
步氏巨猿研究九十年的回顾与展望
2024-01-01 00:00:00张颖奇
人类学学报 2024年6期
关键词:步氏巨猿;人科;神秘古猿;范式转换;上新世间断
1 前言
作为体型最大的灵长类,步氏巨猿(Gigantopithecus blacki) 是一种可以让人产生无限遐想的神秘人科成员。从1935年被首次发现并命名至今,一直都是古人类学界关注的焦点。这种巨型类人猿的化石目前仅发现于中国南方喀斯特地区早更新世到中更新世的洞穴堆积中。令人遗憾的是,尽管几代人已经搜寻了90年,但步氏巨猿的化石材料仍局限于产自19处洞穴堆积化石地点的4件不完整的下颌和近2000颗游离牙齿。而颅骨和颅后骨骼的缺乏使得步氏巨猿确切的系统位置尚需斟酌,确切的位移行为方式(locomotor behavior)也不得而知。目前,在现有的化石材料和地点基础上,除系统发育关系之外,同仁们已经对步氏巨猿的演化趋势、体质量和体型大小、生态、食性、绝灭原因等方方面面进行了探讨。Zhang 和Harrison对步氏巨猿的研究历史进行了系统性的总结[1]。Zhang等在综合研究步氏巨猿绝灭事件时,也对其研究背景和历史进行了简要概括[2]。本文对步氏巨猿发现历史的总结截至20世纪50年代,关于步氏巨猿现有化石材料的形态学也未再赘述,仅在总结其系统发育关系时对其独有的特化形态进行了介绍[1]。步氏巨猿研究中尚待解决的问题与我国早期人类演化息息相关,如步氏巨猿的起源、“神秘古猿”究竟是人是猿,中国南方喀斯特地区的上新世化石记录空白,等等。鉴于此,作者在文末以步氏巨猿发现前后人类起源中心从亚洲逐渐转移到非洲为契机,对解决相关问题的前景进行了粗略展望。期待将来可以发现步氏巨猿的突破性化石材料,进一步破解其神秘与未知,也期待中国南方地区能有新的发现,给人属(Homo) 早期演化相关理论注入中国乃至亚洲元素。谨以此文献给北京猿人第1 头盖骨发现95周年。
2 最初的发现
化石在民间被称为“龙骨”或“龙牙”,在中国被用作中药已有两千多年的历史[3-6]。西方学者在19世纪中期首先认识到它们的科学价值并开始在中国中药铺里购买[4,7-9]。1900年,Haberer KA在中国的中药铺中为位于慕尼黑的博物馆购得一大批化石牙齿,其中包括了一颗人的上臼齿[10]。正是这颗牙齿促使瑞典地质学家J Gunnar Andersson 追根溯源发现了周口店[11]。受到这些化石的启发,时任博物馆助手的von Koenigswald 于1931年开始了以中药铺为目标的化石搜寻之旅,他最初看到的大部分都是上新世的化石组合,但也识别出了一个更新世的化石组合。这一化石组合在药铺中被认为是质量较差的“龙牙”。在马尼拉的一个中药铺中,von Koenigswald 从一批据称来自香港的龙骨中发现了第一颗猩猩的牙齿。于是,1935年他到香港试图找到更多的猩猩化石,结果发现了一颗明显属于人猿超科(Hominoidea) 成员但尺寸巨大的下第三臼齿。基于其大小和独特的形态特征,von Koenigswald 建立了一个新属新种,即Gigantopithecusblacki[12],以纪念在前一年去世的加拿大人类解剖学家步达生(Davidson Black) 及其在中国所做的与北京猿人(Sinanthropus pekinensis) 相关的开创性研究。同年,在随后的中国南方化石搜寻之旅中,von Koenigswald 在广州又获得了一颗保存非常好的上臼齿。1939年,他在香港购得一颗尚未磨耗的下第三臼齿。此外,还有之前被暂且归为步氏巨猿的一些来自其他药铺的牙齿:一颗上第一门齿、一颗上犬齿(含1938 年在爪哇中药铺获得的一颗)和两颗下第四前臼齿。二战期间,von Koenigswald被日军作为战犯关押于爪哇。在被捕之前,von Koenigswald将他收集的化石托付给他的妻子和三位朋友保管[13]。为了防止被日军没收,他将几颗步氏巨猿的牙齿装入一个牛奶瓶并埋在了朋友的院子里[14]。虽然当时这些牙齿确切的产地未知,但von Koenigswald 基于药铺中获得的信息以及化石组合的特征,认为这些牙齿化石的原产地应该是广东或广西的洞穴中[4]。由于关押造成的健康问题,von Koenigswald在二战结束之后临时移居纽约,无法继续其在东南亚的研究工作,也无法继续寻找步氏巨猿的原生层位。
3 原生层位的发现
差不多与von Koenigswald 在中药铺中搜寻步氏巨猿牙齿的同时,中国初代的古生物学者们也开始了中国南方地区的洞穴调查[15,16],寻找步氏巨猿的原生层位成为他们的重要任务。1935年,中国农矿部地质调查所新生代研究室(中国科学院古脊椎动物与古人类研究所的前身)在中国南方地区开展了一次新生代堆积的初步考察,其中就包括广东和广西的洞穴堆积。此次考察识别出了中国南方地区洞穴堆积的两个年代地层单元,即下中更新统“黄色堆积”和上更新统或更新世之后的“含贝壳文化堆积”。调查队从所谓的“黄色堆积”中发现了与著名的盐井沟剑齿象—大熊猫动物群可以对比的化石,这成了确定步氏巨猿原始产地的关键证据,因为药铺中购得的步氏巨猿牙齿都是与剑齿象—大熊猫动物群成员的牙齿掺杂在一起的。此外,步氏巨猿牙齿独特的保存状态,如齿根经常由于豪猪的啮咬而缺失及黏附的“黄色泥土”,为其可能的原始产地提供了额外的线索[4,17]。
