
豆粕替代鱼粉添加丁酸梭菌对大口黑鲈生长性能、体组成和抗氧化能力的影响
2024-01-01 00:00:00矣林圆桂聪刘婧杨慧君莫爱杰翟昱翔袁勇超
华中农业大学学报 2024年1期


摘要 为研发大口黑鲈(Micropterus salmoides)优质配合饲料,以初始体质量(50.78±1.15) g 的大口黑鲈幼鱼为研究对象,设置对照组鱼粉含量为30%,在对照组的基础上使用豆粕分别替代25%、50% 和75% 鱼粉记为D25、D50 和D75 处理组,并在3 种豆粕替代基础上添加0.05% 丁酸梭菌设计D25+C、D50+C 和D75+C 3 个处理组,共计7 种试验饲料,进行为期8 周的饲喂试验,研究豆粕替代鱼粉并添加益生菌后鱼体的生长和抗氧化能力差异。结果显示:豆粕替代25% 鱼粉时大口黑鲈的生长性能、体组成和抗氧化能力均未出现显著差异。但替代比例达到50% 后大口黑鲈的增重率和特定增长率均显著降低,饲料系数显著升高,肝脏和血清总抗氧化能力、活性氧和超氧化物歧化酶均发生显著变化。然而,在替代50% 鱼粉的基础上添加丁酸梭菌组与对照相比,增重率、特定增长率、活性氧和超氧化物歧化酶均无显著差异。当替代水平达到75% 时添加丁酸梭菌与未添加组相比大口黑鲈的生长性能和抗氧化性能得到改善,但仍显著低于对照组。结果表明添加0.05% 的丁酸梭菌可改善豆粕替代鱼粉比例达到50% 对大口黑鲈生长性能和抗氧化能力造成的负面影响。
关键词 大口黑鲈; 配合饲料; 饲料添加剂; 丁酸梭菌; 豆粕替代; 生长性能; 抗氧化性能
中图分类号 S965.211 文献标识码 A 文章编号 1000-2421(2024)01-0176-09
随着水产养殖技术和经济效益的不断提升,我国对鱼粉的需求量也越来越高,已成为鱼粉进口量最大的国家[1]。如何解决鱼粉等优质蛋白源紧缺以及养殖成本高等一系列问题越来越重要,寻找优质蛋白源替代饲料中的鱼粉也成为水产饲料行业的热门话题[2]。豆粕作为植物蛋白源具有蛋白含量相对较高、来源广泛以及价格较低等优点[3],成为替代鱼粉的优质蛋白源。然而,豆粕中含有蛋白酶抑制剂、抗维生素和致敏原等多种抗营养因子[4],会损伤水产动物对饲料营养物质的吸收功能,从而降低生长性能[5]。有研究显示,在豆粕等植物蛋白源替代鱼粉的同时补充饲料添加剂,可以有效改善植物蛋白源替代过程中造成的负面影响,以提高植物蛋白源在饲料中的使用比例[6]。益生菌作为有效饲料添加剂具有促进鱼类生长、提高饲料利用率和机体免疫力等优点[7]。丁酸梭菌(Clostridium butyricum)能厌氧生长并产生芽孢,具有耐热、耐酸、耐碱及耐高温等特性,属于芽孢杆菌种类的益生菌[8],添加到饲料中可以有效改善机体的生长性能和消化酶活性,显著升高养殖对象的抗氧化能力[9],并对鱼体的免疫能力和肠道健康均有一定的促进作用[10]。但丁酸梭菌作为益生菌在提高豆粕替代鱼粉比例中的研究较为少见。
大口黑鲈(Micropterus salmoides)因具有生长速度快、适应低温、营养物质含量高等优点,在我国广泛养殖[11]。然而目前对大口黑鲈营养需求量及饲料策略的评估严重滞后于产业发展,在配合饲料研发方面仍然存在局限和挑战[12]。本研究以豆粕替代鱼粉,并在含豆粕饲料中添加丁酸梭菌,从生长性能和抗氧化能力等方面探究大口黑鲈对豆粕的最适耐受量及益生菌的改善作用,旨在为探索豆粕等植物性蛋白源在大口黑鲈优质配合饲料中的使用策略及提高其利用效率提供一定的理论依据。
1 材料与方法
1.1 饲料制备
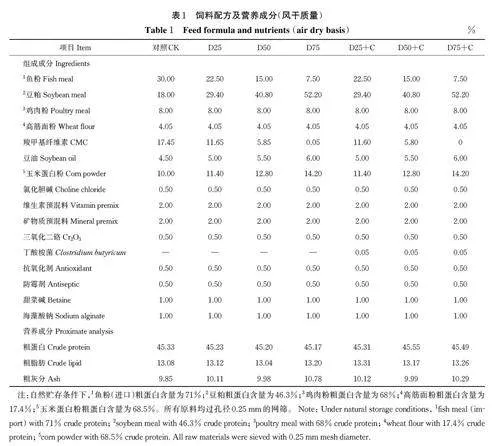
