
不同规格绿鳍马面鲀耗氧率、排氨率与窒息点研究
2024-01-01 00:00:00杨哲李欣阳孟艳莎杨祯王淑慧高铭鸿曹新宇姜晨
水产科技情报 2024年4期

摘 要:为研究鱼体大小对绿鳍马面鲀耗氧率、排氨率以及窒息点的影响,利用静水封闭式呼吸室法,在水温22 ℃、盐度30条件下,测定了4种规格[体质量分别为(21.51±4.78)、(44.55±3.18)、(76.98±7.33)、(153.79±4.99)g]绿鳍马面鲀的耗氧率、排氨率和窒息点。结果显示,绿鳍马面鲀的耗氧率和排氨率均随着鱼体规格的增大呈现逐渐降低的趋势,且各组间的耗氧率均存在显著性差异(P<0.05);绿鳍马面鲀的窒息点随着鱼体规格的增大而提高,规格最小的试验鱼,其窒息点最低,且与规格较大的2个组有显著性差异(P<0.05),其余3组之间则差异不显著(Pgt;0.05);试验测得的氧氮比为6.27~18.53,表明在试验条件下绿鳍马面鲀主要的供能物质为蛋白质。试验结果表明,鱼体大小对绿鳍马面鲀的耗氧率、排氨率均有显著影响,但对其窒息点的影响不显著。
关键词:绿鳍马面鲀;规格;耗氧率;排氨率;窒息点
绿鳍马面鲀(Thamnaconus septentrionalis)俗称橡皮鱼、扒皮鱼、马面鱼等,隶属于鲀形目、单角鲀科、马面鲀属,因其面部较长、形似马面,且背鳍、臀鳍、尾鳍等鳍条呈现绿色而得名。该鱼分布于中国、朝鲜半岛以及日本沿海,在我国渤海、黄海、东海等海域均有分布,为外海暖温性底层鱼类,在冬季水温低时需要进行越冬洄游[1]。绿鳍马面鲀的鱼皮较厚,可用于制取明胶和提取胶原蛋白,其肝脏经加工提炼后可制成鱼肝油,其肌肉富含蛋白质,且脂肪含量较低,是良好的蛋白质来源[2-3]。在我国,绿鳍马面鲀的年捕捞产量最高曾达30万~50万t,但因过度捕捞,20世纪90年代后,其产量急剧衰减[4]。随着绿鳍马面鲀人工繁育技术的突破[5],以及近几年绿鳍马面鲀陆海接力养殖技术(即利用陆地工厂化养殖设施为深水大网箱提供大规格鱼种,完成养殖阶段的越冬,起到避集中上市、调节市场需求的作用)的发展等[6],绿鳍马面鲀产量在近年来得以恢复。
水中的溶解氧和氨氮浓度能较好地反映鱼体呼吸和代谢状况的指标。过低的溶解氧会抑制鱼体新陈代谢,降低鱼体活力,甚至造成鱼体死亡。氨态氮浓度不仅可以反映鱼体代谢水平,而且作为水体环境中的主要污染物,其浓度过高会导致鱼体中毒。窒息点则反映了鱼体所能耐受的最低溶解氧浓度。本试验测定了相同温度、盐度条件下4种规格绿鳍马面鲀的耗氧率、排氨率以及窒息点,探究鱼体大小与耗氧率、排氨率和窒息点的关系,以期掌握绿鳍马面鲀个体规格与代谢之间的规律,为开展集约化养殖提供参考。
