
秸秆覆盖与施磷对西南冬小麦叶肉导度和CO2同化效率的影响
2023-12-26 02:44马宏亮黄秀兰祝婷婷樊高琼1杨洪坤1
生态学报 2023年23期
贺 鹏,谢 炜,马宏亮,黄秀兰,祝婷婷,樊高琼1,,杨洪坤1,,*
1 西南作物基因资源发掘与利用国家重点实验室,成都 611130
2 农业农村部西南作物生理生态与耕作重点实验室,成都 611130
磷素在能量代谢中起着至关重要的作用,也是叶片核酮糖-1,5-二磷酸(RuBP)再生过程中磷酸化的重要底物[1]。西南麦区是我国第三大小麦产区,70%小麦分布在丘陵旱地,其气候冬干春旱,土壤速效磷含量低[2]。缺磷严重限制作物光合作用,直接影响生物量和产量[3]。磷在土壤中的移动性差,且无机磷容易被吸附固定,导致磷肥的利用率低[4]。此外,磷矿储量逐渐枯竭,过量的磷肥投入导致土壤质量退化,还引起地表水富营养化[5]。在麦玉轮作系统中,采用玉米秸秆粉碎覆盖还田,不仅可有效缓解长期施用化肥造成的土壤退化,促进土壤磷形态的转化并提高土壤速效磷的含量[6—8],能改善群体结构,延缓叶绿素降解,延长光合时间,提高光合速率[9—10]。
花后旗叶光合在作物产量形成中起着关键作用[11],其光合效率的高低取决于气体交换、CO2扩散和碳固定,这些因素都受到磷素有效性的影响[12—13]。徐龙龙[14]等发现玉米覆盖提高小麦的光合势和光合速率从而实现增产。岳俊芹[15]等研究表明随施磷量(P2O50—225 kg/hm2)的增加,小麦花后同化物、光合速率及产量均不同程度增加。Zhang[16]等研究认为秸秆覆盖增加花后净光合速率、气孔导度、最大羧化速率和最大光合电子传输速率。前人研究已明确缺磷条件下光合作用降低归因于气孔导度或叶肉导度的降低[17]。磷胁迫下,大气CO2向叶绿体的扩散受到限制是光合作用降低的主要原因[18]。但也有研究表明,磷素限制大豆光合作用的主要机制不是CO2扩散,而是叶绿素、光化学和羧化效率的降低[9]。然而,秸秆覆盖与施磷下小麦旗叶叶肉导度、CO2同化效率和叶片磷含量的关系仍然不清楚。
前期研究表明:秸秆覆盖配施固定氮钾肥不仅可以蓄水保墒缓解分蘖期干旱,还可增加土壤速效磷、旗叶净光合速率和产量[19—21]。在前期研究基础上,本研究采用秸秆覆盖和磷素水平二因素裂区实验,明确小麦旗叶光响应曲线、叶肉导度、CO2同化效率和光合限制因素对叶片磷含量的响应,阐明秸秆覆盖与施磷对西南冬小麦光合限制因素的影响机制,以期为西南冬小麦高产磷高效提供理论依据和技术支撑。本研究假设秸秆覆盖与施磷提高小麦旗叶比叶重和磷素含量,增加旗叶叶肉导度和CO2同化效率,从而提高旗叶的净光合速率。
1 材料与方法
1.1 试验地概况
