
不同红椿家系苗期及幼林期在江汉平原的生长表现分析
2024-01-01 00:00:00张亚东马林江樊孝萍王昌庆胡建柳张新叶
中南林业科技大学学报 2024年6期
摘 要:【目的】红椿是湖北的重要乡土树种,但在江汉平原未有天然分布及人工栽培。选择出适宜于江汉平原地区发展的优良红椿家系,为该地区提供珍贵用材树种造林资源。【方法】以湖北恩施地区的17个红椿优树半同胞家系为研究材料,采用随机区组设计,在江汉平原石首市开展平茬后2 a苗期及2 a幼林期对比试验,对不同家系连续4 a胸径(1年生时为地径)、树高和材积性状进行分析比较,开展红椿优良家系早期选择。【结果】不同家系间胸径(地径)和树高性状在所有龄期都存在极显著差异。平茬后自苗期第2年起,后续3 a的所有胸径和树高性状相互间都呈显著或极显著相关。苗期第2年起后续3 a(2020—2022年),红椿家系的胸径、树高及材积性状遗传力皆超过0.85。根据不同家系不同性状的遗传增益大小,以材积增益为主,从17个家系中筛选出3个早期表现优良的家系,即鄂13家系、鄂15家系及鄂06家系,选择强度为17.65%。【结论】3个早期优良家系材积遗传增益均超过群体平均20%,选择强度大,早期生长优势明显,年均胸径生长量2.09 cm,年均树高生长量1.78 m,这些家系将成为湖北江汉平原地区红椿良种选育的重要种质资源。
关键词:红椿;生长性状;家系选择;江汉平原
中图分类号:S722.3+3 文献标志码:A 文章编号:1673-923X(2024)06-0011-08
基金项目:“十四五”国家重点研发计划项目(2021YFD2200304)。
Analysis of growth performance of different families of Toona ciliata at seedling stage and young forest stage in Jianghan plain
ZHANG Yadong1, MA Linjiang1, FAN Xiaoping2, WANG Changqing2, HU Jianliu2, ZHANG Xinye1(1. Hubei Academy of Forestry, Wuhan 430075, Hubei, China; 2. Shishou Poplar Research Institute of Hubei Academy of Forestry, Shishou 434400, Hubei, China)
Abstract:【Objective】Toona ciliata is an important local species in Hubei province, but it has not been naturally distributed and artificially cultivated in the Jianghan plain. In order to select superior families of T. ciliata which are suitable for the Jianghan plain area, and to provide valuable timber tree species afforestation resources for this area.【Method】A comparative afforestation experiment was carried out in Shishou city, which was located in the Jianghan plain, using a randomized block design for 17 T. ciliata superior halfsib families from Enshi area in Hubei province to compare the 2-year seedling stage and 2-year young forest stage, and to carry out the early selection of superior families by analyzing and comparing the characteristics of DBH (the ground diameter was measured in the first year), tree height and volume of different families for 4 consecutive years.【Result】There were significant differences in DBH(ground diameter) and tree height traits among different families at all ages. All DBH and tree height traits in the following 3 years were significantly correlated with each other from the second year of seedling stage. The family heritability of DBH, tree height and volume traits exceeded 0.85 from 2020 to 2022. According to the genetic gain of different traits of different families, mainly by volume gain, 3 families with excellent early performance were selected from 17 families, namely E13, E15 and E06 families, with the selection intensity 17.65%.【Conclusion】3 families with excellent early performance were selected from 17 T. ciliata superior half-sib families, whose volume genetic gain exceeded 20% of the population average, with high selection intensity and obvious early growth advantage. The average annual DBH was 2.09 cm and annual tree height was 1.78 m. These families will become important germplasm resources for breeding of improved varieties of T. ciliata in Jianghan plain, Hubei province.