1955年,中国科学院古脊椎动物与古人类研究所(后文简称古脊椎所)派遣由裴文中和贾兰坡带领的考察队去广西考察,试图解决步氏巨猿的产地和时代问题。为确定产出化石的洞穴的位置,考察队对广西和广东的合作社和土产出口公司进行了筛查,并获得了47颗巨猿牙齿[18]。基于这些信息,从1956 年到1960 年,考察队共调查了三百多个山洞,其中88 处含有哺乳动物化石[5,6]。1956 年,考察队发掘了大新县榄圩人民公社正隆村那隆屯牛睡山黑洞,发现了步氏巨猿的一颗下第三臼齿、一颗下第三前臼齿和一颗上臼齿以及大量的伴生哺乳动物化石[18]。其中,第一颗出土自原生层位的步氏巨猿牙齿是发掘的第一天贾兰坡在黑洞发现的[19]。根据裴文中和吴汝康的描述,黑洞中含化石的堆积物可以分为上下两层,三颗步氏巨猿的牙齿“是从胶结坚硬的黄色角砾岩层下的一层红紫色洞穴土状堆积中掘出的”,而哺乳动物化石则产自上层“坚硬的角砾岩层”[18]。这是步氏巨猿首次于原生层位中发现,从而使其产地及层位得到了确认。为了纪念这一历史性的发现,黑洞被命名为“大新巨猿洞”[20]。
1956年4月,政府禁止了“龙骨”的发掘和收购。当年夏天,柳城县凤山公社社冲村村民覃秀怀在附近的楞寨山硝岩洞意外收获了一些化石,即“龙骨”。之后拿去洛满圩合作社售卖。时任洛满人民银行行长的韦耀社认为当中一件下颌骨可能会有一定的科学价值,便说服覃秀怀将其捐献给了政府。1956年冬,这件下颌骨被送到正在广西考察的古脊椎所的科学家手上[6,21]。而该下颌骨正是他们要找的步氏巨猿,这也是世界上第一个被发现的步氏巨猿下颌骨。1957 年1 月,考察队对硝岩洞开始了系统发掘,发掘一直持续到1963年,期间一共进行了八个野外季度的发掘工作,时间从三个月到五个月不等,共出土了步氏巨猿的两件下颌骨以及一千多颗游离牙齿,硝岩洞被重新命名为“巨猿洞”。为了与先前的“大新巨猿洞”相区别,硝岩洞被称为“柳城巨猿洞”[20,22,23]。到目前为止,柳城巨猿洞仍然是出土步氏巨猿化石最多的地点。
之后的二三十年里,中国的学者又陆续发现了一些新的步氏巨猿化石地点[1],这是搜寻步氏巨猿的第一波热潮。这一阶段的野外考察和室内研究明确了步氏巨猿产自中国南方地区的洞穴堆积,增进了我们对其形态和年代的理解。其中,1965年冬发现的武鸣步氏巨猿牙齿化石值得关注。该化石是古脊椎所于武鸣县甘圩人民公社步拉利山半山腰一山洞中进行试掘时,在洞内残留的胶结坚硬的黄色砂质黏土中发现的,共12 颗步氏巨猿牙齿[24]。这一地点之所以重要,一是因为其时代更晚,二是因为其牙齿化石明显更大,指示了步氏巨猿牙齿随时间增大的趋势。1973年,von Koenigswald 受邀来到中国亲自观察这些新发现的布氏巨猿标本,并与裴文中、杨钟健和吴汝康等中国学者会面探讨[14]。
4系统发育关系
von Koenigswald 最初建立步氏巨猿这个种时,认为它与印度西瓦里克(Siwaliks)中新世晚期的人猿超科成员存在相似性,是一种特化的猿,可能代表了西瓦古猿类(以Sivapithecus 为代表)的一个旁支[12]。Broom[25] 以及Broom 和Schepers[26] 最早认为步氏巨猿是人类谱系成员,并与南方古猿属(Australopithecus) 紧密相关。Weidenreich 最初认为步氏巨猿是一种体型巨大的猿[27],但后来他认为步氏巨猿属于人族(Hominini)并且是直立人(Homo erectus) 的直接祖先[17]。von Koenigswald 同意步氏巨猿属于人族的观点,但认为其特化程度太高并且在时代分布上与当时已知的直立人化石相重叠,所以不大可能是亚洲人族成员的直系祖先[4];他更倾向于把步氏巨猿当作一种与主流人类谱系平行演化的特化人族成员,而其祖先则有可能是印度晚中新世的巨印度猿(Indopithecus" giganteus)。
在接下来的几十年当中,步氏巨猿的系统发育关系一直摇摆不定。许多学者同意Weidenreich[17] 和von Koenigswald[4] 的观点, 赞同其作为人族成员的地位[22,28-35]。Weidenreich[36] 和Weinert[28] 甚至提议将巨猿属(Gigantopithecus) 更名为巨人属(Gigantanthropus 或Giganthropus),但都属于不符合命名法则的提议。也有很多学者认为步氏巨猿是一种特化的大猿[18,37-42]。
步氏巨猿的齿颌形态高度特化,拥有一系列独有的特征将该属与所有其他的化石和现生人猿超科成员区别开来。这些特征可以总结如下:下颌骨粗大厚实,具有结实的联合部和深且粗壮的下颌体;下面部相对短且深(基于下颌骨的形态);下颌骨齿隙缺失或非常小;下颌门齿列异常狭窄并且竖直着生,齿冠低且呈钉状;上、下犬齿低冠且粗壮;下颌第三前臼齿具有双齿尖,近中颊侧面的形态更加适合与上犬齿紧密咬合;下颌第四前臼齿相对大且臼齿化;下颌臼齿窄长,宽度向远中略收缩,齿冠中部在颊舌向收缩强烈,下次小尖小;上颌臼齿向远中方向仅略微变窄,齿冠中部的颊舌向收缩也很微弱;上下颌臼齿高冠,齿尖圆钝肥硕并挤满咬合面,不甚发育的齿脊低且圆钝,咬合面的凹浅且不甚发育,齿尖之间以很深的缝隙间隔,釉质褶皱形成很多附属结节使咬合面呈复杂多尖状;臼齿拥有极厚的釉质和低矮的齿质齿尖;上下颌第三臼齿小于第二臼齿。所有这些特化都可看作是步氏巨猿的独有衍征(autapomorphy),但极厚的釉质和低矮的齿质齿尖可能是对祖先人科成员原始形态型的保留,因为这些特征在所有人猿超科冠群成员中均已丢失。然而,步氏巨猿的许多独特特征及某些形态特征组合都可以在早期人族成员中找到,这就导致了很多学者最初将步氏巨猿作为人族谱系的一个早期演化旁支或者直立人直系祖先[4,17,22,32,34]。步氏巨猿与早期人族成员如南方古猿所共有的关键形态特化包括粗壮的下颌骨、相对短且深的下面部、减小的齿隙大小、门齿竖直、犬齿低冠、下颌第三前臼齿双尖且与上颌犬齿咬合方式发生变化、颊齿相对大并具有低矮圆钝且肥硕的齿尖和齿脊、釉质厚等等。然而,尽管这些相似性看起来非常显著(尤其是与粗壮型南方古猿相比),但如果仔细考量每一个特征的详细形态,就会发现它们只是表面的相似。与其说这些特征支持与人族紧密的系统发育关系,倒不如说这些共有的特征是齿列和下面部为响应相似的结构——功能需求所作的趋同演化[40,41]。
