
益生菌帮助动物抵御流感的机制研究进展
2023-12-29 08:20韩美晴王迪彭先启李艳
浙江大学学报(农业与生命科学版) 2023年6期
韩美晴,王迪,彭先启,李艳,2*
(1.浙江大学动物科学学院动物预防医学研究所/浙江省动物预防医学重点实验室,浙江 杭州 310058;2.浙江大学海南研究院,海南 三亚 572025)
流感病毒(influenza virus, IFV)是正黏病毒科(Orthomyxoviridae)的代表种,常引起机体呼吸道感染,具有传染性强、传播速度快、传播范围广的特点。流感是由流感病毒引起的一种呼吸道传染性疾病,也是一种危害较大的人畜共患传染病,长期威胁着公共卫生安全。虽然当前疫苗接种是预防流感的基本手段,但因流感病毒容易发生抗原转移和抗原漂移,疫苗株和流行株的不同往往使疫苗效果不佳,因此,寻找广谱抗流感病毒的方式成为新的研究热点。
益生菌是一类在一定剂量下对机体有益的活性微生物。有研究发现,益生菌不仅能帮助机体抵抗细菌病,而且在抗病毒病、抗癌等方面也有显著效果。许多研究结果已经表明,益生菌具有增强机体抵抗流感病毒感染的作用,包括:直接阻挡病毒的黏附过程,从源头掐断病毒进入机体的途径;自身产生代谢物质或者调节机体的代谢过程,利用有效物质抵抗流感病毒侵袭;刺激机体产生免疫反应,改善免疫稳态;调节宿主免疫反应,维持肠道内菌群稳态并产生干扰素,从而抑制病毒诱导的细胞因子风暴带来的不良反应。本文介绍了流感病毒和有抗流感效果的益生菌,分类、整理和归纳了近年来有关益生菌保护动物抵御流感的研究,总结了目前研究中益生菌抵抗流感的相关机制,为未来益生菌的深入研究与应用提供参考。
1 流感的危害与防治
1.1 流感的流行与危害
流感是一种由流感病毒引起的呼吸道传染性疾病。根据流感病毒内部核蛋白(nucleoprotein,NP)和基质蛋白(matrix protein, M)的抗原性,可以将其分为甲型、乙型和丙型流感病毒,其中甲型流感病毒(influenza A virus, IAV)根据血细胞凝集素(hemagglutinin, HA)和神经氨酸酶(neuraminidase,NA)又可以分为多个亚型[1]。甲型流感病毒是引起人类流感疾病的最主要类型[2]。流感每年可导致300万~500万例重症病例和29万~65万例死亡病例[3]。1918年至今,人类历史上主要有5次流感大流行,引起大流行的流感病毒亚型主要是H1N1、H2N2以及H3N2,这几次流感大流行都是由动物源性的流感病毒跨越宿主屏障感染人而引起的[4]。此外,动物源流感(尤其是禽流感)不断地跨越宿主限制感染人,例如人流感病毒和禽流感病毒均可感染猪,猪相当于流感病毒的“混合器”;同样的,猪流感病毒(swine influenza virus, SIV)也可以反过来再感染禽和人,造成持续散发或小规模的人传人感染疫情[5]。
流感病毒是分节段的负链RNA 病毒,由于RNA 依赖性RNA 聚合酶缺乏相关的校对机制,因而流感病毒基因组在复制过程中极易产生突变,且不同病毒的基因组间易发生重组,往往造成不同类型病毒之间的抗原转移和抗原漂移[6]。有研究表明,H9N2亚型流感病毒为新型H7N9亚型流感病毒提供了内部基因[7]。尽管一些流感病毒亚型致病性低,但如果不同亚型的流感病毒在不同动物间传播并发生重组而产生新的亚型,也可能造成全球流感大流行,并伴有较高的发病率和死亡率[8],威胁着公共卫生安全和社会稳定。