Keywords: Toona ciliata; growth traits; family selection; Jianghan plain
红椿Toona ciliata为楝科Meliaceae香椿属Toona落叶大乔木,是热带、亚热带地区的珍贵速生用材树种,有“中国桃花心木”之美誉,具有很高的经济价值和开发前景[1]。由于开发过度、自然环境变化和天然更新缓慢等原因,红椿分布区逐渐缩小,零星分布于我国南方多个省区,《中国植物红皮书》将红椿列为国家II级保护濒危物种[2]。近年来,红椿研究与开发越来越受到大家的关注,在资源保护[3]、种群结构[4]、苗木繁育[5]、光合特性[6]、生理生态[7]等多方面都有学者进行了研究,其中对红椿的良种选育和遗传改良方面的研究尚处于苗期测定及早期选择的初步阶段[8-12]。
红椿是湖北省的重要乡土树种,主要天然分布在鄂西地区,在咸宁九宫山等地也有零星分布,但在江汉平原地区没有天然分布,在2016年之前也没有开展过引种栽培。江汉平原是由长江、汉水及湖泊泛滥淤积而成的冲积平原,土层深厚肥沃、质地疏松、理化性能较好,是南方型黑杨种植面积最大的原料林建设基地。随着多年单一树种的大面积栽培,出现材种结构单一、病虫害严重、产品附加值低和林农积极性不高等问题,急需引进并推广一些新型珍贵速生用材树种来促进当地林业产业发展[13]。因此,为了引种并选育出适宜江汉平原发展的优良珍贵用材红椿资源,湖北省林科院近年来广泛收集省内及省外红椿资源,开展育苗及栽培研究。本研究对江汉平原引种栽培的一批红椿半同胞家系连续4 a的生长量指标进行遗传分析,对表现优良的家系进行早期评价选择,以期为江汉平原引种红椿资源提供早期适应性评价和相关试验数据,为进一步在江汉平原选育红椿优良品种奠定基础。
1 试验地概况
红椿不同家系播种苗培育及造林地点均位于湖北省石首市东升镇(112°25′16″E,29°40′25″N),海拔45~50 m。播种苗在南碾村苗圃进行培育,造林地在红椿造林前为杨树林地,皆伐翻耕后进行红椿造林。该地区属典型的亚热带季风气候地区,具有阳光充足、热量丰富、无霜期长和降水充沛等特点。年平均无霜期285 d,年平均气温16.4 ℃,年平均日照时数1 844.3 h,年平均降水量1 322.6 mm。土壤为沙壤潮土,pH值7.8,土壤颗粒均匀、质地疏松,理化性能良好。
2 材料与方法
2.1 试验材料
参试材料为17个红椿半同胞家系,优树种子采自湖北恩施地区的不同县市。具体采种优树信息如表1所示。


2.2 试验方法