本试验于2020—2022年在四川省眉山市仁寿县珠嘉镇踏水村(30°04′N, 104°13′E)进行。该地区属亚热带季风湿润气候,丘陵地貌,海拔482m,前茬作物为夏玉米。年均气温17.9℃,年均日照986.6h,无霜期312d。2021—2020和2021—2022年小麦生长季总降雨量分别为195.5mm和228.4mm,两个生长季均出现严重的冬春干旱。土壤类型为碱性石灰性紫色土,供试土壤(0—20cm)贫氮、缺磷和富钾是该地区土壤的主要特征(表1)。

表1 供试土壤基础理化性状
1.2 试验设计
本试验采用二因素裂区设计,主区为玉米秸秆粉碎覆盖(SM)与不覆盖(NSM),裂区为三个磷素水平。在麦玉轮作系统中,夏玉米5月中旬播种,8月底收获后将玉米秸秆粉碎覆盖在各试验小区,覆盖量为8000kg/hm2。裂区三个磷素水平为:0(P0)kg/hm2、75(P75)kg/hm2和120(P120)kg/hm2磷肥,共计6个处理,每个处理均设3次重复。供试品种为川麦104,由四川省农业科学院提供。小麦于每年10月30日采用开沟点播的方式播种,2021年5月9日和 2022年5月6日收获。行距20cm,穴距10cm。氮肥为普通尿素(N 46.2%),磷肥为过磷酸钙(P2O512.5%),钾肥为氯化钾(K2O 60.0%),氮肥和钾肥用量分别为150 kg/hm2和75 kg/hm2,60%氮肥和全部磷、钾作为基肥,其余部分氮肥在拔节期作为追肥,施肥方式采用撒施。小麦收获后种植夏玉米,品种为正红6号。该区域为典型雨养农作系统,主要通过秸秆覆盖蓄积土壤水分满足小麦水分需求。其它栽培管理措施与当地高产栽培管理技术一致。商用除草剂、杀虫剂和杀菌剂在分蘖后每隔一个月进行预防,以避免产量损失。
1.3 测定项目与方法
1.3.1旗叶比叶重、磷含量和净光合速率
记录各处理小麦开花期,于同天上午的8:00至11:00选择生长和受光一致且完全展开小麦主茎旗叶进行净光合速率、光响应曲线、CO2响应曲线和荧光参数测定,测定时随机选取整张叶片的中间部分,各处理重复测定5次。采用LI-6800便携式光合作用系统(美国LI-COR公司,内布拉斯加州,美国),在1200μmol m-2s-1光照强度、65%±5%相对湿度和400μmol m-2s-1的大气CO2浓度(Ca)下记录净光合速率(Pn)、细胞间CO2浓度(Ci)和气孔导度(gs)。
完成叶片气体交换测量后,每个处理收集10片旗叶用于测定旗叶比叶重(LMA)和磷(P)含量。从每片旗叶剪下叶片中间部位5cm长,测量宽度,计算旗叶面积。旗叶在105℃杀青30min,70℃下干燥至少48h,然后称重计算旗叶的LMA,使用钒钼黄比色法[22]进行分析确定单位质量旗叶P含量。
旗叶的比叶重 LMA(g/m2)为单位旗叶面积的干物重,其计算公式[23]为:
LMA=DW/LA
其中,LA为旗叶面积(cm2),DW为干物重(g)。