二十世纪六十年代,关于步氏巨猿系统位置的争论发生了转折。起因是印度北部西瓦里克赫里塔良戈尔(Haritalyangar) 附近发现的一件时代为晚中新世的下颌骨。这件下颌骨被认为是巨猿谱系的一个原始成员,即比拉斯普尔巨猿(Gigantopithecusbilaspurensis)[40,41,43,44]。其发现于巴基斯坦博德瓦尔高原(Potwar Plateau) 道克派珊组(DhokPathan Formation) 地层中,推定年代为860万年前[45-47]。比拉斯普尔巨猿的牙齿比西瓦里克更早的西瓦古猿属(Sivapithecus) 中最大的种类还要大很多,其下颌骨的下颌体更深更粗壮。正如人们预期中的步氏巨猿祖先那样,比拉斯普尔巨猿的牙齿和下颌都比前者要小很多。其下颌颊齿同样比步氏巨猿更为原始,并与西瓦古猿属更为相似。与步氏巨猿的不同之处在于,比拉斯普尔巨猿的臼齿并不是很高冠,也没有繁杂多尖的咬合面形态以及在颊舌方向的强烈收缩,下颌第四前臼齿相对大但没有臼齿化,下颌第三前臼齿为一单尖齿且保留了与上颌犬齿咬合的近中颊侧珩磨面。如Simons 和Chopra所描述[43],比拉斯普尔巨猿的珩磨面相对短,与犬齿小这一特征相匹配。这与雌性大猿中的情况非常吻合,并不意味着在其他大猿中也很典型的上颌犬齿/ 下颌第三前臼齿的珩磨组合在比拉其普尔巨猿中有明显的不同。比拉斯普尔巨猿的下门齿没有保存,但是犬齿之间的距离狭窄,意味着门齿相对小且排列紧密,这一点与步氏巨猿相同。下犬齿与犬后齿相比相对显小。
此后,比拉斯普尔巨猿的命名及系统位置几经修订。Pilgrim 记述了产自巴基斯坦博德瓦尔高原哈斯诺(Hasnot) 附近一处地点一颗游离下臼齿(GSI-D175),将其命名为巨森林古猿(Dryopithecus giganteus),年代与比拉斯普尔巨猿的下颌骨相近[48]。这一颗牙齿从形态上看明显应与比拉其普尔巨猿属于同一物种。von Koenigswald 将这颗下臼齿(GSI-D175) 归入印度猿属(Indopithecus)[49],并认为其与巨猿属存在亲缘关系[4,50]。最终,比拉斯普尔巨猿被认为是巨巨猿(Gigantopithecus giganteus) 的晚出异名[42,51-53]。Cameron 后来重新启用了Indopithecus 这一属名[54]。他认为南亚的化石人猿超科成员与欧洲同时代的希腊猿属(Graecopithecus) 关系更为密切,而步氏巨猿则是亚洲包括猩猩属(Pongo)、西瓦古猿属和禄丰古猿属(Lufengpithecus) 在内的猩猩亚科这一分支的成员。大部分后来的学者都接受这些印度—巴基斯坦标本的独特性,但倾向于将它们称为巨印度猿[45,55-60],但也存在不同观点[61]。
20世纪七八十年代,随着中新世的人猿超科成员与早期人族成员的关系逐渐明确及支序系统学的兴起[42,62-66],步氏巨猿与人族的亲缘性逐渐淡化,人们开始将其看作是一种猿。比拉斯普尔巨猿(现已修订为巨印度猿)和步氏巨猿之间可能的祖裔关系成为确定后者系统发育位置最为关键的因素,而前者明显衍生自西瓦古猿属中的某个类型[40,41,43,44]。Simons曾提出巨猿属与意大利晚中新世山猿属(Oreopithecus) 之间可能存在关系[67],但是这一观点也随着后来西瓦里克地区的新发现而被研究者们摒弃[44]。
当前的共识,认为步氏巨猿是一种特化的猩猩亚科成员,可能代表了Sivapithecus—Indopithecus 这一分支中的一个末梢成员[53,59-61,68-72]。基于目前的证据,这的确是一个合理的假说,但与此同时我们还要清醒地认识到,所有关于步氏巨猿的认识都局限于牙齿和下颌骨,而这些化石材料对于解决人猿超科成员的系统发育关系存在先天不足。虽然步氏巨猿的齿颌形态具有高度的衍生性,理论上讲应该很容易在潜在的姐妹群中甄别出共有衍征(synapomorphy),但事实上建立这样的系统发育关系并非易事。例如,几乎没有能够将巨猿属和猩猩属联系在一起的确凿的共有衍生形态特征支持其属于猩猩亚科分支。釉质极厚的臼齿及低矮的齿质齿尖、浅的齿凹、平缓的咀嚼面起伏是为数不多的唯独由步氏巨猿与猩猩属所共有的特征[73]。然而,这些特征组合也可能出现于中中新世欧亚和东非的人科干群成员(stem hominids),如谜猿属(Griphopithecus) 和肯尼亚猿属(Kenyapithecus),可能代表了原始的人科成员形态[59,60,74]。
因此,目前巨猿属之所以被归入猩猩亚科,是因为其与印度猿属的祖裔关系而间接建立的与西瓦古猿属(一个有颅骨特征支持的确切的猩猩亚科成员)之间的联系。然而,如Wood 和Harrison[75] 以及Begun[60] 所述,步氏巨猿的典型特征(退化的前齿,前臼齿臼齿化,颊齿相对尺寸增大)在其他化石人猿超科成员尤其是在中新世晚期的类群中也发生了独立的演化。如果趋同特征(homoplasy) 也被混在一起考虑,加上可用于系统发育关系重建的特征范围仅限于牙齿和下颌,以及步氏巨猿齿列中的独有衍征,支持步氏巨猿与任何中新世的人猿超科成员或者猩猩属亲缘关系的证据都是薄弱的。然而,如前所述,巨猿属和印度猿属之间的系统发育关系的确是最为合理的解释,我们暂且可以将步氏巨猿看作猩猩亚科分支中于晚中新世起源于亚洲的支系中的末梢成员。尽管Welker 等从吹风洞的一颗步氏巨猿牙齿釉质中成功提取了古蛋白质组,序列对比表明步氏巨猿在现生大猿中与猩猩互为姐妹群[76],但考虑到序列片段的高度碎片化以及对比样本仅有现生种类,并不能反映相关类群形态演化的历史,且不能排除蛋白质组序列存在趋同的可能性,今后在探讨步氏巨猿的系统位置时,上述的问题仍然不应该忽视。