2017年秋,采集恩施地区的25株红椿优树种子,于2018年春催芽后在大田搭小拱棚播种育苗。由于不同家系种子发芽率不同,成苗后密度不一,当年苗木平均地径约9.9 mm,平均株高24.8 cm。2019年3月,将25个家系中保存株数超过100株且生长较健壮的17个家系的苗木全部按40 cm×100 cm的密度,平茬后按随机区组设计重新定植培育在同一块苗圃大田中,在苗圃地连续培育2 a后利用大苗进行造林。造林苗木为裸根苗,保留5~6根长度约15~20 cm的粗壮主根及侧根,剪除长根及多余根系,造林时将苗木根部打泥浆,泥浆中施生根剂,栽入50 cm×50 cm×50 cm的穴中,按8株小区、3次重复进行随机区组设计,密度4 m×4 m。造林后每年除杂除草2次,未施肥未套种。
每年冬季苗木落叶后进行生长量调查,2019年底调查苗木地径和树高生长量,2020—2022年分别调查苗木的胸径和树高生长量。
2.3 数据分析


3 结果与分析
3.1 不同家系苗期生长分析
2019年春,从2018年播种育苗的25个家系中选出17个表现较好的家系,苗木重新平茬后再培育,栽植密度40 cm×100 cm。为培育无节大苗,在苗圃中连续培养2 a。表2列出了平茬后2 a不同家系苗的生长状况,从中可以看出,平茬后1年生17个红椿家系的平均地径和株高分别为2.52 cm和1.05 m,但第2年所有红椿家系的平均胸径(非地径)和株高分别达到3.83 cm和4.32 m,径生长和高生长都增长迅速,体现出了红椿的速生性。2019年末17个家系的平均地径生长区间为2.10~3.11 cm,平均地径最大的为鄂09家系,最小的为鄂06家系,最大家系平均地径是最小家系的1.48倍;平均株高生长区间为0.79~1.40 m,平均株高最高的仍为鄂09家系,最低的为鄂04家系,最高家系平均株高是最低家系的1.77倍。2020年末,17个家系平均胸径生长区间变化为3.05~4.46 cm,平均胸径最大的家系变为鄂17家系,最小的变为鄂07家系,最大家系平均胸径是最小家系的1.46倍,与2019年末最大最小家系的相差倍数相近;2020年末17个家系平均株高生长区间为3.62~4.83 m,平均株高最高的变为鄂16家系,最低的变为鄂07家系,最高家系平均株高是最低家系的1.33倍,比2019年最高最低家系间相差倍数明显降低。从2年的生长数据对比可以发现,平茬后第1年和第2年苗期生长表现最佳和最差的家系都发生了变化,不同家系在不同时期的生长速度处于不稳定状态。
对不同家系苗期1~2年生的地径、胸径和株高分别进行方差分析(表3),从表3可以看出,不同家系间无论是苗期第1年还是第2年,地径、胸径和株高性状的差异均达到了极显著水平,进一步对17个家系苗期性状进行多重比较(表2)。根据第1年的地径性状,生长前5的家系为鄂09、鄂17、鄂25、鄂22和鄂12家系,第2年胸径生长表现前5的家系为鄂15、鄂09、鄂16、鄂13和鄂12家系,可以发现连续2 a径生长较好且较稳定的家系是鄂09和鄂12家系。同样,对于株高性状,第1年表现前5的家系为鄂09、鄂15、鄂24、鄂17和鄂12家系,第2年高生长前5的家系为鄂16、鄂09、鄂12、鄂14和鄂15家系,连续2 a高生长较好且稳定的有3个家系,即鄂09、鄂12和鄂15家系。综合径生长和高生长2个指标,苗期连续2 a表现稳定且生长速度较快的家系为鄂09家系和鄂12家系。