1 材料和方法
1.1 试验材料
试验用绿鳍马面鲀来自大连天正实业有限公司,是由野生苗种在室内工厂化车间内养成的,试验鱼健康、无伤、行为正常。试验开始前,将试验鱼停饲1 d,使其体内食物排空,然后于室内放置的水槽中暂养,使用空调以及加热棒使水温逐渐升高至试验水温(22 ℃)。试验用海水盐度为30,海水来源为学校附近的黄海海域,采用自然光照。试验鱼的生物学数据见表1。
1.2 试验方法
1.2.1 耗氧率、排氨率以及窒息点的测定
根据预试验结果,共设置4种鱼体规格(见表1)。每种规格设置3个平行组,其中Ⅲ、Ⅳ组每个平行组试验鱼2尾,Ⅰ、Ⅱ组每个平行组试验鱼3尾。
本试验采用静水封闭式呼吸室法。试验所用容器为11 L玻璃瓶,瓶盖为磨砂口,可有效防止试验过程中空气进入。瓶中注满22 ℃的海水,并在水槽中使用加热棒以保持试验全程水温保持在22 ℃。放入试验鱼后,立即盖上瓶盖并开始计时,分别检测试验起始和1 h后(即终末)水体溶解氧和氨态氮质量浓度。每种规格3个平行组的试验结果取平均值。
使用美国YSI多参数水质分析仪检测试验起始、终末、窒息点以及试验鱼全部死亡时的溶解氧。使用纳氏试剂法测定氨态氮质量浓度。试验鱼死亡的判断依据为:口和鳃盖不再有呼吸,鳍条静止不动,鱼浮于水面或沉于水底静止不动。试验鱼死亡时的溶解氧质量浓度为窒息点。
1.2.2 计算方法
耗氧率(RO)和排氨率(RN)的计算公式如下。
RO=(CO0-COt)×v/(w×t) (1)
RN=(CN0-CNt)×v/(w×t) (2)
式(1)~(2)中,RO为耗氧率,CO0为起始溶解氧质量浓度(mg/L),COt为终末溶解氧质量浓度(mg/L),v为容量瓶体积(L),w为试验鱼体质量(g),t为试验时长(h),RN为排氨率,CN0为起始氨态氮质量浓度(μg/L),CNt为终末氨态氮质量浓度(μg/L)。
氧氮比(RO/N)是反映动物呼吸底物的重要参数,反映了水产动物在不同状态下对能源物质(糖类、脂肪和蛋白质)的代谢比例关系和对生命代谢底物的利用情况。氧氮比的计算公式如下。
RO/N=RO/RN(3)
式(3)中,RO/N为氧氮比,RO、RN分别为耗氧率和排氨率。
1.3 数据统计和分析
试验数据以平均值±标准差表示。采用SPSS 26.0软件对试验数据进行单因素方差分析,使用Duncan’s多重比较分析组间差异,设显著性水平为0.05。
2 结果
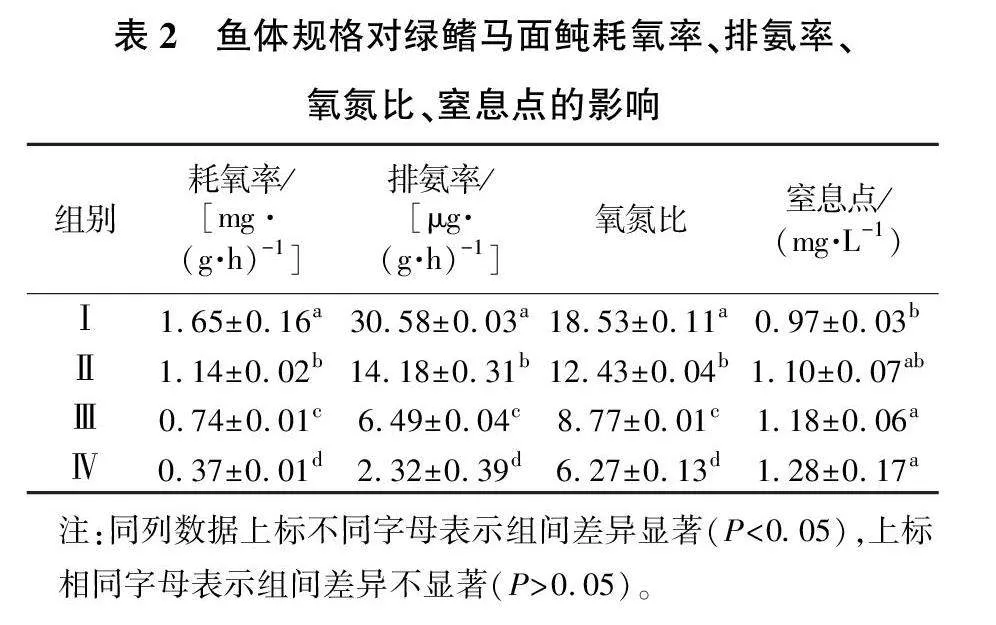