5 国外疑似步氏巨猿化石的甄别
20世纪60年代,越南学者们在越南北部靠近中越边界的地方发现了很多洞穴地点[77-80]。Tham Khuyen 洞是其中最为重要的洞穴之一,先前的研究表明直立人曾在此处与步氏巨猿共生,其年代约为47.5 万年前[81]。但是这里出土的步氏巨猿标本,包括一件左下犬齿(TK65/122) 以及一件带疑问的下门齿(TK65/146) 被重新鉴定为猩猩[1]。Lopatin 等报导了产自越南北部Thanh Hoa 省Lang Trang 洞的两颗晚更新世的步氏巨猿牙齿,包括一颗完整且带齿根的右下第二臼齿(PIN no. 5644/439) 及一颗左下第二臼齿的远中半部齿冠(PIN no. 5792/490)[82]。根据其研究可知,这两件标本齿冠低、冠面起伏平缓、各齿尖靠近齿冠边缘、釉质褶皱细密、齿带明显、下跟凹宽广,明显区别于步氏巨猿的臼齿形态特征,应归为魏氏猩猩(Pongo weidenreichi)。
Noerwidi 等报导了产自爪哇Semedo地点的两件步氏巨猿化石[83]。Semedo 3417为一段左侧下颌体残段带第一臼齿齿冠基部以下部分和第二、第三臼齿,下颌体臼齿列前后部分均未保存。Semedo 3418为两段下颌体残段被挤压错位并由堆积物胶结在一起的标本,左侧残段带第二臼齿,右侧残段带第一和第二臼齿。从其外部特征判断,这两件标本确属步氏巨猿无疑。根据Noerwidi 等的叙述,这两件标本由一位名为Dakri 的当地人分别于2014 年6 月和8 月采集于Semedo 地点4 km2 范围内的地表,具体的位置和层位不详,这名当地人还于2011 年在同一地区以同样的方式采集到一件直立人的颅骨残片[83]。考虑到von Koenigswald提及爪哇的中药铺中也普遍存在产自中国南方喀斯特地区洞穴堆积中的龙骨[12],作者认为这两件标本的真实来源十分可疑。在这两件标本的真实产地和层位水落石出之前,我们暂时对其作存疑处理。
Bocherens 等报导了产自泰国中更新世地点Pha Bong 的一件巨猿右下第四前臼齿[84]。但该标本齿冠偏低、冠面细微釉质褶皱明显,当属魏氏猩猩无疑。因此,目前步氏巨猿的可靠分布仅限于中国境内。
6地理和时代分布
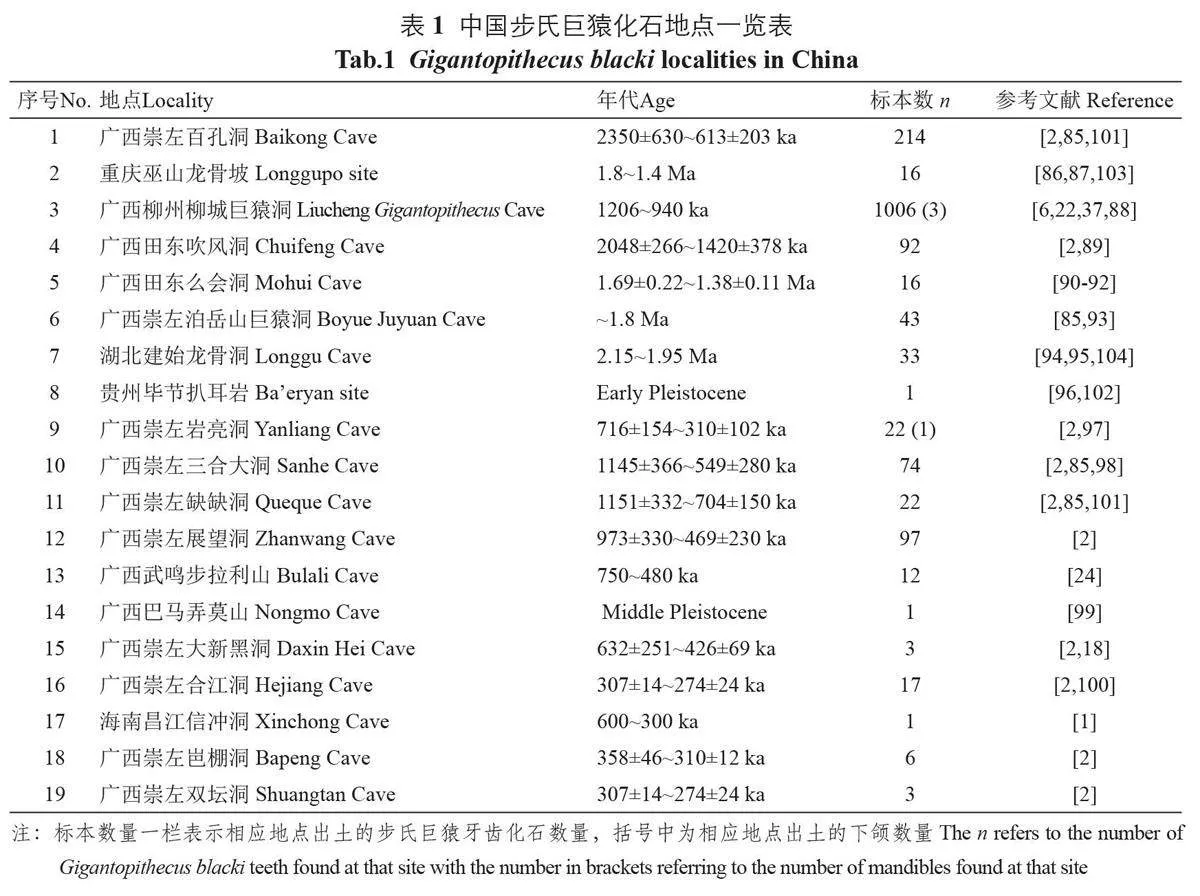
对步氏巨猿的搜寻一直持续至今。截至目前,已公开发表的步氏巨猿化石地点共有19 处(表1)。除Zhang 和Harrison 总结的17 处[1] 之外,还有Zhang等研究步氏巨猿绝灭事件时涉及的岜棚洞和展望洞[2]。在这19 处地点当中,有15 处在广西,这其中又有9处在崇左。新的化石材料和地点随着调查的继续还会不断增加。
在中国,步氏巨猿的化石地点广泛分布于长江和海南岛之间、云贵高原以东的中国南方喀斯特地区[85-100](表1)。已经发现步氏巨猿化石地点的省市包括重庆、湖北、贵州东部、广西和海南,然而这些地点当中的绝大多数都分布于广西境内,似乎表明该地区为步氏巨猿一个重要的演化和分布中心[101]。最靠北的步氏巨猿化石地点为位于长江南岸的湖北建始龙骨洞和重庆巫山龙骨坡,而最靠南的地点位于海南省昌江黎族自治县。但步氏巨猿的地理分布随时间推移也发生了显著的变化。早更新世步氏巨猿分布范围更广,可以延伸到更靠北的贵州高原[102] 和长江南岸地区[103,104]。或许是为了逃避北方分布区域变得干冷的气候,中更新世的步氏巨猿则退缩到更靠南也更加温暖和湿润的广西和海南[101,105]。
从目前的化石记录来看,步氏巨猿的时代分布为早更新世至中更新晚期。Zhang 等从区域性的视角出发,以广西境内22处洞穴堆积化石地点为研究对象,基于157个放射性测量年代结果,将步氏巨猿的生存年代范围锁定在230 万~25.5万年前[2]。最古老的步氏巨猿化石地点为崇左百孔洞,其年龄最老可以达到230万年。时代最新的步氏巨猿化石地点为25.5 万年的崇左双坛洞。而步氏巨猿最终绝灭的窗口期为29.5万~21.5万年前。
7体质量和体型大小
颅后骨骼尤其是四肢骨骼化石材料的缺乏,致使推测步氏巨猿的体质量和体型大小非常困难。根据步氏巨猿颊齿硕大的尺寸以及下颌骨的粗壮程度,可以推断其应该比任何现生人猿超科成员都要大。与体型最大的现生大猿西部大猩猩(Gorilla gorilla) 相比,步氏巨猿上下臼齿的平均面积分别达57.8% 和33.0%。步氏巨猿臼齿的大小超过了所有亚洲中新世的人猿超科成员,但是南亚晚中新世的巨印度猿和安纳托利亚(Anatolia) 中部晚中新世的土耳其天猿(Ouranopithecus turkae),接近步氏巨猿的整体大小[1]。Weidenreich 算出步氏巨猿臼齿的尺寸为大猩猩的两倍[17]。按照雄性大猩猩的平均体质量为169 kg[106] 来计算,步氏巨猿则应为338 kg。还有学者提出225~500 kg 的估算体质量[11,41,44,72,107] 以及2.7~3.7 m 的站立高度[6,11,44]。根据下第一臼齿与体质量的线性相关函数计算出来的体质量为204 kg[108]和280 kg[109]。很显然,使用牙齿大小作为化石灵长类体质量估算依据存在一定的问题,因为牙齿大小受功能和个体发育因素影响,在任何体型的灵长类中都存在很大的差异[110]。但是,根据已知的解剖特征,步氏巨猿可能是一种相对的硕齿型(megadont) 人猿超科成员。因此,任何基于臼齿尺寸的体质量估算都可能存在高估的可能性。没有颅后骨骼材料,很难给出一个步氏巨猿的可靠的体质量估算,但是200~300kg应该算是一个可以接受的参考。Ciochon 等基于步氏巨猿下颌的尺寸,按照猩猩颅骨和下颌的比例计算出步氏巨猿的头骨高度约为46 cm,然后以略大于现代人头身比(通常为1:7)的比例1:6.5 计算出其身高大约为3 m[11]。这样的体型基本上排除了步氏巨猿为树栖性的可能性,因此我们推测其主要生活于地表。但需要强调的是,只有找到颅后骨骼尤其是四肢骨骼,才能确切地知道其位移行为方式。