以鱼粉、鸡肉粉和豆粕为蛋白源,高筋面粉作为碳水化合物设计饲料,试验饲料配方如表1 所示。维生素预混料组成成分(mg/kg):维生素A 16 000 IU;维生素D3 8 000 IU;维生素B1 17.60;维生素B2 48.00;维生素B6 30.00;维生素 B12 0.25;维生素E156;维生素C 800;维生素 K3 14.70;烟酸胺79.00;泛酸钙73.50;叶酸6.50;生物素 0.64。矿物质预混料组成成分(mg/kg):镁(MgSO4 ·H2O) 52.50;锌(ZnSO4·5H2O) 34.40;铜(CuSO4·5H2O) 2.10;锰(MnSO4·7H2O) 6.10;碘(Ca(IO3)2)1.60;铁(FeSO4·7H2O) 21.10;硒(Na2 SeO3 ) 0.19;钴(CoCl2·5H2O) 0.24。共设计7 组饲料,设计对照组鱼粉的水平为30%,用豆粕分别替代25%、50% 和75% 的鱼粉记为D25、D50 和D75 组,并在各替代组添加0.05% 丁酸梭菌(蔚蓝生物股份有限公司,青岛;活菌数为2×1010 cuf/g)分别记为D25+C、D50+C 和D75+C 组。饲料原料由武汉澳华农牧科技有限公司提供,依据饲料配方表准确称取各组饲料原料过孔径0.25 mm 网筛后充分混匀。按饲料与水质量10∶3 的比例量取蒸馏水,将丁酸梭菌溶于水中搅拌均匀后混入饲料制成面团状,通过球团机(霍巴特混合机,型号A200)90~110 ℃压制成3 mm 直径的颗粒饲料,置于烘干机中45 ℃烘干,封于自封袋,-20 ℃冰箱中保存备用。
1.2 供试鱼及饲养管理
大口黑鲈购自湖北黄优源渔业发展有限公司,试验于华中农业大学水产基地的玻璃养殖缸(60cm×80 cm×60 cm)中进行。玻璃缸清洗并用高锰酸钾浸泡消毒后,注入新水,每个养殖缸放置1 个充气石,增气泵24 h 持续增氧,采用静水养殖。养殖过程管理与同一课题组桂聪等[13]的试验一致,每天换水40%~50%,以维持养殖水体的健康状态。选取规格相近、生长状态良好的初始体质量(50.78±1.15) g 的大口黑鲈420 尾,随机放入21 个玻璃缸中,每缸20 尾。供试鱼在玻璃缸中暂养2 周以适应养殖环境。暂养结束后,每组饲料随机投喂于3 个缸中。每天08:00 和18:00 进行投喂前,使用虹吸管吸取玻璃养殖缸中的粪便和残饵,保持良好的养殖水体环境,投喂量为鱼体质量的2%~5%,每天记录摄食量。水温维持在25~28 ℃,溶氧大于5.20 mg/L,氨氮为(0.25±0.12) mg/L,pH 为7.2±0.2,玻璃缸的水位保持在45 cm,饲养试验进行8 周。
1.3 样品采集
8 周试验结束后,将鱼禁食24 h,使用MS-222 溶液进行麻醉处理,记录每个缸中鱼体终末体质量、存活数,并测量体长。每个玻璃缸随机选取3 尾鱼作为全鱼样放入-80 ℃冰箱冷冻保存。再挑选3 尾鱼无菌操作下尾静脉采血后分离出内脏和肝脏并称质量、计算形体学指标,其余供试鱼取肝脏及肌肉组织液氮速冻后放入-80 ℃冰箱保存,用于抗氧化酶活性和常规营养成分的检测。血液样品在37 ℃恒温水浴30 min,3 000 r/min 离心15 min,分离出血清,-80 ℃冰箱保存用于检测抗氧化酶活性。
1.4 测定指标与计算方法
试验结束后对大口黑鲈末体质量(final weight,FBW)、增重率(weight gain rate, WGR)、特定生长率(special growth rate, SGR)、饲料系数(feed conversionratio, FCR)、存活率(survival rate, SR)、脏体比(viscerosomatic index, VSI)、肝体比(hepatosomaticindex, HSI)、肥满度(condition factor, CF)、摄食量(feed intake, FI)等指标参数进行测定,各指标具体的计算方法参考桂聪等[13]。