在水温22 ℃、盐度30条件下,4种规格绿鳍马面鲀的耗氧率、排氨率、氧氮比、窒息点见表2。
由表2可见,绿鳍马面鲀在单位时间内的耗氧率和排氨率均随着鱼体规格的增大呈现逐渐降低的趋势,且不同规格组间均存在显著性差异(P<0.05)。
绿鳍马面鲀的氧氮比随着鱼体规格的增大呈现逐渐减小的趋势,且不同规格绿鳍马面鲀的氧氮比之间存在显著性差异(P<0.05),说明鱼体大小对绿鳍马面鲀氧氮比的影响较为显著。
绿鳍马面鲀的窒息点随着鱼体规格的增大呈现逐渐升高的趋势,规格最小的第Ⅰ组,其窒息点最低,且与个体规格较大的Ⅲ、Ⅳ组有显著性差异(P<0.05)。据观察,在试验初始,试验鱼的行为正常;随着水体中的溶解氧逐渐降低,试验鱼开始在瓶内头朝上游动,之后侧翻;鱼体侧翻后,开始平躺于瓶底,随着溶解氧继续降低,试验鱼开始挣扎,头朝下尾朝上向上方游动,身体疯狂抽搐,不久便沉于瓶底死亡。
3 讨论
3.1 鱼体规格对耗氧率和排氨率的影响
鱼类作为一种低等脊椎动物,耗氧率能够真实地反映其代谢水平及速率。在相同养殖条件下,不同规格的同一种鱼,其耗氧率不同且存在规律性变化。本试验测定了4种规格绿鳍马面鲀的耗氧率,结果显示,耗氧率随着鱼体规格的增大呈现逐渐下降的趋势,这与花鲈[7]、珍珠龙胆石斑鱼[8]、翘嘴红鲌[9]等鱼类的相关研究结果基本一致。造成这种规律的原因可能是:(1)个体规格越小的鱼,其生长更为迅速,能量需求更高,代谢速度更快,会消耗更多的营养用于正常的生理活动,因此氧气的消耗量较高;(2)在幼鱼阶段,鱼类的脑、肝、肾、脾等维持生命活动的器官和组织占体质量的比例较大,而且这些器官相比于肌肉、骨骼等运动器官和组织消耗能量更多;(3)在成鱼阶段,肌肉、骨骼这些运动器官和组织消耗能量的比例逐渐增大,维持生命活动的组织消耗能量的比例反而减少,因此总体消耗的能量逐渐减少[10]。
排氨率也是能够反映鱼体代谢速率的指标。鱼类主要的排氨方式为蛋白质的代谢过程。本试验中,4种规格绿鳍马面鲀的排氨率随着鱼体规格的增大而逐渐降低,这与于宏等[11]对黄金鲫的研究结果相似。本试验排氨率的结果与耗氧率测定结果变化趋势相一致,可能说明鱼类蛋白质代谢与氧气消耗有关,个体规格越小的鱼,其所需的氧气也越多,耗氧率也越高,代谢较快,机体对蛋白质的消耗也随之增加,因而排氨率也随之升高。
3.2 氧氮比变化与鱼体规格的关系
