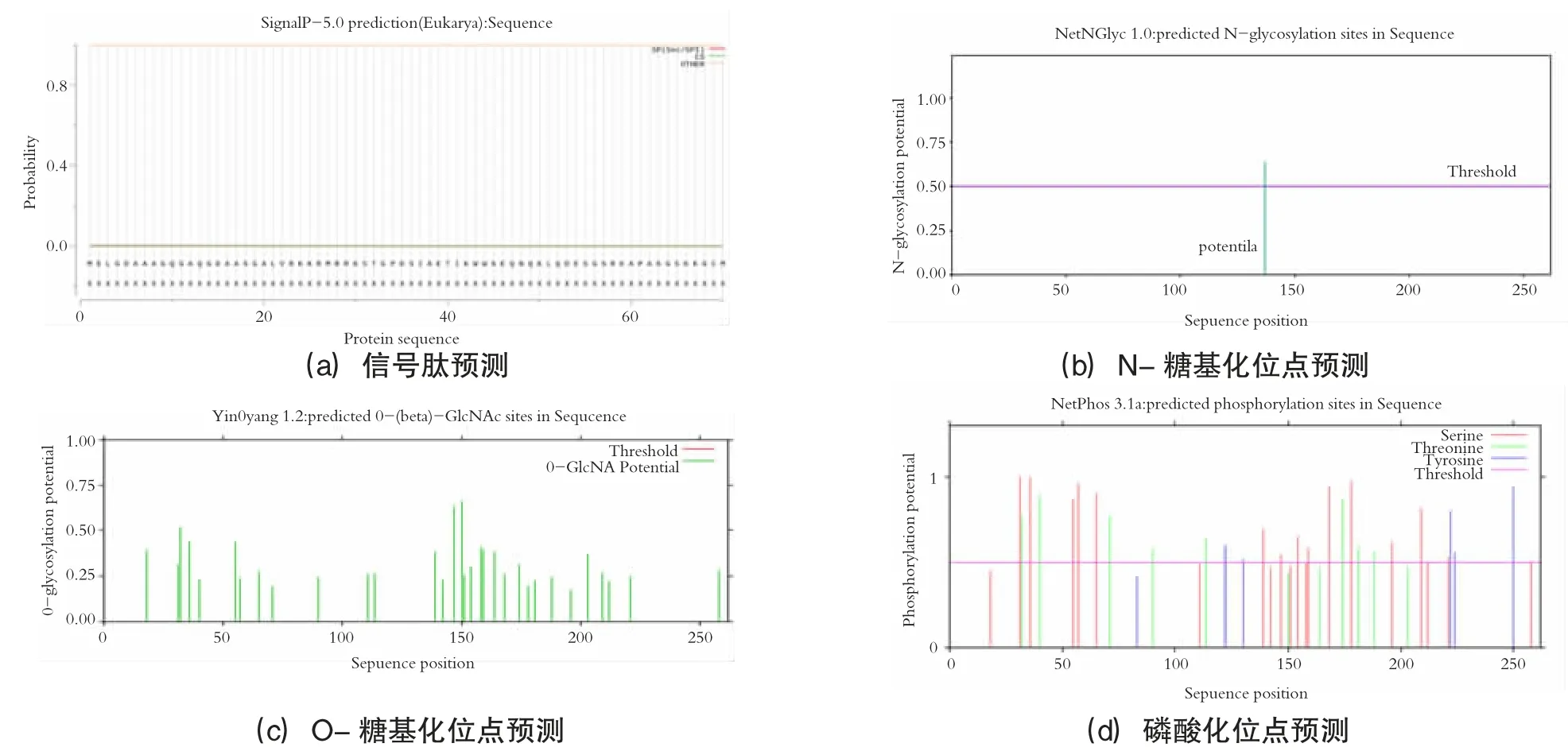
图6 甘蔗ScDREB2 转录因子活性位点预测
分析图6(d) 可知,其存在30 个磷酸化位点,其中17 个是丝氨酸(Ser),8 个是苏氨酸(Thr),5 个是酪氨酸(Tyr)。
2.3.4 甘蔗ScDREB2 转录因子结构预测
甘蔗DREB2 类转录因子的二级结构预测见图7;甘蔗DREB2 类转录因子的二级结构预测分析见表1。
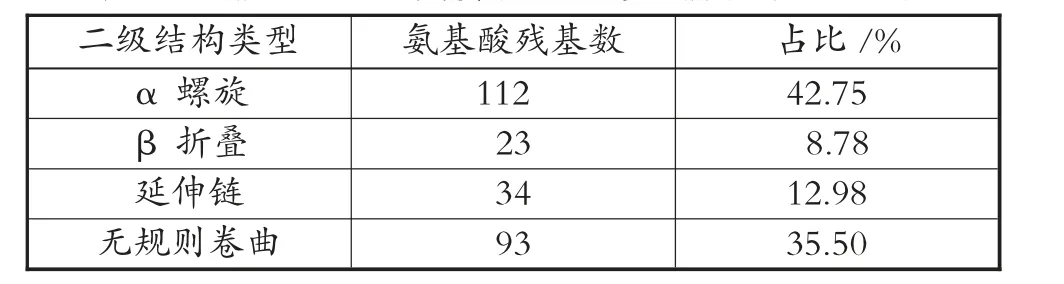
表1 甘蔗DREB2 类转录因子的二级结构预测分析

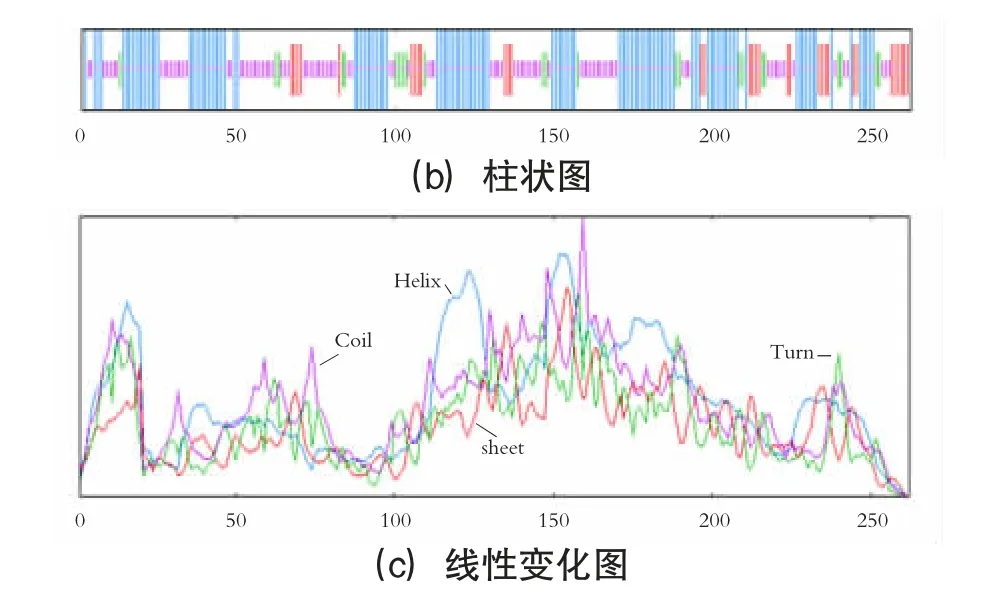
图7 甘蔗DREB2 类转录因子的二级结构预测
显示该蛋白α 螺旋占比最高,占比为42.75%;其次为无规则卷曲、延伸链、β 折叠,占比分别为35.50%、12.98%、8.78%。建模以5wx9.1.A 序列作为模板,构建甘蔗DREB2 类转录因子三级结构,与目标序列的一致度为58.33%,远大于30%,满足同源建模的基本条件。甘蔗DREB2 类转录因子三级结构预测见图8。
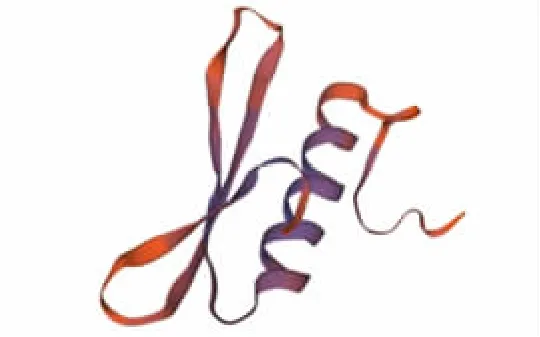
图8 甘蔗DREB2 类转录因子三级结构预测
3 分析与讨论