8 性别差异与社会性行为
步氏巨猿的犬齿和下颌均展现了高度的性别差异[22]。柳城巨猿洞下颌骨III 和I 下颌体尺寸的差异(下颌骨III 第一臼齿处下颌体深度比下颌骨I 大40%)表明雌性和雄性的头骨也存在显著的差异。步氏巨猿下颌体深度的性别差异超过了包括大猩猩在内的现生大猿。在大猩猩中,雄性平均下颌体深度仅比雌性大16%~20%[111]。步氏巨猿上下犬齿的大小分布存在双峰性[1]。雄性和雌性上犬齿最大径的均值分别为21.1 mm 和15.4mm,前者与后者比率为1.37。与现生狭鼻猴类(catarrhines) 比较,步氏巨猿上犬齿性别差异水平超过除大猩猩之外的所有现生人猿超科成员。关于步氏巨猿性别差异,Oxnard也得到了相似的结论[112]。总之,步氏巨猿的雄性比雌性拥有更大的体型,明显更大的犬齿,表明雄性之间的竞争相对激烈[113]。然而,由于犬齿在两性当中都相对较小,即使在雄性当中,其仅稍微伸出颊齿咀嚼面。据此可以推断,犬齿炫耀(canine display) 在步氏巨猿的竞争技能中并不是重要的组成部分。此外,正如大猩猩那样,灵长类中高度的性别差异通常与一雄多雌(polygyny)的种群性别结构有关。Oxnard 认为步氏巨猿种群中雄性和雌性的数量几乎相等,所以其性别结构可能并不是所谓的一雄多雌[112]。一项以包括黑猩猩和大猩猩在内的7 个野生灵长类群体的研究表明所有群体的雌雄比例都接近于1:1[114]。导致这一现象的原因主要在于,和大猩猩一样,独居的雄性以及全雄性群体通常会生活在一雄多雌群体的周边[115]。基于这些研究和步氏巨猿的性别差异程度,步氏巨猿种群应有与大猩猩相似的性别结构。
9 演化趋势
步氏巨猿游离牙齿标本的数量较多,加上越来越多新地点的发现以及更加可靠的年代测定,使得讨论步氏巨猿是否随时间发生过形态变化成为可能。Zhang[116] 和张银运[117]分析了其牙齿大小随时间的变化。他所使用的样本包括最初发现的5 个洞穴地点[18,22,24,94,99],以及地点未知的采购标本[22]。基于当时对步氏巨猿的年代学认识,Zhang 将样本按时代分为两组。早更新世组包括柳城巨猿洞(638颗牙齿),而中更新世组包括建始、巴马、大新和武鸣(19颗牙齿)[116]。而采购的标本均被归入中更新世组。原因在于这些标本很有可能来自洞口低、易于到达的洞穴地点,所以应该晚于早更新世。Zhang 的结论是早更新世组的颊齿比中更新组在统计学意义上要小,表明步氏巨猿的牙齿随时间有增大的趋势[116]。此后,张银运重新测量了所有牙齿以避免观察者间误差,并增加了湖北采购的180 颗牙齿到样本中[117]。年代分组与Zhang[116] 相同,但较年轻的组的年代被认为是晚早更新世、早中更新世或更晚。采购的标本也同样被归入时代较晚一组。张银运再次确认了其先前的结论,较年轻的组要比早更新世的组显著大[117]。
Zhang 等基于产自14个地点1218颗步氏巨猿犬后齿测量数据重新验证了这一问题[118]。在更加可靠的年代数据基础上,Zhang等将样本按时代早晚分为3组:早早更新世的百孔洞、龙骨坡、柳城巨猿洞、吹风洞和么会洞,晚早更新世的岩亮洞、泊岳山巨猿洞、建始龙骨洞、三合大洞和缺缺洞,以及中更新世的武鸣、巴马、大新黑洞和合江洞[118]。分析结果支持Zhang[116] 和张银运[117] 的结论,步氏巨猿的牙齿从早更新世到中更新世存在逐渐增大的趋势。
除了随时间增大之外,步氏巨猿牙齿可能还发生了形态的变化。Zhang等基于micro-CT 数据研究了步氏巨猿上前臼齿的釉质—齿质界面形态,将产自最年轻的洞穴地点之一合江洞(40万~32万年前)的三颗上前臼齿的形态与柳城巨猿洞、三合大洞和采购标本进行了对比[119]。结果表明,合江洞上前臼齿的主要齿脊比其他对比标本的发育要更锐利清晰。此外,合江洞上前臼齿上的附属齿脊,尤其是源自前尖的附属齿脊数量更多且更发育。与其他地点步氏巨猿的上前臼齿不同,合江洞上第三前臼齿的原尖前棱与前附尖脊之间没有相互连接,上第四前臼齿上与前尖前棱之间不存在连接,导致了近中缘脊出现了一个缺口。由于合江洞的年代较新,一个可能的解释就是这些形态差异反映了步氏巨猿种群由于承受生存压力而濒临绝灭之前牙齿形态发生的趋于复杂化的微演化。
10埋藏学