将恒温烘箱设定在105 ℃测定饲料、全鱼和肌肉中的水分含量,样品酸化消煮后使用全自动凯氏定氮分析仪测定粗蛋白含量,粗脂肪使用索氏抽提法测定,马弗炉550 ℃高温灼烧测定粗灰分。选取一定量的肝脏组织加入预冷生理盐水以体积比1∶9 制成匀浆,4 ℃下3 000 r/min 离心10 min,取离心后上层清液,按一定比例稀释后用于后续抗氧化指标活性分析。血清和肝脏中超氧化物歧化酶(SOD)活性(A001-3)、过氧化氢酶(CAT)活性(A007-1-1)、总抗氧化能力(T-AOC)(A015-2-1)、丙二醛(MDA)含量(A003-1)、活性氧(ROS)水平(E004-1)测定采用南京建成生物工程研究所的试剂盒,具体测定方法见试剂盒说明书。
1.5 数据处理
按照相关指标的计算公式对所测得的数据进行运算整理,再使用 SPSS Statistics 25 软件进行单因素方差分析(One-way ANOVA),多重差异显著性比较用Duncan’s 法,显著水平α=0.05,所有数据表示为“平均值±标准差”。
2 结果与分析
2.1 豆粕替代鱼粉添加丁酸梭菌对大口黑鲈生长性能的影响
由表2 可知,大口黑鲈末体质量随豆粕替代水平的上升而下降,替代水平达到50% 时出现显著性变化(Plt;0.05),添加丁酸梭菌后得到明显改善。豆粕替代水平达到50%,大口黑鲈的WGR 和SGR 显著性低于对照组(Plt;0.05),同时FCR 出现显著升高(Plt;0.05),添加0.05% 的丁酸梭菌后大口黑鲈的WGR、SGR 和FCR 得到明显改善,并与对照组相比无显著差异(Pgt;0.05),但75% 替代组添加丁酸梭菌后仍与对照组存在显著差异(Plt;0.05)。豆粕替代鱼粉以及添加丁酸梭菌对大口黑鲈的SR 和FI 未造成显著影响(Pgt;0.05)。
2.2 豆粕替代鱼粉并添加丁酸梭菌对大口黑鲈体组成和形体学指标的影响
由表3 可知,豆粕替代鱼粉未对大口黑鲈肌肉和全鱼的水分、粗蛋白、粗脂肪和粗灰分等营养成分造成显著影响(Pgt;0.05)。在豆粕替代的基础上添加丁酸梭菌,全鱼和肌肉中粗蛋白的含量呈现一定的升高趋势,表明丁酸梭菌使鱼体对饲料中蛋白质的沉积率得到提升,但仍低于对照组。
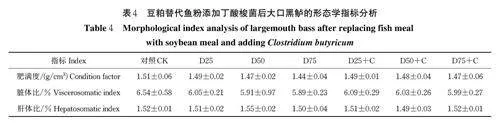
由表4 可知,豆粕替代不同比例的鱼粉以及添加丁酸梭菌对大口黑鲈的肥满度(CF)、脏体比(VSI)和肝体比(HSI)都未造成显著差异(Pgt;0.05)。大口黑鲈的肥满度随豆粕替代比例的升高总体呈下降趋势,通过添加丁酸梭菌在一定程度上得到改善,但仍低于对照组。
2.3 豆粕替代鱼粉并添加丁酸梭菌对大口黑鲈体肝脏和血清抗氧化能力的影响
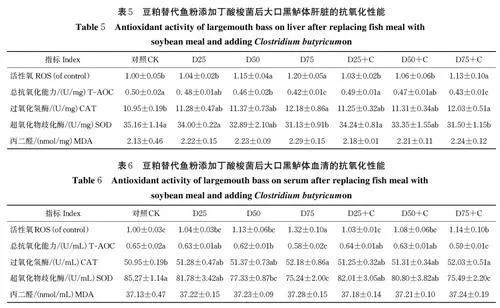
从表5 可知,豆粕替代鱼粉比例达到50% 时,大口黑鲈肝脏活性氧含量显著升高,添加丁酸梭菌可以降低活性氧含量,但当替代比例达到75%,即使添加丁酸梭菌也未能显著降低活性氧含量。超过50%的豆粕替代比例会显著降低肝脏T-AOC(Plt;0.05),补充丁酸梭菌后T-AOC 与对照组相比无显著差异(Pgt;0.05);然而豆粕替代75% 鱼粉时,即使添加丁酸梭菌T-AOC 仍显著低于对照组(Plt;0.05)。与对照组相比,豆粕替代水平为75% 的组及补充丁酸梭菌组的肝脏CAT 和SOD 活性发生显著变化(Plt;0.05),其他处理组无显著差异(Pgt;0.05)。豆粕替代鱼粉以及添加丁酸梭菌对大口黑鲈肝脏MDA 含量无显著影响(Pgt;0.05)。