1.2 流感的防治
目前,防治流感的方法主要有提前接种疫苗以及使用抗病毒药物治疗。美国食品和药物管理局(Food and Drug Administration, FDA)批准用于人类流感治疗的药物有3类:神经氨酸酶抑制剂(如奥司他韦)、M2离子通道阻断剂(如金刚烷胺)和聚合酶抑制剂。虽然接种疫苗是控制流感病毒感染的最重要手段,但其中仍存在许多挑战和不确定因素,例如,持续的抗原漂移和零星的抗原转移可能导致流感病毒产生免疫逃逸;目前预测新抗原性变化的准确性亟待提高[9-10];疫苗产生的免疫应答谱相对较窄且有效期较短[6]等。此外,耐药病毒变种不断出现,高效的抗病毒药物投入量产成为巨大难题[11-12],故迫切需要新的防疫方法或药物来减少和控制流感暴发。
2 呼吸道和肠道中的益生菌
2.1 益生菌的来源及定义
动物肠道中定植着大量微生物,并且细菌在肠道微生物群中占主导地位。肠道中的细菌大致可分为有益菌、共生菌和有害菌,其中有益菌就是我们常说的益生菌[13]。1965年,LILLY和STILLWELL首次将益生菌定义为:微生物产生的促生长因子(growth-promoting factors produced by microorganisms)[14]。直到2002年,联合国粮食及农业组织(Food and Agriculture Organization of the United Nations, FAO)和世界卫生组织(World Health Organization, WHO)将益生菌定义为:在适当剂量下,一种对宿主健康产生有益影响的活的微生物[15]。
2.2 “肠-肺轴”
益生菌可以用于治疗病毒性消化道疾病(如轮状病毒感染)和其他病毒性感染疾病(如人类免疫缺陷病毒感染),也可用于预防常见的呼吸道病毒病(如流感),减轻感染症状[15-16]。虽然益生菌主要定植在肠道内,但对系统免疫反应有着根本性的影响,它可以在身体其他黏膜部位(包括肺)定植并刺激免疫反应[17]。相比肠道菌群,呼吸道中细菌量相对较少,但外源益生菌可以在上呼吸道中定植,特别是在淋巴结和其他淋巴系统中定植。有研究表明,在口服含植物乳杆菌DSM 9843 的发酵燕麦粥8 h 后,能从6 名受试者的扁桃体上皮中分离出该菌[18],证明了植物乳杆菌DSM 9843 具有在呼吸道内定植的能力。
肠道菌群和肺组织之间的影响是双向的,此调控网络或信息交流系统被称为“肠-肺轴”(“gutlung axis”)[19]。一方面,肠道菌群产生的内毒素和代谢物可以通过血液循环影响肺部;另一方面,肺部的炎症也可以影响肠道菌群的丰富度[20-21]。健康的肠道微生物群可以通过“肠-肺轴”改善肠道免疫和肺部免疫来提高机体的免疫力,从而预防呼吸道感染,具体的作用机制可能是:1)通过大量的细菌代谢物(如脂多糖、肽聚糖和短链脂肪酸)在器官间传递信号以进行免疫调节;2)细菌本身促进肺泡巨噬细胞的极化以及辅助性T 17(T helper 17, Th17)细胞和白细胞介素-17(interleukin-17, IL-17)的产生,从而促进机体固有免疫的发展[22]。