单位叶面积磷含量PA(g/m2),其计算公式为:
PA=PM×LMA/100
其中,PM(mg/g)为单位质量的磷含量,LMA为比叶重(g/m2)。
1.3.2旗叶光响应曲线
采用便携式光合作用系统(LI-6800)测量旗叶光响应曲线。在2000、1800、1600、1400、1200、1000、800、600、400、200、100、50、0μmol m-2s-1的光量子通量密度(PPFD)和Ca浓度为400±5μmol/mol下测定旗叶的光响应曲线。使用非线性双曲线模型[24]拟合叶片光响应曲线,如下所示:
其中,α表示表观量子产率(AQY),I表示光量子通量密度,Pnmax表示最大净光合速率,Rn表示暗呼吸速率。使用SPSS 19.0进行线性回归分析,PPFD为0至2000μmol m-2s-1。该线与x轴(光合作用有效辐射,PAR)的交叉点是光补偿点(LCP,μmol CO2m-2s-1)。沿y轴交叉点的相应x轴值为光饱和点(LSP,μmol CO2m-2s-1)。
1.3.3旗叶荧光参数
采用便携式调制叶绿素荧光仪(PAM-2500,德国Walz公司,巴伐利亚州,德国)测定叶绿素荧光诱导动力学参数。先用暗适应叶夹固定叶片,控制环境温度在30℃左右,相对湿度50%—60%,充分暗适应20min,然后测定暗适应下的叶绿素荧光参数(叶片的初始荧光(Fo)、最大荧光产量(Fm)、可变荧光(Fv)和实际光化学效率(ΦPSII))。
1.3.4旗叶叶肉导度与叶绿体CO2浓度
采用便携式光合作用系统(LI-6800XT)测量旗叶CO2响应曲线。其中在1200μmol m-2s-1的稳定PPFD下,Ca从500μmol/mol依次降低至400、300、200、150、100和50μmol/mol,并从50μmol/mol增加至400、600、800、1000、1500和2000μmol/mol。每个处理取10 片旗叶,用锡箔纸包裹旗叶,迅速用液氮冷冻5min,-80℃ 冰箱储藏用于Rubisco酶活测定。Rubisco活性采用蛋白质检测器酶联免疫吸附试验(ELISA)试剂盒(KPL,Gaithersburg,USA)测量。
采用Harley[25]等的方法计算叶肉电导(gm)和叶绿体CO2浓度(Cc)。
其中Pn表示净光合速率;I*表示线粒体呼吸缺失时的CO2补偿点,根据Bernacchi[26]等计算为44.04 mol/mol;Ci表示细胞间CO2浓度;Rd表示光中的非光呼吸呼吸,计算为暗呼吸速率(Rn)的一半[27]。
1.3.5旗叶光合限制因素
Pn的相对限制,包括气孔导度(Ls)、叶肉导度(Lm)和生化(Lb)限制,计算公式如下[28]。
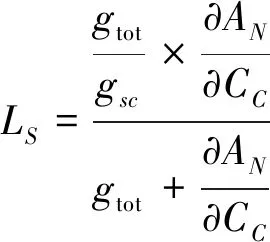