由表6 知,大口黑鲈血清活性氧含量在豆粕替代水平达到75% 时显著上升,添加丁酸梭菌后有一定的下降但仍与对照组存在显著差异(Plt;0.05)。替代水平达到50% 时大口黑鲈血清T-AOC 与对照相比显著下降(Plt;0.05),添加丁酸梭菌后得到明显改善且与对照组相比无显著差异(Pgt;0.05)。75% 的豆粕替代鱼粉会显著改变血清CAT 和SOD 活性,添加丁酸梭菌有一定程度的改善但仍与对照组存在显著差异(Plt;0.05)。各组血清MDA 含量均无显著差异(Pgt;0.05)。
3 讨论
本研究结果显示,豆粕替代25% 的鱼粉时,大口黑鲈的生长性能和饲料利用率未被显著影响,但豆粕替代水平达到50% 后,大口黑鲈生长性能明显降低。其他鱼类也有类似报道,王赛等[14] 研究发现豆粕替代20% 鱼粉时未对褐点石斑鱼(Epinephelus fus⁃coguttatus)幼鱼的生长性能和摄食造成显著影响,但当豆粕替代比例到达一定值后,其生长受到抑制。大口黑鲈因具食欲强且抢食凶狠的食性特点,可能对豆粕替代引起的适口性变化敏感性较差,因此各试验组摄食量无显著变化。但高比例的豆粕中大量的抗营养因子可能会使大口黑鲈对饲料中营养物质的利用和吸收能力下降。然而,本研究发现豆粕替代50% 鱼粉时添加丁酸梭菌,能够抵抗豆粕对大口黑鲈生长抑制的作用,还能在一定程度上提高饲料的利用效率。Poolsawat 等[15]对罗非鱼(Oreochro⁃mis niloticus×O. aureus)的研究也有相似的结果。丁酸梭菌可产生蛋白酶、淀粉酶等多种消化酶,将多聚糖转化为低聚糖,从而提高机体的消化吸收功能,同时产生的某些寡糖有益于肠道益生菌的生长繁殖,以对肠道微生态起到调节作用,促进动物肠道的全面吸收功能[16]。但当替代水平达到75% 时即使添加丁酸梭菌,大口黑鲈生长性能得到改善,但仍显著低于对照组,可能因为本试验中丁酸梭菌的添加量较低而限制了改善效果。饲料抗营养因子水平、不同物种及自身健康状况、添加量等因素可能会影响丁酸梭菌等益生菌的饲用效果,如在Zhang 等[17]对肉鸡和朱振祥[18]对鲤(Cyprinus carpio L.)的研究报道中,饲料添加丁酸梭菌类制剂,未能显著提高动物的生长性能。
饲料营养组成和原料的不同,会对养殖对象的形态学指标和机体营养成分造成一定的影响。有研究显示,日本尖吻鲈(Lateolabrax japonicus)[19]和石斑鱼(Epinephelus lanceolatus)[20]饲料豆粕水平过高后,鱼体粗蛋白和粗脂肪成分显著降低。高水平的豆粕使机体粗蛋白出现下降趋势,这可能是饲料中不平衡的氨基酸组成造成的,大部分氨基酸被代谢消耗提供能量,用于蛋白质合成的氨基酸减少,从而降低了鱼体粗蛋白质沉积。水生动物对豆粕的耐受性具有物种特异性,在本研究中鱼体营养成分未出现显著变化,表明大口黑鲈鱼可能对豆粕替代的耐受性较强。豆粕替代的基础上添加丁酸梭菌未对大口黑鲈的体形造成显著影响,对其体组成具有一定的提升作用,但均未引起显著变化,这与兰菲菲[8]对鳗鲡的研究结果相似。提高饲料丁酸梭菌添加比例能否显著增加机体营养成分比例有待进一步研究。