例如,感染或抗生素处理引起的胃肠道生态失调,可能改变这些微生物产生的信号并损害肺部的抗病毒免疫反应,而补充益生菌有助于增强或恢复胃肠道和肺部的有效免疫功能(图1)。虽然“肠-肺轴”是一个被普遍接受的概念,胃肠道和肺黏膜都对吸入的抗原有反应并产生交叉调节免疫反应,但局部和远端黏膜部位在产生黏膜免疫中如何相互作用仍有待确定[23]。尽管益生菌定植的系统与呼吸道病毒生存的系统有一定差异,但肠道微生物群与肺之间存在密切联系,益生菌可直接或间接抑制病毒侵入肺部,帮助免疫系统抵御呼吸道病毒。

图1 病毒性呼吸道感染下的“肠-肺轴”工作模式Fig.1 Working pattern of the “gut-lung axis” during viral respiratory infections
3 益生菌抗流感病毒的作用机制
临床试验和动物实验结果表明,特定的益生菌具有抗特定病毒的作用[24],但其潜在作用机制尚不明确。此外,菌株与菌株之间在菌株特性和功效方面的差异可能相对较大。如有些益生菌菌株可以产生抗病原体或抗黏附的物质,从而间接调节免疫系统;还有一些菌株通过改变细胞状态、刺激先天免疫和适应性免疫或增强/抑制Toll 样受体相关分子信号通路来干扰病毒。益生菌可能的抗流感病毒机制(图2)包括:1)直接抑制病毒的黏附过程;2)分泌代谢物如一氧化氮(NO)、细菌素、氨基酸等来间接抑制病毒感染;3)增强机体免疫。
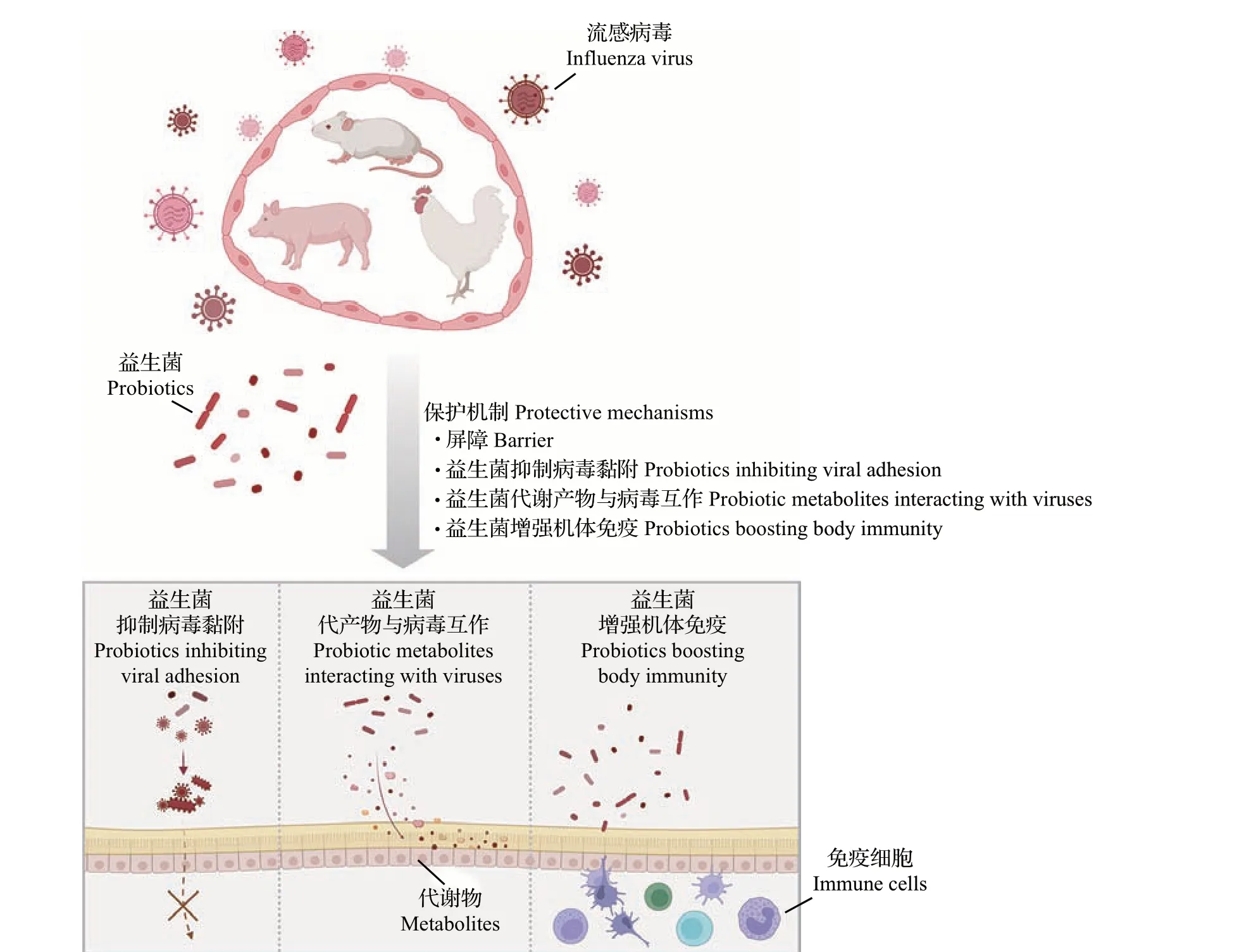
图2 益生菌在抵御流感病毒感染中可能的作用机制示意图Fig.2 Schematic diagram of possible effect mechanisms of probiotics against influenza virus infections
本文将比较和讨论近年来在不同动物模型上所做的益生菌抗流感病毒实验(附表1,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2022.07.182),分析益生菌帮助动物抵御流感的具体作用机制。
3.1 益生菌抑制病毒黏附过程
呼吸道是机体与外界直接沟通的主要通道之一,呼吸道黏膜上皮表面暴露在外环境中,是呼吸道病毒的主要黏附位置和突破口。流感病毒附着于宿主细胞是感染过程中的第一个重要步骤,而阻断其附着有利于提高宿主对流感病毒的抵抗力[25],益生菌可以直接与病毒结合,抑制其进入上皮细胞。有研究表明,乳杆菌属(Lactobacillus, LAB)和双歧杆菌属(Bifidobacterium)可以通过诱捕病毒、干扰病毒与宿主细胞受体的结合,从而维持机体健康[26]。WANG 等[27]发现,屎肠球菌(Enterococcus faecium)可以直接吸附和捕获猪流感病毒(H1N1和H3N2),对猪流感病毒在体外的繁殖起到抑制作用。但以上研究仍停留在体外实验阶段,体内实验较少或并未涉及,因此还需要更多临床数据来补充说明直接抑制病毒黏附作用对宿主的影响。
3.2 益生菌的代谢物与病毒互作