其中,gtot表示CO2从环境空气到叶绿体的总传导率(1/gtot=1/gsc+gm),gsc表示气孔对CO2的传导率(gsc=gs/1.6)。∂AN/∂CC计算为Cc范围为50—100 mol/mol时Pn-Cc响应曲线的斜率[29]。
1.4 数据处理
试验数据用Excel 2016进行整理计算,用SPSS 19.0进行统计分析及相关性分析, 最小显著性差异法对不同处理间差异显著性进行多重比较(P<0.05),采用 Origin 2019b作图。
2 结果与分析
2.1 秸秆覆盖与施磷对旗叶比叶重、磷含量和净光合速率的影响
秸秆覆盖与施磷显著提高旗叶比叶重(LMA)、磷含量(单位面积的磷含量PA和单位质量的磷含量PM)和净光合速率(Pn),二者的交互作用对其影响不显著(P>0.05,表2)。与不覆盖(NSM)相比,秸秆覆盖(SM)条件下小麦旗叶LMA、PA、PM和Pn分别提高4.1%、16.9%、12.2%和6.9%。与不施磷(P0)相比,施磷75kg/hm2(P75)小麦旗叶LMA、PA、PM和Pn分别提高3.9%、23.3%、18.5%和4.9%,施磷120kg/hm2分别提高了4.0%、24.5%、19.7%和9.8%。说明秸秆覆盖配施磷肥提高小麦旗叶磷含量,从而促进旗叶干物质的积累和净光合速率。

表2 秸秆覆盖与施磷对旗叶比叶重、磷含量和净光合速率的影响
2.2 秸秆覆盖与施磷对旗叶光响应曲线的影响
秸秆覆盖与施磷显著影响不同光强下的旗叶净光合速率(图1)。当光合有效辐射(PAR)<300μmol m-2s-1时,Pn随着PAR的增加而线性增加。随着PAR的进一步增加,旗叶出现最大净光合速率(Pnmax)出现。在P75和P120下,Pn在PAR为1400μmol m-2s-1下检测到最大值,然后缓慢下降并保持在较高水平。在P0,当PAR>1200μmol m-2s-1时,Pn随着PAR的持续增加而不改变。由表3可知,两个试验年度,SM较NSM的小麦旗叶最大净光合速率(Pnmax)、表观量子效率(AQY)、光补偿点(LCP)和光饱和点(LSP)分别提高了10.0%、27.0%、18.8%和18.2%。P75与P120较P0的Pnmax、AQY、LCP和LSP分别提高了5.3%—10.1%、16.6%—22.5%、20.7%—27.6%和13.9%—19.5%。说明秸秆覆盖与施磷主要影响高PAR下对小麦旗叶的光合效率。

图1 秸秆覆盖与施磷对旗叶光响应曲线的影响
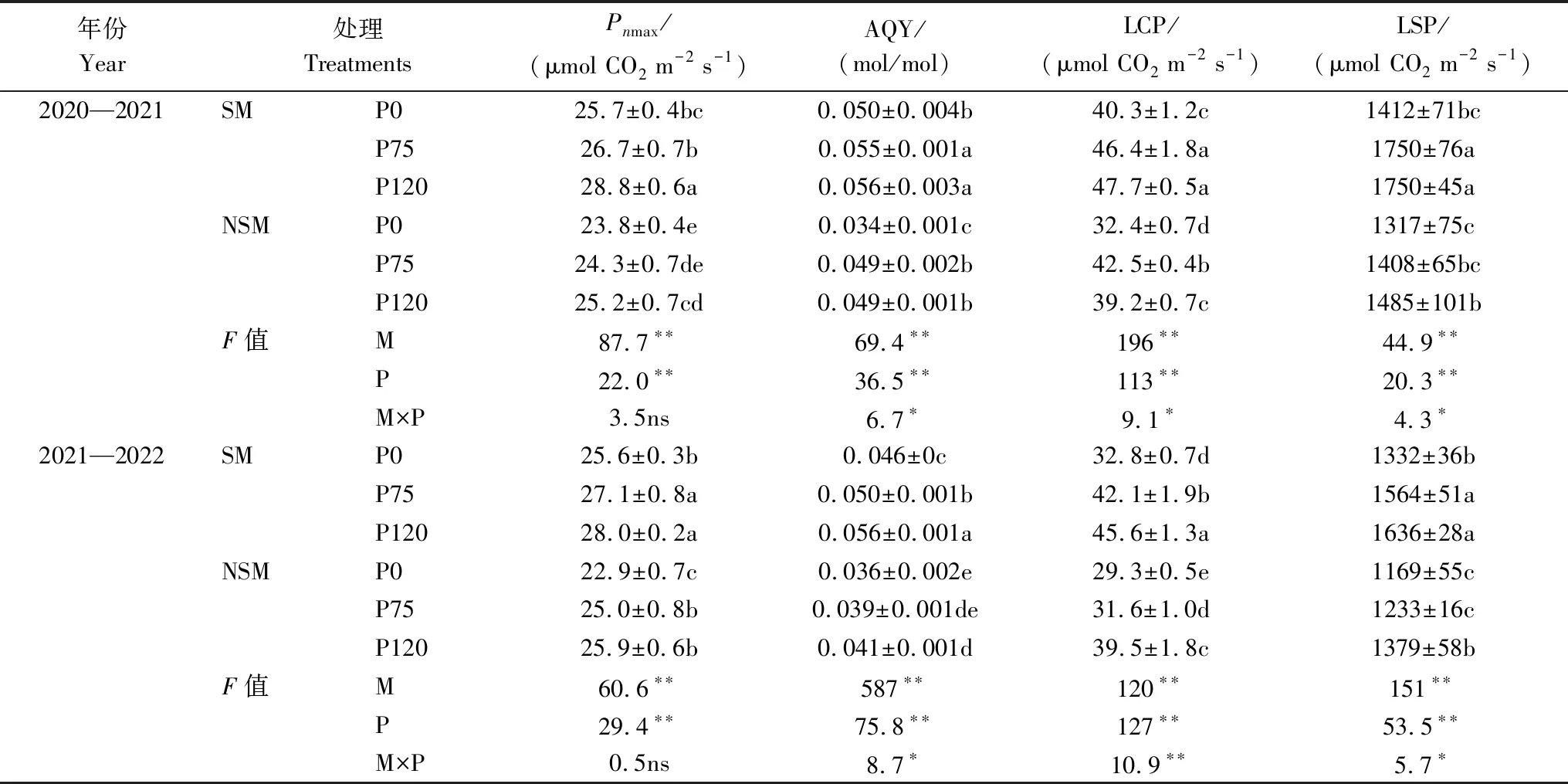
表3 秸秆覆盖与施磷对旗叶光响应参数的影响
2.3 秸秆覆盖与施磷对旗叶叶绿素荧光参数的影响