自从von Koenigswald 于1935年首次发现并命名步氏巨猿以来,在中国南方喀斯特地区陆续已经发现19处步氏巨猿化石地点(表1),但其化石材料仅限于4 个不完整的下颌骨[1] 和近2000颗游离牙齿。游离牙齿和下颌骨数量的丰富与颅骨和颅后骨骼的缺乏形成了鲜明的对比。遗憾的是,中国南方喀斯特地区含步氏巨猿化石的洞穴堆积的埋藏学研究还没有系统开展。仅White 对柳城巨猿洞的埋藏学进行了初步观察[120]。同样是由于出土化石中骨骼的缺乏与游离牙齿的压倒性多数形成了鲜明的对比,并且多数牙齿齿根上都存在啮齿类啮咬的痕迹,所以他认为造成中国南方地区洞穴堆积化石地点游离牙齿化石富集但缺乏骨骼的罪魁祸首是豪猪。豪猪有搜集和啮咬骨骼的习性。它们能够将骨骼搬运或拖拽到它们的巢穴中,并啃食骨骼和牙齿的齿根,而牙齿的齿冠由于有坚硬的釉质包裹则被丢弃。对现生南非豪猪的研究表明,它们每年可以搜集数十块骨骼用以啮咬[121]。这样的速度如果放到地质历史的尺度,确实可以造成牙齿化石的大量富集。豪猪也是中国南方地区更新世动物群中最为常见的成员之一[101,122]。这就解释了在中国南方洞穴地点中游离牙齿与骨骼保存数量的巨大差异。其他的哺乳动物,如食肉类,可能也对化石的富集起了一定的作用,但是它们的作用明显都只是辅助性的。这些化石牙齿上也很少能见到因滚动而造成的磨损,因此除了季节性流水的冲积之外,流水的长距离搬运因素应该可以被排除。步氏巨猿游离牙齿的丰富和颅后骨骼的缺乏也是出于同样的原因。但耐人寻味的是,如果没有豪猪在洞穴中对这些骨骼所作的破坏,没有它们作为化石富集的中介,我们可能永远也不会知道步氏巨猿的存在。如此看来,步氏巨猿的颅骨及颅后骨骼只能寄希望于将来特异埋藏的发现。好在中国南方喀斯特地区第四纪洞穴堆积非常丰富,只要分区域开展拉网式的主动性洞穴调查,步氏巨猿的突破性化石材料总有一天会重见天日。
11古生态、食性以及绝灭
目前,除了最靠南的海南信冲洞位于热带雨林带之外,步氏巨猿的其他化石地点都分布在中国南方地区的亚热带常绿阔叶林地区[123]。虽然亚热带的北界在整个更新世期间随着冰期旋回在不断变化,但可能步氏巨猿生存时期的植被和气候大体与现在并无二致。分布区内南部早更新世和中更新世洞穴地点由东洋界适应了热带和亚热带气候的动物群成员占主导,而北部的地点则更趋于东洋界和古北界成员的混合[101]。广西崇左三合大洞的小哺乳动物群中以森林为栖息环境和树栖的种类占主导地位,它们的生态偏好确认了步氏巨猿栖息于热带和亚热带森林环境[124]。更新世期间,在广西地区与步氏巨猿共生的狭鼻猴灵长类非常多样,包括猕猴属(Macaca)、仰鼻猴属(Rhinopithecus)、白臀叶猴属(Pygathrix)、乌叶猴属(Trachypithecus)、冠长臂猿属(Nomascus) 和猩猩属[85]。虽然在现代的动物群中并没有可以与之相匹配的灵长类群落,但越南北部的热带雨林环境中栖息的灵长类与之最为接近[125]。
早更新世缺缺洞巨猿化石层位中的孢粉研究结果指示了一个温暖的温带到亚热带气候,落叶和常绿阔叶林的混合环境[126]。森林由桦树、橡树和栗树为主导,并有多样化的灌木和草本植物林下层,包括蕨类、草和双子叶植物。
早更新世步氏巨猿牙釉质C、O同位素研究表明其栖息于潮湿、茂密且林冠封闭的亚热带森林中[105,127-129]。Zhang 等的孢粉和C、O 同位素研究同样表明,步氏巨猿更加偏好林冠封闭的森林环境[2]。与广东省鼎湖山自然保护区的生态环境非常匹配[128,130],其代表了一种亚热带季风常绿阔叶林,年均温为21℃,年降水量1900 mm[131]。
由于其高度特化的齿颌形态,步氏巨猿摄食行为的重建也是研究者们关注的问题。有多种方法和手段被用来进行相关研究,每一种都为步氏巨猿的食性偏好提供了重要的线索,但是难点在于将如何将多种证据整合到一个统一的功能—行为模型当中。
深且粗壮并向后逐渐加深的下颌体以及厚实的联合部都表明步氏巨猿咀嚼坚韧食物时下颌承受了强烈的应力[132-134]。遗憾的是,食物种类、物理性质以及下颌骨形态之间的关系还不清楚[134,135]。例如,步氏巨猿很深的下颌骨在现生高等灵长类中的叶食者、种子食者和硬物食者中也存在[136-139]。同样,向后逐渐加深的下颌体通常也存在于叶食性高等灵长类中,但偶尔也存在于种子食者中。
釉质厚且高冠的臼齿以及倾向于重度磨耗的颊齿,显示出牙齿为研磨食物而发生的适应[140]。臼齿化的前臼齿、相对大且高冠的臼齿以及颊齿加长的齿根说明步氏巨猿可以强有力地咀嚼大块的纤维质食物[141]。然而,起伏平缓的齿尖和发育弱的齿脊表明步氏巨猿并不适应高效地咀嚼树叶尤其是草叶。步氏巨猿犬后齿的高龋齿率[22,105,89] 与现生黑猩猩相似,表明其食物包括碳水化合物丰富的果实[142,143]。较小的门齿表明食物在摄入之前可能并不需要太多的处理,但门齿与颊齿不同的磨耗方式的确意味着门齿的重度使用。
大熊猫与步氏巨猿共生,两者均有粗壮的下颌、很厚的釉质、形态复杂的齿尖以及丘齿型臼齿,这使得一些研究者认为步氏巨猿可能也专门以竹子为食[44,51,120,141,144]。然而,步氏巨猿更厚的釉质以及更高的齿冠指示了不同的功能需求[140]。Groves 还注意到了步氏巨猿和山地大猩猩(Gorilla gorilla berigei)之间的相似性,而后者以粗糙且富含纤维质的植物为食[145]。
Ciochon等在步氏巨猿臼齿的表面提取到了可以进行分类鉴定的植硅体[144]。其中有一部分源自草本植物的营养器官,而大多数则与桑科(或者密切相关的类群)果实毛细胞相似。桑科植物为木本灌木或者树木,主要生长在亚热带和热带地区,可以产生各种可以食用的果实,如桑椹、无花果、面包果、榴莲和榕树果。最近,Qu 等报道了步氏巨猿牙齿表面的结石中发现的淀粉粒,表明步氏巨猿也食用根茎[129]。
Daegling 和Grine分析了一个步氏巨猿小样本的微磨痕特征[146]。结果表明,在现生灵长类中步氏巨猿与普通黑猩猩(Pan troglodytes) 最为相似,而其进食坚硬食物的种类(如婆罗洲猩猩Pongo pygmaeus) 的典型微磨痕构造显著不同。但是现生哺乳动物中特化为竹子食性的种类(如灰驯狐猴Hapalemur griseus 和大熊猫)不存在可以与步氏巨猿进行对比的微磨痕类型。微磨痕分析的结果并不能排除竹子是步氏巨猿主要食物的可能性,所以更为可能的是,其拥有与黑猩猩相似的广谱食性[146]。