3.2 不同家系幼林期生长分析
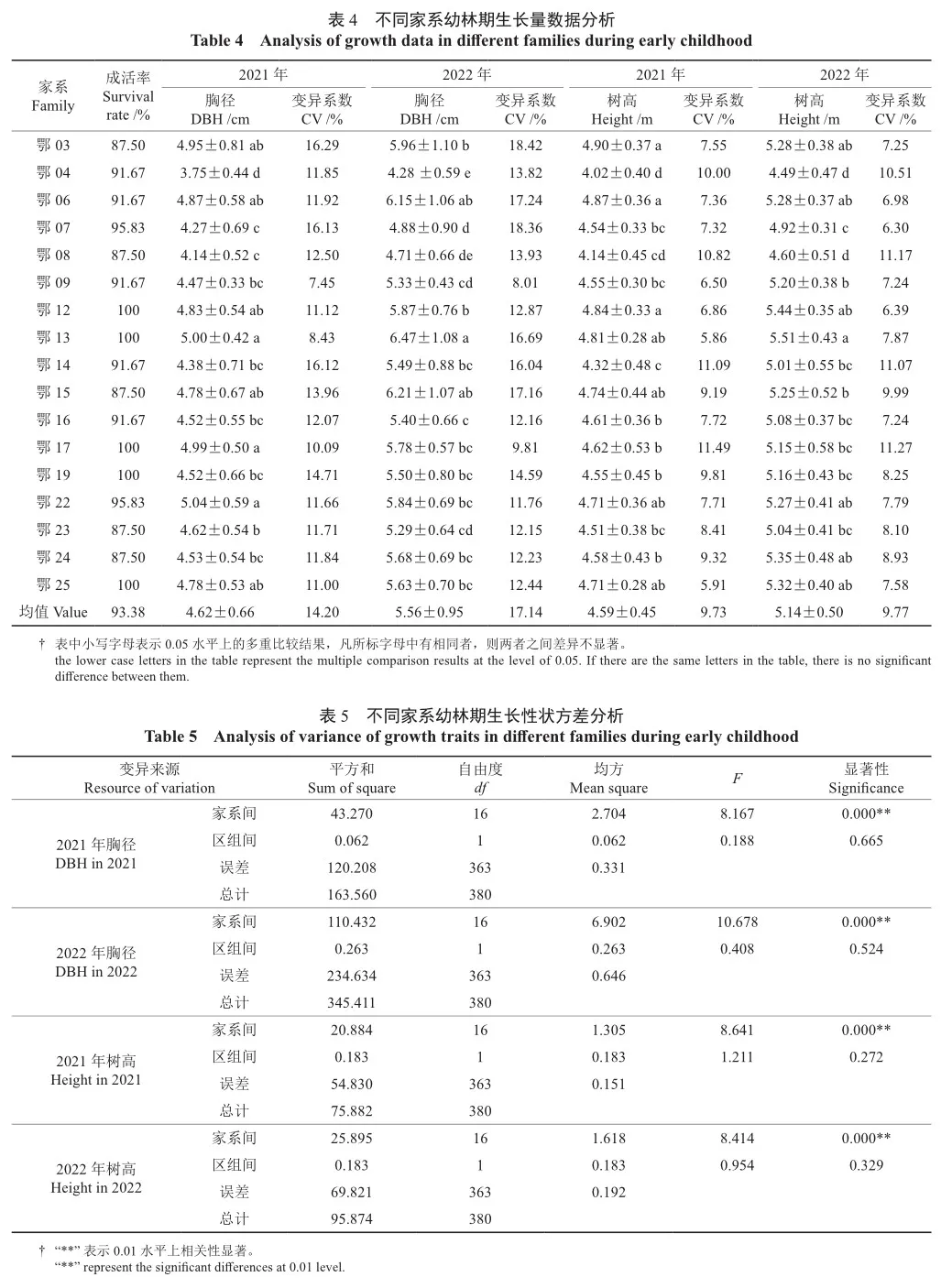
2021年春,利用红椿不同家系2年生大苗进行大田造林,分别于2021年和2022年冬落叶后进行生长量调查。表4为造林后2 a不同家系苗的生长性状分析结果。每个家系造林前皆为24株,从表4可以看出,造林后不同家系苗木的成活率有所不同,最高可以达到100%,最低为87.5%。造林第1年,所有家系苗平均胸径和树高分别为4.62 cm和4.59 m,不同家系平均胸径为3.75~5.04 cm,平均树高为4.02~4.90 m,平均胸径最大的家系为鄂22家系,最小的为鄂04家系,平均树高最大的为鄂03家系,最小的仍为鄂04家系。造林第2年,所有家系平均胸径为5.56 cm,其中最大平均胸径是鄂13家系,达到6.47 cm,最小是鄂04家系,平均胸径仅4.28 cm,两者相差1.51倍。造林第2年所有家系平均树高为5.14 m,最大树高家系为鄂13,最小树高家系为鄂04。
利用2年生大苗进行造林,对不同家系造林后 1~2年生的胸径和树高性状分别进行方差分析(表5),从表5可以看出,不同家系间无论是造林后第1年还是第2年,其胸径和树高性状的差异均达到了极显著水平,表明幼林期不同家系生长性状间也存在显著差异,根据其差异从中选择幼林期生长优良的家系是也是完全可行的。对17个家系的2 a幼林期生长性状进行多重比较(表4),结果表明,造林后第1年胸径最大的5个家系为鄂22、鄂13、鄂17、鄂03和鄂06家系,造林后第2年胸径最大的5个家系为鄂13、鄂15、鄂06、鄂03和鄂12家系,连续2 a胸径表现稳定优良的家系有鄂13家系、鄂06和鄂03家系;对于树高性状,造林第1年树高最大的5个家系为鄂03、鄂06、鄂12、鄂13和鄂15家系,第2年树高最大的6个家系为鄂13、鄂12、鄂24、鄂25、鄂03和鄂06家系,连续2 a树高表现稳定优良的家系有鄂13、鄂12、鄂03和鄂06家系。
3.3 不同家系苗期与幼林期联合选择
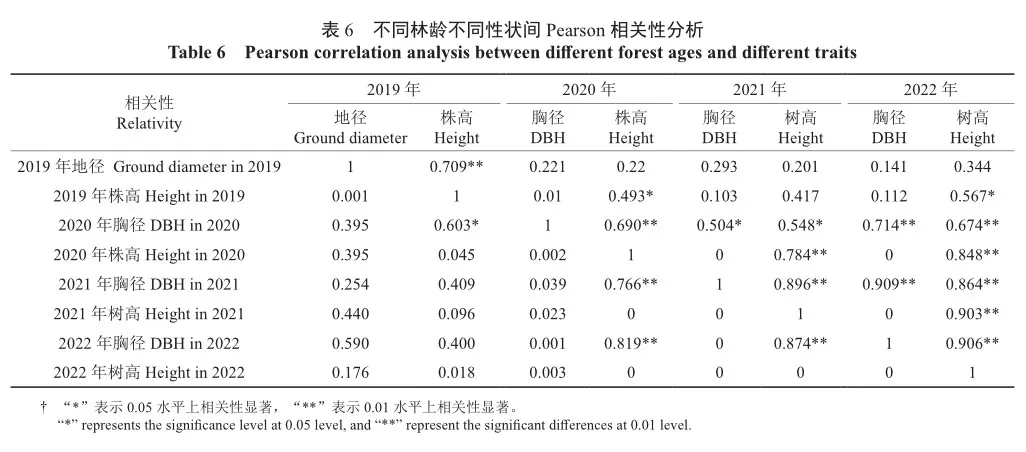
为了解平茬后2 a苗期生长性状与造林后2 a幼林期生长性状之间的相关性,对所有家系连续4 a的生长性状进行Pearson相关性分析,结果如表6所示。表中对角线上半部分为不同性状间Pearson相关性系数,对角线下半部分为不同性状间显著性参数。从表6数据可以看出,平茬后第1年苗期地径只与当年的株高呈极显著相关,而与苗期第2年及造林后2 a的胸径和树高都不显著相关。但自苗期第2年起,后续3 a的所有胸径和树高性状相互间都呈显著或极显著相关,尤其是造林后2 a的胸径和树高性状的相关系数皆超过0.85,从而表明对优良家系的早期选择可以综合考虑苗期第2年及造林后2 a的生长数据,苗期第1年的数据由于与后期数据不显著相关可以不予考虑。