由图2可知,秸秆覆盖与施磷显著影响旗叶光系统II的最大光化学效率(Fv/Fm)和实际光化学效率(ΦPSII),且二者的交互作用对ΦPSII影响显著。随着施磷量的增加,不同处理的Fv/Fm和ΦPSII呈现出增加的趋势,且均以秸秆覆盖配施磷肥120kg/hm2最高。两个试验年度,SM较NSM的Fv/Fm和ΦPSII分别提高了8.6%和17.6%,P75较P0的Fv/Fm和ΦPSII分别提高了6.9%和16.8%,P120较P0的Fv/Fm和ΦPSII分别提高了8.3%和23.8%。随着施磷量的提高Fv/Fm和ΦPSII呈上升趋势。说明秸秆覆盖与施磷保护光系统II的活性,增加光系统II的最大光化学效率和实际光化学效率。

图2 秸秆覆盖与施磷对旗叶荧光参数的影响
2.4 秸秆覆盖与施磷对旗叶光合参数的影响
秸秆覆盖与施磷显著增加旗叶最大羧化效率(Vcmax)、初始羧化效率(CE)和核酮糖-1,5-二磷酸羧化酶(Rubisco)活性,但不影响电子传递效率(Jm)(图3)。SM较NSM的Vcmax、CE和Rubisco分别提高了17.3%、6.8%和32.0%。与P0相比,P75的Vcmax、CE和Rubisco分别提高了27.9%、6.5%和30.9%。P120分别提高了35.3%、9.1%和61.7%。施磷显著提高了旗叶CO2传导效率,但秸秆覆盖及与施磷的交互作用对胞间CO2浓度(Ci)、叶绿体CO2浓度(Cc)和叶肉导度(gm)没有显著影响。与NSM相比,SM的气孔导度(gs)提高了5.3%。与P0相比,P75与P120的Ci、gs、Cc和gm增幅分别达到1.4%—3.6%、4.8%—13.1%、9.0%—10.2%和16.0%—16.9%。

图3 秸秆覆盖与施磷对旗叶光合生理参数的影响
2.5 秸秆覆盖与施磷对旗叶光合限制因素的影响
由表4可知,秸秆覆盖和施磷显著影响旗叶气孔限制(Ls)、叶肉限制(Lm)和生化限制(Lb)。然而,二者的交互作用对旗叶Pn的限制因素没有显著影响。秸秆覆盖下净光合速率的提高主要受生化限制(0.46—0.55)的影响,施磷降低Ls和Lm显著降低,而Lb显著增加。在NSM条件下,Ls和Lm是不施磷条件下Pn提升的主要限制因子。综上,秸秆覆盖与施磷提高旗叶净光合速率是由于降低了气孔限制和叶肉限制值。
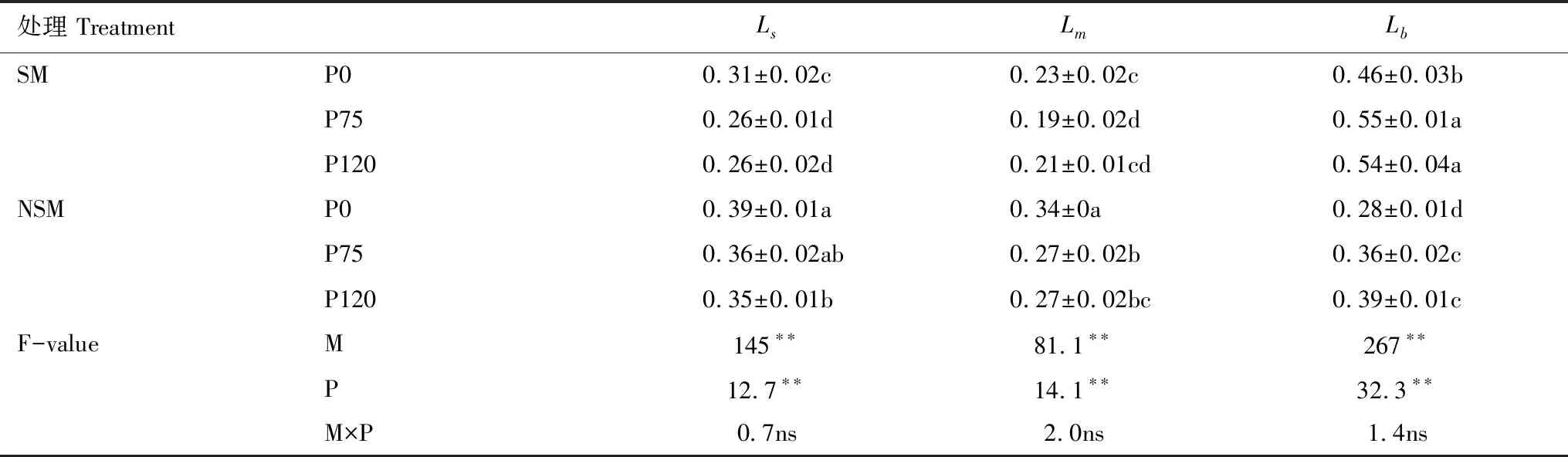
表4 秸秆覆盖与施磷对旗叶净光合速率相对限制的影响
2.6 旗叶磷含量和比叶重与光合生理参数的关系
旗叶Pn与LMA、PA、PM呈极显著正相关(表5),说明小麦旗叶LMA和磷含量的增加有利于旗叶Pn提高。旗叶Pn与ΦPSII、Vcmax、Rubisco、Ci和Cc呈极显著正相关,说明小麦旗叶Pn的提高与羧化效率、叶肉导度和CO2同化密不可分。旗叶Pn与gs和gm呈极显著正相关,说明小麦旗叶Pn的增加主要是由于气孔导度和叶肉导度的提高。由图4可知,旗叶Pn、gs和gm随着PA、PM的升高而升高。旗叶中的PA每增加1个单位,旗叶的Pn、gs和gm分别增加4.76、0.09和0.15个单位。旗叶中的PM每增加1个单位,旗叶的Pn、gs和gm分别增加1.31、0.03和0.04个单位。说明在秸秆覆盖与施磷处理下,旗叶磷含量的增加导致旗叶gs和gm的增加,最终提高旗叶Pn。