多种细菌会产生次生代谢物,这些代谢物已成为许多具有不同化学结构和生物活性药物的来源,包括抗菌药、抗癌药和抗炎药。益生菌可以通过分泌代谢物激活黏膜免疫系统以及帮助该系统成熟来间接抑制病毒感染[28]。这些代谢物包括但不限于NO、细菌素、氨基酸以及其他化合物。
无机物NO不仅是抗病毒的重要生理要素和效应分子,还能够调节肠道屏障功能、肠道蠕动和铁的运输,并与许多肠道的感染性和非感染性疾病有关联[27,29]。NO 的产生和活性的增加在宿主对抗多种病毒感染中发挥作用。WANG 等[27]的研究发现,屎肠球菌能够刺激猪肺泡巨噬细胞3D4/21 和牛肾细胞分泌NO,从而抑制流感病毒复制,这和高水平NO导致病毒产生减少的假设是一致的。细菌素是一种多肽或多肽前体,一些细菌素可通过干扰病毒入侵过程,降低细胞病变效应和病毒释放量。例如,德氏乳杆菌(Lactobacillus delbrueckii)细菌素对流感病毒的侵入和脱壳没有影响,但会减少受感染细胞中病毒蛋白的产生[30]。氨基酸是含有碱性氨基和酸性羧基的有机化合物,有研究发现其与病毒的感染和复制有关。MELANO 等[31]展示了精氨酸和赖氨酸及其酯衍生物对2 种包膜病毒(甲型流感病毒和新型冠状病毒)感染的影响,其中赖氨酸及赖氨酸酯使感染H1N1亚型流感病毒的细胞中病毒核蛋白(NP)的表达量下降,病毒RNA减少,说明该物质能够在体外抑制病毒的复制。ZHANG 等[24]在利用动物双歧杆菌(Bifidobacterium animalis)预防H7N9流感病毒感染的过程中发现,灌胃益生菌后小鼠体内的缬氨酸、亮氨酸、异亮氨酸和辅酶A的含量有所上升,后续实验证明口服缬氨酸和腹腔注射辅酶A可以在体内为小鼠拮抗流感病毒感染提供保护作用,但在体外的抗病毒效果甚微。LIU等[32]通过代谢组学和16S rRNA测序结果分析发现,异黄酮是保护小鼠抵御H1N1亚型流感病毒感染的关键物质。
随着转录组学和代谢组学等各种组学技术的飞速发展,人们可以使用更为先进的手段去检测代谢物在机体内是怎样变化的,因此代谢物在抵抗流感病毒感染方面的作用机制也会更加明晰。
3.3 益生菌增强机体免疫
免疫系统是机体行使免疫功能的机构,主要由免疫器官、免疫细胞和免疫分子组成。免疫系统可进一步分为固有免疫系统和适应性免疫系统,分别介导固有免疫和适应性免疫。益生菌可以稳定宿主的免疫反应系统,减少病毒感染时的不良反应[33]。益生菌增强机体免疫的机制主要有:增加免疫细胞及细胞因子的产生,通过Toll样受体和不同信号通路调节固有免疫,调节淋巴系统功能,增强黏膜免疫功能。
3.3.1 益生菌影响免疫细胞及细胞因子
免疫细胞是所有参与免疫应答的细胞及其前体细胞、过渡型细胞、终末效应细胞的统称。成熟的免疫细胞主要包括淋巴细胞、树突状细胞(dendritic cell, DC)、单核细胞、巨噬细胞和粒细胞。其中,在机体受到抗原物质刺激后能分化增殖、发生特异性免疫应答、产生抗体或淋巴因子的免疫细胞被统称为免疫活性细胞,主要有T细胞和B细胞[34]。免疫系统主要有2类监测病毒的细胞因子:干扰素(interferon,IFN)和白细胞介素(IL)。Ⅰ型干扰素促进被感染宿主的抗病毒状态,而其他细胞因子(包括IL)诱导炎症过程并调节免疫反应,以此产生抗病毒作用[35]。益生菌的免疫作用大致分为2种:一是免疫刺激作用,激活IL-12,诱导自然杀伤(natural killer, NK)细胞、Th1和Th2细胞来对抗感染;二是免疫抑制作用,通过Th2、DC、B 细胞和单核细胞诱导抗炎细胞因子IL-10 的产生以及诱导调节性T(regulatory T, Treg)细胞激活,以抑制炎症反应[36]。