基于苗期与幼林期生长数据的相关性,仅对性状显著相关的苗期第2年的生长性状和造林后2 a内的生长性状进行遗传力和遗传增益分析(表7)。从表7数据可以看出,2020—2022年胸径、树高及材积性状的家系遗传力皆超过0.85,由于是平原地区同一立地环境影响较小。根据不同家系不同性状的遗传增益大小,以材积增益为主,以胸径和树高增益为辅,综合3 a的性状变化情况,从17个家系中筛选出3个优良家系,即鄂13、鄂15及鄂06家系,这3个家系2022年材积遗传增益均超过20%,且鄂13家系在连续3 a材积增长中都名列前茅,鄂15家系在2020年材积增长中排名第1位,鄂06家系连续3 a材积增益持续增加。从17个家系中选择出3个早期生长优良的家系,选择强度为17.65%。
4 结论与讨论
4.1 结 论
本研究在江汉平原地区利用17个红椿家系1年生平茬苗开展了连续2 a的苗期生长观察,并利用2年生大苗在该地区进行造林2 a的生长性状测定。结果表明,不同家系间胸径、地径和树高性状在所有不同苗龄和林龄时都存在极显著差异。经测定,平茬后1年生17个红椿家系的平均地径和株高分别为2.52 cm和1.05 m,但第2年所有红椿家系的径生长和高生长都表现出快速增长的趋势,平均胸径(非地径)和株高分别达到3.83 cm和4.32 m。在实施造林后,第1年末所有家系平均胸径和树高分别为4.62 cm和4.59 m,第2年末所有家系平均胸径为5.56 cm,平均树高为5.14 m。对比造林前1年所有家系的平均胸径及树高,造林1 a后分别增长了0.79 cm和0.27 m,造林2 a后相比造林1 a后,平均胸径和平均树高分别增加0.94 cm和0.55 m。对平茬后2 a苗期生长性状与造林后2 a生长性状进行Pearson相关性分析,发现自苗期第2年起,后续3 a的所有胸径和树高性状相互间都呈显著或极显著相关。
4.2 讨 论
邹高顺[15]在福建来舟林业试验场连续11 a对红椿的引种栽培观测结果表明,红椿试验林的胸径和树高速生期在2~5 a。韦绪等[16]对红椿人工林生长规律的研究中指出,红椿人工林胸径年平均生长量曲线波动幅度较小,年均生长量为1.367 cm左右,红椿树高年均生长量为0.8~1.6 m,但总体呈下降趋势。本研究中,从表面上看,造林后苗木生长速度低于苗期第2年的生长速度,没有体现出红椿早期生长的速生性,这主要是由于造林采用2年生大苗,造林第1年苗木根系受损,造林后存在一定的恢复期,当年生长量有限;造林第2年胸径和树高依然没有快速生长,是由于自2022年7月起,江汉平原遭受极端高温干旱天气,伏秋冬连旱综合强度达到历史最强,致使2022年下半年苗木几乎停止生长。张拥兵等[17]在研究不同干旱程度对枣树的影响中表明,重度干旱导致的水分受限和根非结构性碳含量降低会影响到枣树水碳平衡,同时重度干旱对细根和叶这些易再生部位的生长影响较大,影响到生长和之后的存活。如果将苗期第2年与造林后2 a共连续3 a的苗木生长量进行平均来看,在江汉平原地区,17个红椿半同胞家系苗的胸径年平均生长量为1.85 cm,树高年平均生长量为1.71 m,完全符合韦绪等[16]提出的红椿人工林生长规律,甚至红椿在江汉平原的生长速度高于广西国有三门江林场的红椿生长速度。