豆粕在抵抗动物机体对其消化吸收的同时,也可能通过降低机体抗氧化能力而影响动物生长。抗氧化酶系统是当机体受到外界刺激后发生氧化反应进行调控的防御机制。常见的抗氧化酶系统中SOD和CAT 起着重要作用,SOD 可以对自由基进行清除以及抑制自由基的连锁反应[21],其反应过程产生的H2O2 可通过CAT 反应转化为H2O 和O2[22],达到保护机体的目的。有害刺激会增加机体活性氧含量,使得机体氧化-抗氧化作用失衡,发生氧化应激,导致脂质过氧化。MDA 是脂质过氧化物分解后的主要产物[23],T-AOC 体现了机体在发生脂质过氧化物过程中的抑制能力。当豆粕替代水平达到50%,大口黑鲈肝脏中活性氧和T-AOC 发生显著变化,当豆粕的替代水平达到75% 时,大口黑鲈肝脏和血清中T-AOC、CAT、SOD 等抗氧化酶活力及活性氧含量显著改变。当前已有研究显示,豆粕替代鱼粉水平过高会导致齐口裂腹鱼的抗氧化性能显著降低[24],而奥尼罗非鱼(Oreochromis niloticus × O. au⁃reus)肝脏SOD 也随饲料豆粕水平上升而降低[25]。
另外,机体抗氧化作用有多层次相互协同的特点,抗氧化酶活性降低,则非酶系统的抗氧化剂可能加强对多余活性氧的清除[26],活性氧的清除及细胞代谢过程会产生大量H2O2,H2O2 会刺激CAT 对其进行分解,可能因此而保持了高水平的CAT 活性。这可能是本研究中肝脏及血清SOD 与CAT 活性在不同豆粕替代水平下呈现相反趋势的原因。机体抗氧化系统中SOD、CAT 及谷胱甘肽过氧化物酶等抗氧化酶在克服氧化应激升高的酶活性间有代偿性增加效应[27],但这在豆粕替代鱼粉对大口黑鲈的胁迫应激模型中还需要更深入的研究。与此同时,CAT 的强抗氧化性,抑制了脂质过氧化,降低了细胞氧化损伤[28],从而减少了MDA 的产生,这可能表现为高豆粕替代组肝脏和血清MDA 与对照组相比未出现显著差异。有研究表明,豆粕替代鱼粉水平对点带石斑鱼(Epinephelus coioiaes)[29]血清MDA 及对牛蛙[30]肝脏MDA 未产生显著影响,与本研究结果一致。
丁酸梭菌不仅可保护或增强肠道吸收功能,还可能通过提高机体抗氧化能力从而抵抗豆粕对机体产生的氧化胁迫进而维持动物健康生长。在豆粕替代鱼粉的基础上添加丁酸梭菌,大口黑鲈的抗氧化能力得到明显改善,尤其当替代比例达到50% 时,添加丁酸梭菌相比于对照组无显著差异。但当豆粕替代水平为75% 时大口黑鲈的抗氧化性能虽得到改善但仍显著低于对照组。而将益生菌添加到日本囊对虾(Marsupenaeus japonicus)[31]饲料中也出现类似现象。丁酸梭菌在机体内产生的还原型辅酶过氧化物酶和SOD,可对机体内的活性氧进行直接清除[32]。
丁酸梭菌还可能通过增强CAT 活性,而降低组织MDA 含量[33]。但在过高的豆粕胁迫下,丁酸梭菌未能对机体氧化性能起明显的改善作用,可能原因是丁酸梭菌的添加量及其产生的抗氧化作用未能抵抗高水平的抗营养因子对机体产生的氧化应激,进一步提高丁酸梭菌添加量是否会对生长抑制或应激状态的动物机体产生明显的改善效果,还有待更深入的研究。
综上所述,豆粕可能通过影响肠道对饲料营养物质的消化吸收,降低机体抗氧化性能进而抑制动物机体生长,丁酸梭菌在保护肠道吸收功能及增强氧化应激能力方面发挥作用进而改善动物生长性能。在本研究条件下,50% 豆粕替代鱼粉的饲料中添加0.05% 的丁酸梭菌,可维持大口黑鲈的健康生长,降低饲料成本。
参考文献References
[1] 唐启升,丁晓明,刘世禄,等. 我国水产养殖业绿色、可持续发
展保障措施与政策建议[J]. 中国渔业经济,2014,32(2):5-
11.TANG Q S,DING X M,LIU S L,et al.Safeguard measures
and policy recommendations for green and sustainable development
of Chinese aquaculture[J]. Chinese fisheries economics,
2014,32(2):5-11( in Chinese with English abstract).