饲喂益生菌还可以增加肺泡巨噬细胞、中性粒细胞、NK 细胞的数量并调节肺组织中促炎性细胞因子[如肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)和IL-6]的水平[27,37]。YANG等[38]研究发现,经口服植物乳杆菌CRL1506 后可以减少炎症介导的组织损伤,证明了该益生菌(CRL1506)具有帮助机体抵抗流感病毒感染的作用。植物乳杆菌DK119 可在小鼠模型中预防H1N1 和H3N2 亚型流感病毒的感染和降低死亡率,其机制是通过调节肺泡巨噬细胞和DC细胞来增强宿主对流感病毒的天然免疫反应[39]。在感染和未感染流感病毒的情况下,饲喂双歧杆菌MM-2 后,小鼠肺组织和脾脏组织中的NK 细胞活性都显著增强[40]。GROEGER 等发现长双歧杆菌(Bifidobacterium longum)细胞壁内的保护因子能够促进Ⅲ型干扰素和表面活性蛋白D的抗病毒防御反应,同时减轻Ⅰ型干扰素反应所致的肺部炎症损伤[41]。这些研究表明,对抗严重的病毒感染时,益生菌不仅能激活小鼠的固有免疫反应,而且能抑制过强的炎症反应以减少组织损伤,从而起到保护机体的作用。通过各种动物实验,益生菌已被证明具有临床应用潜力,乳杆菌和双歧杆菌可以通过口服或鼻腔给药,对包括流感在内的病毒感染具有抑制作用[42]。
浆细胞样树突状细胞(plasmacytoid dendritic cell, pDC)是识别病毒的适应性免疫细胞,pDC以产生Ⅰ型干扰素而闻名,在诱导NK 细胞的细胞活性方面发挥着重要作用,其产生的IFN-α是通过干扰病毒核酸的转录来抑制病毒复制细胞因子的[23]。许多研究表明,益生菌可以增加CD4+T细胞的数量,并调节TNF-α、IL-6、IL-8、IL-10和IL-12等细胞因子[43-46]的产生。益生菌激活适应性免疫,诱导抗原提呈细胞(antigen presenting cells, APCs)相关的促炎性细胞因子(如IL-12、IL-17、TNF-α、IFN-α)针对病毒侵袭做出防御和抵抗[47-48]。MAHOOTI 等[49]发现灌胃双歧杆菌能促进小鼠淋巴细胞增殖和免疫球蛋白G(immunoglobulin G, IgG)抗体(IgG1 和IgG2a)增加,这说明益生菌在激活Th1/Th2 免疫反应中发挥重要作用。另外一项使用小鼠模型的研究中,KAWASHIMA等[50]发现,植物乳杆菌YU可以改善小鼠体质量下降状况,减少小鼠支气管肺泡灌洗液和肺部中的病毒增殖量并且效果呈剂量依赖性;此外,植物乳杆菌能够激活Th1 的免疫反应,使小鼠支气管肺泡灌洗液和血液中的中和抗体含量升高,从而抑制流感病毒感染。SONG 等[51]分析了鼠李糖乳杆菌(Lactobacillus rhamnosus)抑制流感病毒感染的效果,发现灌胃后小鼠的存活率显著提高,并且灌胃活菌的效果比灌胃灭活菌更好。