氧氮比即耗氧率与排氨率的比值,其比值大小可以反映三大供能物质(糖类、脂肪和蛋白质)在鱼体代谢过程中的供能种类和供能占比[12]。有机体消耗能量的先后顺序依次为:糖类、脂肪、蛋白质,其中,单位质量的脂肪耗氧率较高并且在被消耗时释放能量较多,一般作为贮能物质,而依靠消耗蛋白质作为供能物质的情况并不常见。在鱼类中,糖类很少参与供能,在鱼体内主要以糖原的形式存在。
若全部由蛋白质作为氧化基质来供能,氧氮比约为7[13],若以蛋白质和脂肪作为氧化基质来供能,氧氮比约为24[14]。随着糖类和脂肪供给的能量逐渐增加,蛋白质便几乎不再充当能量供应的角色,氧氮比也会随之增大直至无穷大[15]。本试验中,个体规格最小的Ⅰ组,其氧氮比为18.53,而Ⅱ组、Ⅲ组与Ⅳ组,其氧氮比分别为12.43、8.77和6.27。这说明较大规格的鱼,其主要供能来源是蛋白质,而规格较小的鱼,其供能主要的来源为蛋白质和少量的脂肪。这与同为鲀形目的红鳍东方鲀[16]的氧氮比(12.416~14.208)较为接近。王波等[17]研究表明,牙鲆的氧氮比为38.8,其主要供能物质为糖类、脂肪和少量的蛋白质。根据本试验的结果,建议在绿鳍马面鲀养殖过程中,需要注意所投喂的饲料应保持较高的蛋白质和脂肪含量,并且要合理调控三种供能物质之间的比例。
3.3 鱼体规格对窒息点的影响
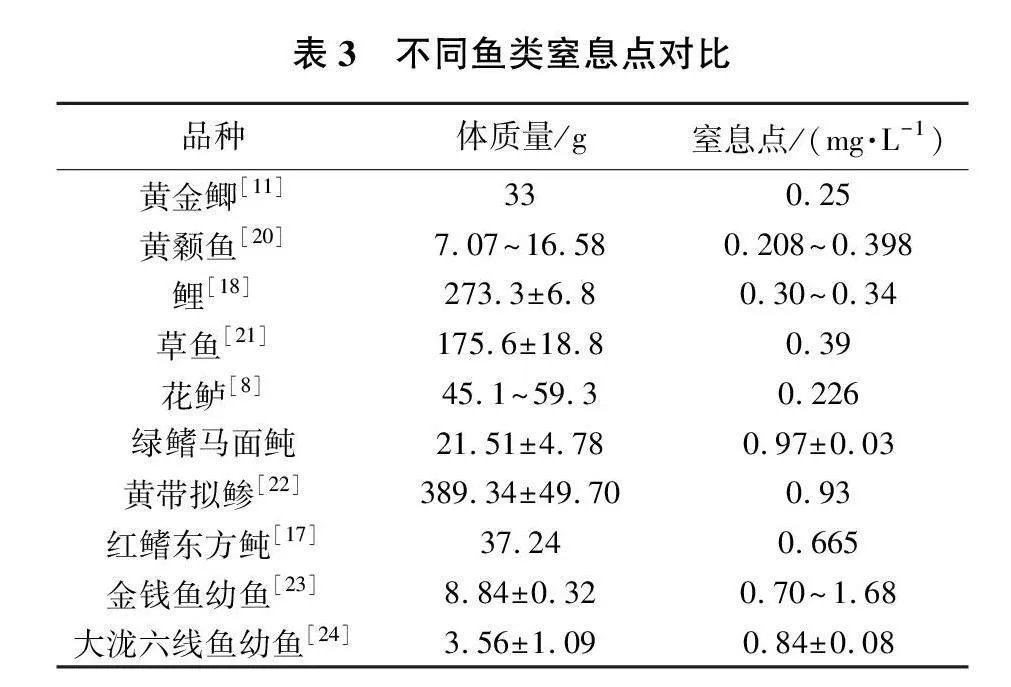
窒息点对于鱼类养殖、越冬以及运输均具有重要意义。不同种类的鱼,其鱼体大小与窒息点的关系也各有不同。例如,鲤[18]和湘云鲫[19]等鱼类的窒息点随着鱼体规格的增大而降低,而黄颡鱼[20]等鱼类的窒息点则随着鱼体规格的增大而升高。
本试验结果显示,绿鳍马面鲀的窒息点随着鱼体规格的增大而升高。可以看出,随着绿鳍马面鲀体质量的增加,其耐低氧能力也在提高,但总体来看,不同规格的鱼,其窒息点的差异并不显著。根据试验结果,建议绿鳍马面鲀在生产运输上应保证水体中的溶解氧保持在(0.97±0.03)mg/L以上。由表3[8,11,17-18,20-24]可见,绿鳍马面鲀、黄带拟鲹、红鳍东方鲀、金钱鱼和大泷六线鱼作为海水鱼,相比于表中其他淡水鱼类的窒息点要高。但鱼类的窒息点是否与海水盐度有关仍需进一步研究确认。
参考文献
[1]李刚,王庆龙,张凯强.绿鳍马面鲀的人工繁育技术[J].中国水产,2021(11):82-84.
[2]郁迪,丁冬各,王斌.绿鳍马面鲀(Navodon septentrionalis)鱼皮胶原蛋白的分离纯化及理化性质研究[J].海洋与湖沼,2016,47(5):1055-1062.
[3]徐大凤,刘琨,王鹏飞,等.绿鳍马面鲀肌肉营养成分分析和营养评价[J].海洋科学,2018,42(5):122-129.