步氏巨猿牙釉质C、O同位素值的范围很广,涵盖了共生有蹄类动物变异范围的大部分区间[2,84,105,127-129]。这些分析结果表明,步氏巨猿的食谱中涵盖了多种C3 植物来源的食物。
综合以上证据表明,步氏巨猿消耗范围很广的植物性食物,包括果实、树叶和茎,可能还包括地下营养存储器官。竹子似乎并不是食谱的重要组成成分。食物主要从枯枝落叶层和林下层获得,还有从林冠层掉落的果实。除了被子植物之外,枯枝落叶层还能够供养种类丰富的可食用真菌和蕨类。牙齿微磨痕和植硅体的分析结果和较高的龋齿率都表明果实是食谱的关键组分。然而,齿列中前齿的尺寸说明门齿并没有预处理果实和其他植物部位的功能。硕大且釉质极厚的颊齿和厚实的下颌骨适合咀嚼范围广泛的充满纤维质的粗糙植物性食物。
Zhang等以广西22处洞穴堆积化石地点为研究对象,对步氏巨猿的绝灭事件进行了综合研究[2]。在这22处地点中,不仅包括11处产出步氏巨猿化石的地点,还包括11 处时代较晚未产出步氏巨猿化石的地点。研究结果表明,步氏巨猿绝灭于29.5万~21.5万年前。70万~60万年前,由于季节性增强,环境开始变得更加多样化,森林群落的结构开始发生变化,致使步氏巨猿偏好的食物资源匮乏,不得不依赖缺少营养的备选食物,其食物的多样性大为减少。尽管如此,它们的体型却变得越来越大,摄食活动的地理范围也大为减少。因此,其种群长期面临生存压力,且不断萎缩,最终走向绝灭。
12与直立人的共存以及人类起源从亚洲到非洲的范式转换
Ciochon 等报道了越南Tham Khuyen 洞共生的直立人和步氏巨猿[81]。如前所述,该地点的步氏巨猿已被重新鉴定为猩猩[1],而其中所谓的直立人也被重新鉴定为猩猩[147,148]。中国也有多处步氏巨猿与所谓直立人或疑似人族成员共生的报道,如重庆巫山龙骨坡(Homo erectus wushanensis[103] 或H. aff. ergaster or habilis[86])、湖北建始龙骨洞(Meganthropus paleojavanicus[104])、广西田东么会洞(H. erectus[90])、广西崇左三合大洞(cf. Hominidae 或cf. Ponginae[98])、广西田东吹风洞(Hominoidea gen. et sp. indet.[89]) 等等。Ciochon 认为这些与步氏巨猿共生的直立人或者疑似人族成员并非人类,而属于一种系统发育位置仍然未知的“神秘古猿”(mystery ape)[148,149]。Ciochon 还进一步得出结论,直立人并不是中国南方地区更新世广义剑齿象—大熊猫动物群的成员之一,而真正的毫无争议的直立人化石地点都发现于秦岭—淮河一线以北,即广义剑齿象—大熊猫动物群分布范围的北方,例如陕西蓝田公王岭、安徽和县、江苏南京汤山、北京周口店等等[148,149]。截至目前,对“神秘古猿”的研究一直在进行,但始终未能验明正身。最为根本的问题还在于化石材料的局限性,因为能够支撑“神秘古猿”这一身份的形态学证据全部来自牙齿。由于形态和大小变异范围的高度重叠,亚洲更新世期间人类(hominins) 和猩猩类(pongines) 的臼齿型牙齿很难相互区分[150,151]。这也是导致“神秘古猿”久久未能展示真颜的根本原因。鉴于此,至少在“神秘古猿”验明正身之前,我们没有理由相信步氏巨猿曾经和人类共存。
在中国南方地区的早期人类被定性为“神性古猿”之后,目前中国被广为接受的最早的人类化石为1.15 Ma[152]或1.63 Ma[153]的陕西蓝田Homo erectus。如此一来,从人类起源的角度看,亚洲也被彻底笼罩在“走出非洲第一幕”(Out of Africa I) 相关理论的阴影之中。但是,如果把时间追溯到20世纪80年代之前,“人类摇篮”并不在非洲,而在亚洲。在20 世纪50 年代之前,古人类学界主流的目光都聚焦于亚洲的发现,如爪哇的“Pithecanthropus erectus”和“Meganthropus palaeojavanicus”[17],北京周口店的“Sinanthropus pekinensis”[27,154,155]。这些化石在当时被认为是人属中最早的成员,也就是说,人属的起源中心在亚洲。但从20 世纪30 年代前后开始,古人类学者们在非洲陆陆续续发现了一些上新世到早更新世期间的重要人族成员南方古猿Australopithecus 的化石,如南非Taung 的A. africanus[156]、南非Sterkfontein附近Kromdraai 的A. robustus[157]、坦桑尼亚Olduvai Gorge 的A. boisei[158] 和H. habilis[159]、肯尼亚East Rudolf 的A. biosei[160-164]、埃塞俄比亚Hadar 的A. afarensis[165] 等等。于是,从20 世纪50 年代开始古人类学界逐渐改变了观点,认为人属的起源中心不在亚洲而在非洲。尽管如此,直到20 世纪70 年代末人们仍然认为产自西瓦里克的腊玛古猿(Ramapithecus) 是最古老的人族成员(hominins)[166,167]。直到20世纪80年代初,巴基斯坦和土耳其出土的西瓦古猿头骨化石材料证实Ramapithecus 为Sivapithecus 的晚出异名,而Sivapithecus与猩猩的亲缘关系更近,与人类的亲缘关系较远[64,65,168]。至此,亚洲在早期人类演化方面的高光时刻彻底终结,取而代之的便是“走出非洲第一幕”的相关理论。如果上述产自中国南方喀斯特洞穴堆积中的“神秘古猿”最终也被证实为猩猩的近亲,就意味着亚洲这块曾被认为是大陆在早期人类演化的进程中并未担当任何形式的舞台。果真如此吗?