从DREB2 类转录因子首次被提取出来至今,短短几十年已经从众多植物中分离出DREB 转录因子,DREB 转录因子能够与DRE/CRT 顺式作用元件或具有DRE 元件核心序列(即TACCGACAT,其中CCGAC 是能特异性结合的最小序列[7]) 特异性结合,进而调控与干旱、高盐、高渗、极端温度等非生物胁迫应答有关的基因表达[8],通过这些基因产物的表达以及协同作用综合提高植物对逆境的抵抗能力[9]。
从种种事例来看,利用转基因技术改良甘蔗遗传性状,可以显著提高甘蔗抗旱性,具有良好的应用前景,是培育甘蔗良种的有力方法。仍存在转基因技术尚未完全成熟、遗传转化率不高等问题,转入的DREB 基因是否会对其他基因造成影响,目前也尚未知晓。但是转基因技术依旧是培养良种的有力手段,未来,随着测序技术发展,充分挖掘与了解DREB 转录因子,可提供充足的后选基因,利用基因工程,对培育抗旱性良好的甘蔗品种具有重要意义。
猜你喜欢
儿童故事画报·自然探秘(2022年12期)2022-11-24
基层中医药(2021年3期)2021-11-22
现代畜牧科技(2021年6期)2021-07-16
小猕猴学习画刊(2019年8期)2019-09-16
特别健康(2018年3期)2018-07-04
浙江农业学报(2017年3期)2017-04-08
中国烟草学报(2016年1期)2016-11-16
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
医学研究杂志(2015年12期)2015-06-10
中国粮油学报(2014年7期)2014-02-06