[2] 代伟伟,麦康森,徐玮,等. 复合植物蛋白源替代鱼粉对半滑
舌鳎生长、生理生化指标和肠组织结构的影响[J]. 中国水产
科学,2016,23(1):125-137.DAI W W,MAI K S,XU W,et
al. Effects of replacing fish meal with plant-based protein on
growth,physiological and biological indices,and intestinal histology
in tongue sole,Cynoglossus semilaevis Güntuer[J].Journal
of fishery sciences of China,2016,23(1):125-137( in Chinese
with English abstract).
[3] AZARM H M,LEE S M.Effects of partial substitution of di‑
etary fish meal by fermented soybean meal on growth performance,
amino acid and biochemical parameters of juvenile
black sea bream Acanthopagrus schlegeli[J]. Aquaculture research,
2014,45(6):994-1003.
[4] SOOKYING D,DAVIS D A,SOLLER DIAS DA SILVA
F. A review of the development and application of soybeanbased
diets for Pacific white shrimp Litopenaeus vannamei[J].
Aquaculture nutrition,2013,19(4):441-448.
[5] GEMEDE H F. Antinutritional factors in plant foods: potential
health benefits and adverse effects[J]. International journal
of food sciences and nutrition, 2014, 3(4): 284-289.
[6] 李维康,李立贤,刘泓宇,等. 低鱼粉饲料添加丁酸梭菌对凡
纳滨对虾生长、抗氧化能力和非特异性免疫的影响[J]. 广东
海洋大学学报,2022,42(2):29-37.LI W K,LI L X,LIU H
Y,et al.Effects of Clostridium butyricum on growth,antioxidant
capacity and non-specific immunology of Litopenaeus van⁃
namei fed with concentrated cottonseed protein replacement of
fishmeal[J].Journal of Guangdong Ocean University,2022,42
(2):29-37( in Chinese with English abstract).
[7] 林艾影,王维政,陈刚,等.2 种乳酸菌对军曹鱼幼鱼生长及消
化酶、免疫酶活性的影响[J]. 广东海洋大学学报,2020,40
(5):112-117.LIN A Y,WANG W Z,CHEN G,et al.Effects
of two lactic acid bacteria on growth performance and activities
of digestive and non-specific immune enzymes of juvenile cobia
(rachycentroncanadum)[J]. Journal of Guangdong Ocean
University,2020,40(5):112-117( in Chinese with English abstract).
[8] 兰菲菲. 饲料中添加丁酸梭菌对鳗鲡生长与健康的促进作用
[D]. 厦门:集美大学,2019.LAN F F.Effect of Clostridium
butyrate added to feed on the growth and health of eel[D].Xiamen:
Jimei University,2019 (in Chinese with English abstract).
[9] 何瑞鹏,奉杰,田相利,等. 酪酸菌对珍珠龙胆石斑鱼生长、消
化酶、血清抗氧化酶和溶菌酶活性的影响[J]. 中国海洋大学
学报(自然科学版),2017,47(11):15-23.HE R P,FENG J,
TIAN X L,et al. Effects of dietary supplementation of Clos⁃
tridium butyricum on the growth and activities of digestive and
serum antioxidant enzymes and lysozyme of hybrid grouper
(Epinephelus fuscoguttatus ♀ × E. lanceolatus ♂)[J].Periodical
of Ocean University of China(natural science edition),
2017,47(11):15-23( in Chinese with English abstract).
[10] 王海瑞,莫文艳,赵红霞,等. 饲料添加丁酸梭菌对黄颡鱼生
长性能及血清生化指标、免疫功能和抗氧化能力的影响[J].
动物营养学报,2022,34(8):5295-5303. WANG H R,MO
W Y,ZHAO H X,et al.Effects of adding Clostridium butyri⁃
cum in diets on growth performance,serum biochemical indices,
immune function and antioxidant ability of yellow catfish
(Pelteobagrus fulvidraco)[J].Chinese journal of animal nutrition,
2022,34(8):5295-5303 (in Chinese with English abstract).
[11] 麻艳群,司楠,介百飞,等. 不同投饲率对陆基圆池养殖大口
黑鲈生长及养殖效能的影响[J]. 渔业研究,2022,44(3):
223-228. MA Y Q,SI N,JIE B F,et al. Effects of different
feeding rates on growth and efficiency of land-based circular
pond culture Micropterus salmoides[J].Journal of fisheries research,
2022,44(3):223-228 (in Chinese with English abstract).