13展望:填补中国南方喀斯特地区上新世哺乳动物及人类化石记录的空白
中国南方地区是喀斯特地形地貌在我国的主要分布区域。根据我国新近纪的哺乳动物化石记录[169],东洋界和古北界在我国南北分治的动物地理区系格局在中新世晚期就已经初步形成。然而,一个长期困扰我们的事实就是,我国古北界的哺乳动物化石记录从晚中新世到更新世是基本连续的,而东洋界的化石记录在整个上新世期间都是空白。在云南开远、保山、禄丰、元谋、昭通等几处产自中新世晚期湖沼相或河湖相地层中与Lufengpithecus 共生的哺乳动物群之后,便是遍布整个南方喀斯特地区的产自更新世洞穴或裂隙堆积中的广义剑齿象—大熊猫动物群,目前还没有发现任何一个位于我国东洋界的上新世哺乳动物群能将二者联系起来。
究竟是哪一种早期人类最早走出非洲目前在学界仍然是存在争议的科学问题[170-172]。一直到20世纪末,“走出非洲第一幕”理论都认为,Homo均起源于东非且H. erectus 为最早走出非洲的早期人类[171]。但到了21世纪初,一系列古人类学和考古学的重大发现使得传统“走出非洲第一幕”理论受到了严峻的挑战。格鲁吉亚Dmanisi 地点1.77~1.85Ma的早期人类化石的发现[173-176] 使得H. erectus 是最早和唯一走出非洲的早期人类这一观点受到质疑。鉴于10万~6万年前的H. floresiensis 原始(与early Homo 或Australopithecus 相似)与衍生(与H. sapiens)形态特征的镶嵌模式,Jungers 等[177] 以及Brown 和Maeda[178]都认为H. floresiensis 的祖先并不是H. erectus,而是另有其“人”,并且这种理应更加原始的早期人类走出非洲的时间是在H. erectus 之前。Argue等甚至认为,H. floresiensis 的祖先走出非洲的时候还处于从Australopithecus 向Homo 演化的过程当中,这一时间是在完全的Homo 衍生形态出现之前,即2 Ma 之前[179]。Tocheri 指出,6.7 万或5 万年前的H. luzonensis 的趾骨与年代上相差两三百万年的A. afarensis 和A. africanus 非常相似,都是弯曲的;其中节指骨也与australopiths 很相似[172]。鉴于此,他认为,与H. floresiensis 一样,H. luzonensis 或许也证明了H. erectus 并不是唯一走出非洲的早期人类。Dennell 在探讨早期人类何时扩散至亚洲这一问题的过程中也注意到该区域晚上新世和早更新世化石记录的明显缺陷[171]。然而,他认为亚洲Dmanisi(1.77~1.85 Ma) 和Sangiran(1.66 Ma) 两个关键早更新世人类化石地点并不足以说明1.8 Ma 之前在如此广袤的区域内人类化石是缺失的,也不足以说明H. erectus 是最早并且唯一在现代人之前到达亚洲的早期人类。
中国目前还不存在像Dmanisi 那样古老的早期人类化石。云南“元谋人”1.7 Ma的年龄因化石层位问题而备受争议[180,181],并未得到学界的广泛接受。中国也不存在像Homofloresiensis 和H. luzonensis 那样镶嵌了early" Homo 或Australopithecus 原始特征的晚期人类。中国目前被广为接受的最早的人类化石为1.15 Ma 或1.63 Ma 的陕西蓝田H. erectus[152,153],而中国早期人类存在的最古老的考古学证据为2.12 Ma 陕西蓝田上陈的人工制品[182]。
如前所述,中国北方地区虽然化石记录相对完整,但从未出现过比蓝田Homo erectus更古老的人类的踪迹,或许说明早期人类并没有通过北方路线迁入中国。中国南方喀斯特地区虽然也缺乏上新世和早更新世的早期人类化石记录,但重庆巫山龙骨坡以及广西诸洞穴地点早更新世的“神秘古猿”会不会代表了一种形态更加原始的早期人类,他们会不会在上新世期间也涉足过中国南方地区这片土地。目前,制约我们对中国早期人类演化历史认识的关键因素就在于该区域上新世哺乳动物和古人类化石记录的空白。正如Dennell指出,证据的缺失并不等于缺失的证据[171],只有在中国南方喀斯特地区找到上新世的化石记录我们才能给出这一问题的答案。除了早期人类到达中国乃至亚洲的最早时间问题以及“神秘古猿”的真正身份问题,中国南方喀斯特地区上新世化石记录中还藏匿着诸多科学问题的答案,如禄丰古猿究竟是不是人科中一个演化分支的终点、另类的步氏巨猿与Indopithecus、Sivapithecus、Lufengpithecus 究竟是怎样的演化关系、Yuanmoupithecusxiaoyuan 究竟怎样凭借一己之力辐射出如此多样的现代长臂猿类群, 等等。
三峡地区存在两级高阶的夷平面,其中之一便是形成于上新世末期的“山原期”夷平面,海拔高度在1200~1500m;此外还有一个海拔高度更高的形成于古近纪晚期的“鄂西期”夷平面,海拔高度在1700~2000m[183,184]。作者在前期调查过程中发现了海拔高度在1350~1400m的青崖洞。该洞穴中出土了包括步氏巨猿在内的大量哺乳动物化石。由于距离青崖洞很近的两处早更新世地点建始龙骨洞和巫山龙骨坡的海拔高度均在800m左右,所以这些化石的年代应不晚于早更新世早期。理论上讲,如果早更新世早期洞穴堆积的层位像青崖洞一样位于“山原期”夷平面上,那么“山原期”夷平面的更高处以及“鄂西期”夷平面上就应该有保存上新世洞穴堆积的可能性。令人欢欣鼓舞的是,在2023年11月的前期调查中,作者的团队在两个高阶夷平面1650~1950m的海拔范围内已经找到多处不同海拔高度且有一定规模的洞穴堆积,进一步说明了在该地区寻找上新世洞穴堆积的可行性。如果在长江三峡地区高阶夷平面上可以找到上新世洞穴堆积,将会填补中国南方喀斯特地区上新世化石记录空白。
如果可以证明这一时期的化石记录中存在早期人类的化石,将会使我们重新认识“走出非洲第一幕”的相关理论。纵观鄂西地区的地形图,恩施板桥镇与屯堡乡之间、重庆奉节长安土家族乡和建始长梁镇之间、宣恩椿木营乡周边、五峰牛庄乡及以南地区、建始龙坪乡和巴东绿葱坡一带均分布有海拔在1800m 以上的“鄂西期”夷平面。而“山原期”的夷平面则主要分布在巴东野三关镇、清太坪镇和水布垭镇北部,建始的花坪及周边也有广泛分布。幸运的是,两级高阶夷平面于作者正在开展调查工作的巴东县境内均有分布。虽然“鄂西期”夷平面的范围不大,仅分布在绿葱坡镇以西的东西向狭长地区以及水布垭镇南端的蛇口山村一带,但可以成为三峡地区两级高阶夷平面洞穴调查的试点。如果真的可以在高阶夷平面上找到上新世或者更古老的洞穴堆积,无疑将为解决中国南方喀斯特地区上新世化石记录空白造成的一系列科学问题带来新的希望。
致谢:作者感谢《人类学学报》副主编吴秀杰研究员邀请作者为“纪念北京猿人第1头盖骨发现95周年”专辑撰稿,同时也感谢两位匿名审稿人富有见地的意见和建议。