根据后续3 a不同家系不同性状的遗传增益大小,以材积增益为主,从17个家系中筛选出3个优良家系,其造林第2年的材积遗传增益均超过20%,年平均胸径生长量达到2.09 cm,年平均树高生长量达到1.78 m。在陈娟等[12]对川中丘陵区的红椿与毛红椿半同胞家系早期选择研究中,根据造林3 a的生长数据从18个半同胞家系中选出3个优良家系,但其选择出的3个优良家系的平均单株材积遗传增益达到113.62%,远高出本研究选出的3个优良家系的材积遗传增益。主要原因在于陈娟等的试验材料来自四川、江西、安徽及湖南4个省的8个地区,且包括红椿和毛红椿2个种,从8个不同种源间进行选择优良家系选择肯定会取得较高的遗传增益,这跟张亚东等[18]利用4个不同红椿种源在武汉地区的生长差异分析取得结果相似,优良种源LZ在6 a时的平均材积可以达到较差种源JG材积的6.14倍,优良种源的遗传增益很高。本研究的试验材料全部来自湖北恩施地区,且经过1 a育苗期从25个家系中筛选保留了17个较优良的家系进行二次筛选,因此从遗传差异相对较小的群体中筛选出的优良家系的遗传增益肯定低于遗传差异大的群体。
林木早期选择的首要问题就是时间问题,即利用何种方法、采取何种指标、如何缩短长周期生长林木的合理选择时间。陈益泰[19]曾对林木的早期选择作过较为全面的论述,认为早期选择在林木育种中是十分必要的,但针对具体树种的早期选择还需根据选择的方法、选择的性状等进行多因素分析。在许秀环等[20]和张亚东等[18]开展的相同红椿材料研究中发现优良种源苗期的选择结果与6年生时的选择结果发生了很大变化;在吴际友等[10]和陈艺等[11]的相同红椿半同胞家系材料选择研究中,也发现5年生和8年生时优良家系的选择结果发生了变化。这些都说明无论是红椿种源还是红椿家系在早期生长性状都不稳定,只有通过苗期及林期的长期观测才能真实反映出不同种源或家系在某地的生长适应性及生长潜力。因此,本研究利用4 a的生长数据选择出3个优良家系,仅代表其在江汉平原地区早期生长优良,并不能代表其中、后期生长也一定优良,还需通过长时间的继续观测分析才能正确选择出真正适合江汉平原发展的红椿优良家系。下一步将持续加大红椿不同种源及家系在江汉平原地区的选择研究,进一步借鉴用材林及经济林遗传改良与品种创制技术[21],为江汉平原红椿优良品种选育工作提供技术支撑。
参考文献:
[1] 郑万钧.中国树木志[M].北京:中国林业出版社,1983. ZHENG W J. Flora of China[M]. Beijing: China Forestry Publishing House,1983.