[12] 林仕梅,陈拥军,周文豪,等. 大口黑鲈高质量发展之营养调
控策略[J]. 饲料工业,2022,43(10):12-17.LIN S M,CHEN
Y J,ZHOU W H,et al. Nutritional regulation strategies for
high-quality development of largemouth bass[J]. Feed industry,
2022,43(10):12-17( in Chinese with English abstract).
[13] 桂聪,邓琦琦,杨慧君,等. 低蛋白饲料添加蛋氨酸和赖氨酸
对大口黑鲈生长性能和抗氧化能力的影响[J]. 华中农业大
学学报,2022,41(6):184-191.GUI C,DENG Q Q,YANG
H J,et al.Effects of methionine and lysine supplementation in
low protein diets on growth performance,body composition,
antioxidant capacity and immune enzymes of largemouth bass
(Micropterus salmoides)[J].Journal of Huazhong Agricultural
University,2022,41(6):184-191( in Chinese with English abstract).
[14] 王赛,陈刚,张健东,等. 不同蛋白质源部分替代鱼粉对褐点
石斑鱼幼鱼生长性能、体组成以及血清生化指标的影响[J].
动物营养学报,2012,24(1):160-167.WANG S,CHEN G,
ZHANG J D,et al. Influence of partial replacement of fish
meal by different protein sources on growth performance,body
composition and serum biochemical indices of juvenile Epi⁃
nephelus fuscoguttatus[J].Chinese journal of animal nutrition,
2012,24(1):160-167( in Chinese with English abstract).
[15] POOLSAWAT L,LI X Q,HE M,et al.Clostridium butyricu⁃
mas probiotic for promoting growth performance,feed utilization,
gut health and microbiota community of tilapia (Oreo⁃
chromis niloticus × O. aureus)[J]. Aquaculture nutrition,
2020,26(3):657-670.
[16] 胡文攀,朱振祥,李克克. 丁酸梭菌的生物学功能及在水产养
殖中的应用[J]. 当代水产,2022,47(1):76-77.HU W P,
ZHU Z X,LI K K.Biological function of Clostridium butyri⁃
cum and its application in aquaculture[J]. Current fisheries,
2022,47(1):76-77( in Chinese).
[17] ZHANG B K,YANG X,GUO Y M,et al.Effects of dietary
lipids and Clostridium butyricum on the performance and the
digestive tract of broiler chickens[J].Archives of animal nutrition,
2011,65(4):329-339.
[18] 朱振祥. 丁酸梭菌对鲤生长、免疫及肠道微生态的影响[D].
新乡:河南师范大学,2019.ZHU Z X.Effects of Clostridium
butyricum on growth,immunity and intestinal microecology of
common carp(Cyprinus carpio L.)[D].Xinxiang:Henan Normal
University,2019( in Chinese with English abstract).
[19] ZHANG C X,RAHIMNEJAD S,WANG Y R,et al.Substituting
fish meal with soybean meal in diets for Japanese sea‑
bass (Lateolabrax japonicus):effects on growth,digestive enzymes
activity,gut histology,and expression of gut inflammatory
and transporter genes [J]. Aquaculture,2018,483:
173-182.
[20] GARCÍA-ORTEGA A,KISSINGER K R,TRUSHENSKI
J T. Evaluation of fish meal and fish oil replacement by soybean
protein and algal meal from Schizochytrium limacinum in
diets for giant grouper Epinephelus lanceolatus[J]. Aquaculture,
2016,452:1-8.
[21] 蒋锦坤. 壳聚糖对虹鳟(Oncorhynchus mykiss)和星斑川鲽
(Platichthys stellatus)幼鱼生长及非特异性免疫的影响[D].
上海:上海海洋大学,2012.JIANG J K.Effects of chitosan on
growth and nonspecific immunity of juvenile rainbow trout
(Oncorhynchus mykiss) and Platichthys stellatus[D]. Shanghai:
Shanghai Ocean University,2012 (in Chinese with English
abstract).
[22] DORVAL J,HONTELA A. Role of glutathione redox cycle
and catalase in defense against oxidative stress induced by endosulfan
in adrenocortical cells of rainbow trout (Oncorhyn⁃
chus mykiss)[J].Toxicology and applied pharmacology,2003,
192(2):191-200.