[4]许学龙,郑元甲,刘松.东、黄海绿鳍马面鲀的资源评估[J].海洋与湖沼,1992,23(6):651-656.
[5]刘琨,刘刚,黄亮,等.绿鳍马面鲀大规格苗种网箱培育试验[J].渔业现代化,2019,46(6):54-60.
[6]胡丽萍,徐晓莹,张宁,等.绿鳍马面鲀陆海接力规模化养殖技术研究[J].水产养殖,2022,43(12):24-28.
[7]崔宽宽,尤宏争,丁子元,等.不同规格花鲈的耗氧率与窒息点研究[J].科学养鱼,2018(3):57-58.
[8]仇登高,黄种持,郑乐云,等.温度对不同规格珍珠龙胆石斑鱼(棕点石斑鱼♀×鞍带石斑鱼♂)呼吸和排泄的影响[J].中国水产科学,2017,24(2):295-305.
[9]雷建军,何安尤,施军,等.翘嘴红鲌鱼苗耗氧率和窒息点的观测[J].广西水产科技,2005(3):10-12.
[10]刘金兰,陈志伟,张家全,等.不同规格乌苏里拟鲿耗氧率及窒息点研究[J].天津农学院学报,2012,19(3):4-7.
[11]于宏,孙学亮,程民杰,等.不同规格黄金鲫耗氧率及排氨率研究[J].水产养殖,2015,36(1):10-14.
[12]王辉,强俊,王海贞,等.温度与盐度对吉富品系尼罗罗非鱼幼鱼能量代谢的联合效应[J].中国水产科学,2012,19(1):51-61.
[13]MAYZAUD P.Respiration and nitrogen excretion of zooplankton.IV.The influence of starvation on the metabolism and the biochemical composition of some species[J].Marine Biology,1976,37(1):47-58.
[14]IKEDA T.Nutritional ecology of marine zooplankton[J].Memoirs of the Faculty of Fisheries,1974,22:1-97.
[15]CONOVER R J,CORNER E D S.Respiration and nitrogen excretion by some marine zooplankton in relation to their life cycles[J].Journal of the Marine Biological Association of the United Kingdom,1968,48(1):49-75.