图4 秸秆覆盖与施磷处理下磷含量与Pn、gs和gm的关系

表5 旗叶磷含量和比叶重与光合生理参数的相关关系
3 讨论
3.1 秸秆覆盖与施磷提高旗叶磷素含量和比叶重,促进旗叶的光能利用
磷在调节核酮糖-1,5-二磷酸(RuBP)再生、羧化效率和气孔导度方面起着关键作用[30],但西南丘陵旱地石灰性紫色土富钾贫磷,尤其是土壤速效磷缺乏[31]。本研究发现,秸秆覆盖和施磷提高小麦旗叶磷含量、LMA和Pn。究其原因,秸秆覆盖显著提高土壤速效磷含量,增加土壤蓄水保墒能力,利于小麦根系生长,维持较高的根系活力,扩大根系吸收磷素面积,从而提高根系磷吸收和磷素吸收效率[32—33]。旗叶磷含量和LMA与Pn呈极显著的正相关,与先前的研究一致[34]。一定施肥量范围内施磷能增加旗叶叶绿素含量,改善旗叶光合性能[35],但施磷过多不能继续提高旗叶光合速率[36]。LMA增加说明秸秆覆盖与施磷增加旗叶厚度,但由于CO2从空气扩散到叶绿体与叶片厚度呈负相关。因此,植物遭受非生物胁迫时较厚的叶片并不总是伴随高光合能力[37]。较薄的叶片通过减少有效CO2路径长度有利于CO2固定,但这不足以补偿缺磷导致的叶片光合作用减少[38]。
植物叶片的光饱和点与光补偿点反映了植物对光照条件的要求。光补偿点较低、光饱和点较高的植物对光环境的适应性较强[39]。小麦旗叶LMA和磷浓度的改变导致旗叶对外部光环境的适应能力的改变,在本试验中发现,秸秆覆盖和磷肥增加了小麦旗叶Pnmax、AQY、LCP和LSP,光合作用过程中最大量子效率在0.08—0.125之间。本文对光合曲线的拟合得出在0.036—0.056之间,小于理论上的最大值,因此是合理的。在遮荫和氮肥条件下观察到类似的适应策略[40—41]。说明秸秆覆盖与施磷改善了小麦灌浆期旗叶的光合作用能力,增强了小麦旗叶对强光适应能力,同时利用弱光的能力也有所提升,表现出更高的光合效率,与孙旭生等[39]研究结果一致。这是秸秆覆盖与施磷处理下小麦具有较强光合能力的光合生理基础,也就是叶片磷素营养与光合效率密切相关[39],如Rubisco[42]、蔗糖磷酸合成酶和蔗糖合成酶[43]。综上,秸秆覆盖和磷肥增加小麦旗叶在相同质量或相同面积下的磷浓度和LMA,提高旗叶对外部光环境的适应能力和光能利用能力。
3.2 秸秆覆盖与施磷增强旗叶叶肉导度和羧化效率,提高CO2同化效率