[23] 申建飞,陈铭灿,刘泓宇,等. 浓缩棉籽蛋白替代鱼粉对卵形
鲳鲹幼鱼生长性能、血清生化指标、肝脏抗氧化指标及胃肠
道蛋白酶活性的影响[J]. 动物营养学报,2019,31(2):746-
756.SHEN J F,CHEN M C,LIU H Y,et al.Effects of fish
meal replacement by concentrated cottonseed protein on
growth performance,serum biochemical indices,liver antioxidant
indices and gastrointestinal tract protease activities of juvenile
Trachinotus ovatus[J].Chinese journal of animal nutrition,
2019,31(2):746-756 (in Chinese with English abstract).
[24] 向枭,周兴华,陈建,等. 饲料中豆粕蛋白替代鱼粉蛋白对齐
口裂腹鱼幼鱼生长性能、体成分及血液生化指标的影响[J].
水产学报,2012,36(5):723-731. XIANG X,ZHOU X H,
CHEN J,et al.Effect of dietary replacement of fish meal protein
with soybean meal protein on the growth,body composition
and hematology indices of Schizothorax prenanti[J].Journal
of fisheries of China,2012,36(5):723-731 (in Chinese
with English abstract).
[25] LIN S M,LUO L.Effects of different levels of soybean meal
inclusion in replacement for fish meal on growth,digestive enzymes
and transaminase activities in practical diets for juvenile
Tilapia,Oreochromis niloticus × O. aureus[J]. Animal feed
science and technology,2011,168(1/2):80-87.
[26] 王新民. 人体抗氧化防御系统浅谈[J]. 卫生职业教育,2011,
29(14):156-158.WANG X M.Discussion on human antioxidant
defense system[J].Health vocational education,2011,29
(14):156-158( in Chinese).
[27] LIMAYE P V,RAGHURAM N,SIVAKAMI S. Oxidative
stress and gene expression of antioxidant enzymes in the renal
cortex of streptozotocin-induced diabetic rats[J]. Molecular
and cellular biochemistry,2003,243(1):147-152.
[28] 朱安南,刘欢,孟勇,等. 过氧化氢酶的功能及在畜禽中的研
究进展[J]. 饲料研究,2022,45(10):143-145.ZHU A N,LIU
H,MENG Y,et al.Research progress on function of catalase
and its application in livestock and poultry production[J].Feed
research,2022,45(10):143-145 (in Chinese with English abstract).
[29] 安贸麟,范泽,王庆奎,等. 豆粕替代鱼粉对点带石斑鱼生长、
消化和抗氧化能力的影响[J]. 江苏农业科学,2018,46(16):
128-132.AN M L,FAN Z,WANG Q K,et al.Influences of dietary
replacement of fishmeal by soybean meal on growth performance,
digestive ability and antioxidant ability of Epinephe⁃
lus malabaricus[J]. Jiangsu agricultural sciences,2018,46
(16):128-132( in Chinese with English abstract).
[30] 方卫东,鲁康乐,张春晓,等. 豆粕替代鱼粉对牛蛙生长、体组
成、消化酶活力及肝脏生化指标的影响[J]. 水产学报,2016,
40(11):1742-1752.FANG W D,LU K L,ZHANG C X,et
al. Effects of fish meal replacement by soybean meal on
growth,body composition,digestive enzyme activities and hepatic
biochemical indices of Rana catesbeiana[J]. Journal of
fisheries of China,2016,40(11):1742-1752 (in Chinese with
English abstract).
[31] DUAN Y F,ZHANG Y,DONG H B,et al.Effect of the dietary
probiotic Clostridium butyricum on growth,intestine antioxidant
capacity and resistance to high temperature stress in
kuruma shrimp Marsupenaeus japonicus[J].Journal of thermal
biology,2017,66:93-100.
[32] 贾聪慧,杨彩梅,曾新福,等. 丁酸梭菌对肉鸡生长性能、抗
氧化能力、免疫功能和血清生化指标的影响[J]. 动物营养
学报,2016,28(3):908-915.JIA C H,YANG C M,ZENG
X F,et al.Effects of Clostridium butyricum on growth performance,
antioxidant capacity,immune function and serum biochemical
parameters of broilers[J].Chinese journal of animal
nutrition,2016,28(3):908-915 (in Chinese with English abstract).
[33] ZHANG W H,GAO F,ZHU Q F,et al.Dietary sodium butyrate
alleviates the oxidative stress induced by corticosterone exposure
and improves meat quality in broiler chickens[J].Poultry
science,2011,90(11):2592-2599.
(责任编辑:边书京)