[16]何亚,王华,王伟,等.温度对不同体质量红鳍东方鲀幼鱼耗氧率和排氨率的影响[J].大连海洋大学学报,2014,29(5):481-485.
[17]王波,李继强,曹志海,等.大西洋牙鲆幼鱼标准代谢的初步研究[J].海洋科学进展,2004,22(1):62-68.
[18]周天倪,周洵,王世恒,等.鲤耗氧率与窒息点的测定[J].渔业经济研究,2006(4):37-41.
[19]刘飞,张轩杰,刘筠.湘云鲫耗氧率和溶氧临界窒息点[J].湖南师范大学学报(自然科学版),2000,23(3):72-75.
[20]陈琴,章太卓,徐夏声.黄颡鱼耗氧率与窒息点的初探[J].内陆水产,2001,26(3):9-11.
[21]余来宁,杨东,刘红艳,等.十二种淡水鱼类血红蛋白与窒息点的相关分析[J].动物学杂志,2017,52(3):478-484.
[22]杨祯,杨旭,高铭鸿,等.温度对黄带拟鲹耗氧率、排氨率及窒息点的影响[J].河北渔业,2021(7):1-4.
[23]蒋飞,徐嘉波,施永海.养殖水温对金钱鱼幼鱼耗氧率、排氨率和窒息点的影响[J].西北农林科技大学学报(自然科学版),2022,50(1):36-42.
[24]于燕光,逯云召,薄其康,等.不同温度对大泷六线鱼幼鱼耗氧率和窒息点的影响[J].河北渔业,2020(10):20-23.
Study on oxygen consumption rate,ammonia emission rate and asphyxia point of different sizes of Thamnaconus septentrionalis
YANG Zhe, LI Xinyang, MENG Yansha, YANG Zhen, WANG Shuhui, GAO Minghong, CAO Xinyu, JIANG Chen
(School of Fisheries and Life Science,Dalian Ocean University/Key Laboratory of Mariculture in Northern China,Ministry of Agriculture and Rural Affairs,Dalian 116023,China)
Abstract: To study the effects of fish body size on breath and metabolic status of Thamnaconus septentrionalis,the oxygen consumption rate,ammonia excretion rate and asphyxia point of T. septentrionalis with different sizes[(21.51±4.78),(44.55±3.18),(76.98±7.33) and (153.79±4.99) g] were determined by still water closed breathing chamber method.The results showed that the oxygen consumption rate and ammonia excretion rate of T. septentrionalis significantly decreased with fish size increasing(P<0.05).The asphyxia points of T. septentrionalis increased with the increase of fish body size,and the lowest level was observed in (21.51±4.78) g group(P<0.05),while there was no significant difference among other groups(Pgt;0.05).The oxygen-nitrogen ratio was 6.27~18.53,indicating that the main energy supply substance of T. septentrionalis under the test conditions was protein.The results showed that the fish body size had a significant effect on the oxygen consumption rate and ammonia excretion rate of T. septentrionalis,but had no significant effect on the asphyxia point.
Key words: Thamnaconus septentrionalis; size; oxygen consumption rate; ammonia excretion rate; asphyxia point
猜你喜欢
水产科技情报(2024年6期)2024-01-01 00:00:00
淡水渔业(2023年2期)2023-03-29 04:03:30
当代水产(2022年6期)2022-06-29 01:12:08
宝藏(2021年6期)2021-07-20 06:12:22
河北渔业(2019年7期)2019-08-27 04:01:48
山东冶金(2019年1期)2019-03-30 01:35:30
水产科学(2018年3期)2018-06-04 02:10:58
广东海洋大学学报(2015年3期)2015-12-22 10:05:30
应用海洋学学报(2014年1期)2014-11-22 07:17:50
浙江海洋大学学报(自然科学版)(2014年6期)2014-03-13 03:23:09