[2] 傅立国.中国植物红皮书[M].北京:科学出版社,1991. FU L G. Chinese plant red book[M]. Beijing: Science Press,1991.
[3] 方元平,刘胜祥.湖北星斗山自然保护区国家重点保护野生植物调查研究[J].亚热带植物科学,2005,34(3):48-52. FANG Y P, LIU S X. Research on the national protected plants in Xingdoushan nature reserve in Hubei province[J]. Subtropical Plant Science,2005,34(3):48-52.
[4] 汪洋,冷艳芝,苏长江,等.恩施天然红椿种群结构及空间分布格局[J].浙江农林大学学报,2016,33(1):17-25. WANG Y, LENG Y Z, SU C J, et al. Spatial structure and distribution pattern of natural Toona ciliata populations in the Enshi region[J]. Journal of Zhejiang A F University,2016,33(1):17-25.
[5] 兰健花,冯立新.珍贵用材树种红椿扦插繁殖技术研究[J].广西农学报,2022,37(1):32-36,48. LAN J H, FENG L X. Study on cutting propagation technology of rare timber tree species Toona ciliata Roem[J]. Journal of Guangxi Agriculture,2022,37(1):32-36,48.
[6] 黄国伟,彭婵,陈慧玲,等.不同种源红椿苗期生长及光合特性比较[J].西北林学院学报,2017,32(2):123-129. HUANG G W, PENG C, CHEN H L, et al. Comparison of the growth and photosynthetic characteristics of Toona ciliata seedling from different provenances[J]. Journal of Northwest Forestry University,2017,32(2):123-129.
[7] 汪洋,冉勇军,冷艳芝,等.红椿和毛红椿遗传与生理生态相关研究[J].安徽农业科学,2014,42(20):6910-6912. WANG Y, RAN Y J, LENG Y Z, et al. Relevant study of genetic and physiological of Toona ciliata Roem and Toona ciliata var. pubescens[J]. Journal of Anhui Agriculture Science,2014,42(20):6910-6912.
[8] 文卫华,吴际友,陈明皋,等.红椿优树子代苗期生长表现[J].中国农学通报,2012,28(34):36-39. WEN W H, WU J Y, CHEN M G, et al. Seedling growth performance of Toona ciliate elite trees progeny[J]. Chinese Agricultural Science Bulletin,2012,28(34):36-39.
[9] 李艳,吴际友,邓小梅.红椿种源试验林早期生长表现[J].湖南林业科技,2015,42(5):50-54. LI Y, WU J Y, DENG X M, et al. Early growth test of different Toona ciliate provenances[J]. Hunan Forestry Science Technology,2015,42(5):50-54.
[10] 吴际友,李艳,李志辉,等.红椿半同胞家系生长与早期选择[J].中南林业科技大学学报,2016,36(4):1-4. WU J Y, LI Y, LI Z H, et al. Growth and early selection of half-sib families of Toona ciliate[J]. Journal of Central South University of Forestry Technology,2016,36(4):1-4.
[11] 陈艺,吴际友,程勇,等.红椿半同胞家系生长性状遗传分析[J].中南林业科技大学学报,2018,38(3):40-43. CHEN Y, WU J Y, CHENG Y, et al. Genetic analysis of the growth traits of half-sib families of Toona ciliate[J]. Journal of Central South University of Forestry Technology,2018,38(3):40-43.