在灌胃活菌组中,受感染小鼠肺部分泌型免疫球蛋白A(salivary immunoglobulin A, SIgA)增加,促炎性细胞因子TNF-α和IL-6的表达水平降低。JUNG等[52]研究发现,相比对照组小鼠感染H3N2 亚型流感病毒后死亡率高达100%,饲喂高温灭活的干酪乳杆菌(Lactobacillus casei)DK128 的小鼠感染H3N2 亚型流感病毒后,其体质量状况好转和零死亡,且小鼠的肺部病毒载量减少,机体内的IL-6和TNF-α细胞因子表达量降低,表明高温灭活益生菌菌株作为抗病毒药物有广阔的应用前景。此外,单独或联合使用嗜酸乳杆菌、罗伊氏乳杆菌和唾液乳杆菌可以增强鸡巨噬细胞对禽流感病毒H4N6的抗病毒活性[53]。
3.3.2 益生菌刺激Toll 样受体参与模式识别
Toll 样受体(toll-like receptor, TLR)是一个庞大的模式识别受体家族,是参与固有免疫反应的一类重要蛋白质分子,而髓样分化因子88(myeloid differentiation factor 88, MyD88)是TLR信号通路中的一个关键接头分子[54]。小鼠口服乳酸乳球菌后能够产生更多的IFN-α,而IFN-α的增加和TLR9参与MyD88 信号转导以及TLR2 或TLR4 参与补充信息递呈有关。乳酸乳球菌在固有免疫调节中有“传递”功能,具有TLR9配体或TLR2、TLR4配体的类似作用[55]。鼠李糖乳杆菌介导的对感染流感病毒小鼠的保护作用是呈TLR4-MyD88 依赖性的,而激活TLR4对于Ⅰ型干扰素应答和信号传导非常重要[56]。而CHEN等[57]研究认为,粪肠球菌灌胃小鼠后可引起H1N1 感染的小鼠单核细胞趋化蛋白-1(monocyte chemotactic protein-1, MCP-1)减少,在CC趋化因子受体2(CC chemokine receptor 2, CCR2)敲除的小鼠中也观察到流感病毒的致病性降低现象。因此,尝试让小鼠口服脂磷壁酸(lipoteichoic acid, LTA)和细菌DNA,发现这2种物质均有抗流感的作用。因为LTA和细菌DNA分别为TLR2和TLR9的配体,推测由细菌成分产生的TLR配体可能调节TLR2和TLR9信号,从而调控促炎性细胞因子的表达水平,尤其是TLR2及其下游趋化因子MCP-1的表达水平。
TLR 是连接固有免疫和适应性免疫系统的重要桥梁。上述研究发现,一些益生菌通过激活TLR介导的免疫反应来参与调节机体的固有免疫反应,在免疫刺激的一线就开始发挥作用,因此有利于机体对抗病毒感染。
3.3.3 益生菌调节淋巴系统功能
淋巴系统有免疫监视和防御功能。细菌的结构成分(如脂多糖)或代谢物可以通过肠系膜淋巴系统从肠道转移到肺组织中,通过体循环调节肺部免疫[58],因此肠系膜淋巴系统的调节可以影响流感病毒感染的严重程度。派尔集合淋巴结(Peyer’s patches)是肠系膜淋巴结中的一种淋巴滤泡,YANG等[59]以植物乳杆菌为基础开发了口服疫苗,将来自流感病毒M2 的3M2e 蛋白与IgG 重链的Fc 区相融合,生成3M2e-Fc并克隆到pSIP409载体上,然后将其电转化到植物乳杆菌NC8 里,并命名为NC8-pSIP409-3M2e-Fc。用NC8-pSIP409-3M2e-Fc 接种小鼠,能够引起派尔集合淋巴结中DC细胞活化,分泌IFN-γ的T细胞数量增加以及肠系膜淋巴结中的CD8+IFN-γ+T细胞数量增加;另外,派尔集合淋巴结和肠系膜淋巴结生发中心中高水平的活化B细胞进一步激活了适应性免疫反应。长双歧杆菌MM-2 能够提高NK 细胞的活性,因为该菌刺激肠相关淋巴组织(如派尔集合淋巴结和肠系膜淋巴结)产生信号(包括细胞因子和Th1 细胞)并传递到肺组织,从而达到抵御流感病毒感染的效果[40]。