[12] 陈娟,彭建,肖兴翠,等.红椿与毛红椿半同胞家系早期选择研究[J].中南林业科技大学学报,2020,40(8):17-24. CHEN J, PENG J, XIAO X C, et al. Early selection study of half sib families of Toona ciliata var. ciliata Toona ciliata var. pubescens[J]. Journal of Central South University of Forestry Technology,2020,40(8):17-24.
[13] 张新叶,李振芳,张亚东,等.湖北引种滇楸优良无性系早期选择[J].中国农学通报,2016,32(10):24-29. ZHANG X Y, LI Z F, ZHANG Y D, et al. Early stage selection of fine clones of Catalpa fargesii f. duclouxii in Hubei province[J]. Chinese Agricultural Science Bulletin,2016,32(10):24-29.
[14] 陈晓阳,沈熙环.林木育种学[M].北京:高等教育出版社,2005. CHEN X Y, SHEN X H. Tree breeding[M]. Beijing: Higher Education Press,2005.
[15] 邹高顺.珍贵速生树种红椿与毛红椿引种栽培研究[J].福建林学院学报,1994,14(3):271-276. ZOU G S. Study on introduction and cultivation of precious fast growing species Toona ciliata Roem Toona ciliata var. pubescens[J]. Journal of Fujian College of Forestry,1994,14(3): 271-276.
[16] 韦绪,覃德文.红椿人工林生长规律、生物量及生产力研究[J].福建林业科技,2016,43(4):38-42. WEI X, QIN D W. Studies on the growth, biomass and productive forces of Toona ciliata plantation[J]. Journal of Fujian Forestry Science and Technology,2016,43(4):38-42.
[17] 张拥兵,王林,薛浩,等.不同干旱程度对枣树苗木水碳平衡和生物量分配的影响[J].经济林研究,2022,40(3):208-215. ZHANG Y B, WANG L, XUE H , et al. Effects of different degrees of drought on water-carbon balance and biomass allocation of jujube seedlings[J]. Non-wood Forest Research,2022,40(3): 208-215.
[18] 张亚东,周国清,马林江,等.4个红椿地理种源生长差异分析[J].湖北林业科技,2022,51(2):6-12. ZHANG Y D, ZHOU G Q, MA L J, et al. Growth difference analysis of 4 different provenances of Toona ciliata in Wuhan[J]. Hubei Forestry Science and Technology,2022,51(2):6-12.
[19] 陈益泰.林木早期选择研究进展[J].林业科学研究,1994,7(增刊1):13-22. CHEN Y T. Research Progress on early selection of forest trees[J]. Forest Research,1994,7(Suppl.1):13-22.
[20] 许秀环,彭婵,张亚东,等.不同红椿种源在武汉地区的苗期生长性状比较[J].中国农学通报,2017,33(4):54-58. XU X H, PENG C, ZHANG Y D, et al. Seedling growing traits of different provenances of Toona ciliata in Wuhan[J]. Chinese Agricultural Science Bulletin,2017,33(4):54-58.
[21] 李建安,韩志强.经济林遗传改良与品种创新研究进展[J].经济林研究,2023,41(1):1-13. LI J A, HAN Z Q. Advances in genetic improvement of important traits and variety innovation of non-wood forest[J]. Non-wood Forest Research,2023,41(1):1-13.
[本文编校:吴 彬]
猜你喜欢
东北林业大学学报(2022年7期)2022-08-04 01:48:02
铁道建筑技术(2021年4期)2021-07-21 05:33:12
古今农业(2019年1期)2019-05-11 09:17:30
乡村科技(2018年17期)2018-08-16 02:27:50
湖北林业科技(2016年5期)2016-12-02 10:23:28
时代农机(2016年6期)2016-12-01 04:07:44
湖北农业科学(2015年17期)2015-10-09 22:26:04
安徽文学(2015年8期)2015-09-10 07:22:44
安徽农学通报(2015年15期)2015-08-19 01:14:09
安徽农学通报(2015年12期)2015-07-09 11:51:22