3.3.4 益生菌增强免疫球蛋白反应
免疫球蛋白A(immunoglobulin A, IgA)是黏膜免疫的重要参与者[58],益生菌菌株能够刺激机体产生抗病毒的免疫球蛋白[60]。SONG 等[51]以小鼠为动物模型,分析了鼠李糖乳杆菌M21对流感病毒感染的防御作用。结果表明,该菌显著增加了SIgA的水平,成功减轻了受感染小鼠肺组织的慢性损伤程度,从而提高了小鼠的存活率。有关的临床研究表明,双歧杆菌能使IFN-γ、IL-2、TNF、IgA和SIgA的水平升高[23]。高温灭活的植物乳杆菌AYA也可以使小鼠肺泡灌洗液中的IgA水平显著升高[45]。免疫球蛋白分泌增多是机体免疫力增强的表现,也证明了益生菌能够通过改善免疫状况来抵御流感病毒的侵袭。
机体的免疫应答机制非常复杂,免疫细胞通过识别、摄取、加工处理抗原和调节效应因子启动适应性免疫应答,而T、B细胞参与细胞因子的调节等可以进一步刺激或抑制免疫应答。机体内复杂的免疫细胞都以独特的方式发挥着不同的作用,一些细胞因子的上调和下调也不仅是一种免疫应答在发挥作用,而可能是多种免疫应答同时作用所致,因此益生菌具体通过哪些途径来刺激机体免疫还有待进一步探索。
3.3.5 益生菌调控抗流感病毒相关因子基因的表达
益生菌在机体定植后,可通过调控机体内组织或细胞内抗病毒基因的表达来发挥抗流感病毒的功能;也可以诱导免疫反应基因表达,这些基因可以促进免疫刺激或免疫调节表型表达[53]。KISO等[61]发现灭活的戊糖乳杆菌(Lactobacillus pentosus)b240对小鼠肺组织基因转录有影响,与酰基辅酶A介导的代谢和与蛋白质刺激反应相关的基因表达下调,其中表达下调的Egr1基因与表达上调的Rsad2基因同属抗流感病毒基因。NAKAYAMA等[62]研究表明,小鼠口服加氏乳杆菌(Lactobacillus gasseri)SBT2055后,对清除病毒至关重要的Mx1、Oas1a基因在肺组织中的表达量增加;口服加氏乳杆菌LG2055 也可减少感染后流感病毒H9N2 亚型在肺组织中的复制,说明LG2055 能通过诱导抗流感病毒基因的表达来下调病毒的复制,从而有效预防流感病毒的感染。GAO 等[63]从嗜酸乳杆菌ATCC 4356 中分离出新的生物抗病毒材料——S层蛋白,其可增加DC细胞中干扰素刺激基因Mx1、Isg15、Ddx58的表达,诱导DC细胞产生强烈的应答,抑制流感病毒H9N2亚型的入侵。SHOJADOOST等[53]研究结果显示,使用乳杆菌刺激巨噬细胞后,调控IL-1β、IFN-γ和IFN-α等细胞因子以及干扰素调节因子-7(interferon regulatory factor-7, IRF-7)、2,5-寡腺苷合成酶(2, 5-oligoadenosine synthetase, OAS)和干扰素诱导的跨膜蛋白M3(interferon induced transmembrane protein M3, IFITM3)的基因表达显著增加。上述研究通过使用体外模型证明了乳杆菌可以有效增强鸡巨噬细胞的抗流感病毒反应。
4 小结与展望
流感病毒是常见的人畜共患病原,对人类和动物健康造成严重威胁。甲型流感病毒基因组由8段负义单链RNA 组成,易发生变异和基因片段间重组,流感大流行就是甲型流感病毒出现新亚型或旧亚型重现引起的,因此预防和控制流感成为一项非常艰巨的任务。在“替抗减抗”的大环境下,益生菌可以平衡肠道菌群组成、加强肠道屏障功能和产生保护性免疫反应等,应用优势日益凸显。现有的研究发现,饲喂益生菌能够在不同动物模型上抵御流感病毒感染,包括猪、鸡、小鼠和秀丽隐杆线虫等,但其作用机制尚不明确。有研究认为益生菌可广泛引起机体非特异性和特异性免疫反应,还有研究认为是益生菌的代谢物在发挥作用,但仍需要大量实验为证明益生菌在缓解疾病和改善疾病预后中的作用提供基础数据。
如今,益生菌可以添加在多种食品中,也可以作为人们的保健品和医药产品,但如何正确和安全使用益生菌仍然存在激烈的争议。最早在2008年,BESSELINK 等[64]在一项随机实验中发现益生菌处理组的小鼠死亡率较高,虽然不能直接证明这种高死亡率与益生菌相关,但这种反常的情况也引起了很多关注。此外,细菌的代谢物可能在治愈疾病中起反作用,例如乳杆菌会产生吲哚类代谢物,从而激活肿瘤相关巨噬细胞上的芳香烃受体,抑制CD8+T细胞释放干扰素杀伤肿瘤细胞的效果,从而导致胰腺导管腺癌患者病情加重[65]。益生菌在全球的市场份额迅速扩大,但其相关危害和风险需要专家去识别、讨论和论证,因此,如何正确使用益生菌仍是今后关注的重点。
随着益生菌相关研究的深入,人们发现并非所有益生菌都具有抗病毒能力,并且其效果具有疾病特异性和菌株特异性,因此每种益生菌的作用机制仍有待阐明并需要进行有效的临床评估。如何选择最能应对流感大流行的菌株(单独或联合剂量),如何为一个群体选择标准菌株,目前很难得到统一的答案,因为现有的研究方案,包括干预时间、实验模型(儿童、成人或动物)、细菌剂量(1×106~1×1010CFU/d)、生存环境及给药方式等仍存在差异。虽然我们列出了一些相关研究的益生菌菌株,但不同的菌株可能会有不同的作用机制,而了解和挖掘益生菌对应作用机制对于抗病毒研究具有重要的意义。因此,深入了解和挖掘每种益生菌的具体作用机制对于推动益生菌抗病毒研究具有重要的意义,这将有助于确立更为精准和有效的益生菌应用策略,为未来的抗病毒研究提供科学依据。
猜你喜欢
基层中医药(2020年12期)2020-07-22
广东医科大学学报(2020年6期)2020-02-06
现代检验医学杂志(2016年3期)2016-11-15
现代检验医学杂志(2016年1期)2016-11-12
兽医导刊(2016年12期)2016-05-17
分析测试学报(2015年6期)2016-01-13
中国病理生理杂志(2015年8期)2015-12-21
中国当代医药(2015年30期)2015-03-01
癌变·畸变·突变(2014年2期)2014-03-01
当代畜禽养殖业(2014年10期)2014-02-27