植物叶片的叶绿素荧光参数能直接或间接反应光合作用的原初反应、电子传递以及CO2同化过程[44]。有研究显示,当植物处于逆境时,叶片叶绿素荧光参数ΦPSII和Fv/Fm等则呈下降趋势,说明此时光系统Ⅱ反应中心可逆性失活[45]。缺磷会损害光系统II(PSII)、光系统I(PSI)、电子传输、降低类囊体膜流动性、Rubisco活性和细胞膜稳定性[46]。在本研究中,秸秆覆盖与施磷显著提高花后旗叶的ΦPSII和Fv/Fm。说明小麦旗叶在秸秆覆盖与施磷条件下对土壤磷素的缺乏具有一定的适应性反应,能应对磷素缺乏以热耗散的形式做出自我保护,防止光合机构的破坏。进一步说明生育后期秸秆覆盖与施磷保护小麦旗叶光系统Ⅱ反应中心,避免受磷素缺乏胁迫可逆性失活,光合机构受到破坏,从而提高旗叶光系统Ⅱ的光化学效率和净光合速率。
气孔因素是可恢复胁迫水平下Pn降低的主要原因,非气孔限制被认为是不可恢复胁迫条件下Pn下降的主要原因[47—49]。本研究中,秸秆覆盖与施磷显著提高气孔导度、叶肉导度和羧化效率。这表明,旗叶磷素缺乏降低羧化效率和电子传递效率,限制CO2向气孔和细胞间隙空间的扩散,导致Pn的降低。气孔关闭和植物扩散过程中的进一步限制如细胞壁增厚及细胞间隙增大是植物对磷缺乏的常见反应[50]。在本研究中也发现,秸秆覆盖与施磷显著提高旗叶的Rubisco酶活,增强了旗叶的碳固定,这与秸秆覆盖与施磷条件下小麦处于充足的水分和磷营养供给状态有关。秸秆覆盖和磷肥降低气孔限制和叶肉限制值,生化限制是限制Pn增加的主要因子,进一步说明叶肉导度和CO2同化效率在提高Pn方面的关键作用。以上研究证实了的假设,即秸秆覆盖和施磷提高小麦旗叶磷含量,增强叶肉导度和CO2同化效率,并提高净光合速率。
4 结论
通过两年大田定位试验,研究秸秆覆盖与施磷对西南麦区冬小麦叶肉导度与CO2同化效率的影响。主要的结论如下:(1)秸秆覆盖与施磷增加小麦旗叶磷含量和比叶重,从而提高净光合速率;(2)秸秆覆盖与施磷显著提高小麦旗叶的Pnmax和AQY,促进其在高PAR下的光能效率;(3)秸秆覆盖与施磷显著提高小麦旗叶ΦPSII和Fv/Fm,增加光系统II的光化学效率;(4)秸秆覆盖与施磷显著增加气孔导度、叶肉导度和羧化效率,生化限制是限制小麦光合速率增加的主要因子。秸秆覆盖与施磷提高旗叶磷含量和比叶重,增加叶肉导度和CO2同化效率,从而提高净光合速率。
猜你喜欢
生态学报(2024年1期)2024-01-25
作物学报(2022年9期)2022-07-18
蔬菜(2022年12期)2022-01-01
干旱地区农业研究(2021年6期)2021-12-08
今日农业(2021年17期)2021-11-26
浙江农业科学(2019年11期)2019-11-09
农村百事通(2019年1期)2019-01-30
福建农林大学学报(自然科学版)(2019年1期)2019-01-25
西南农业学报(2016年6期)2016-04-16
农家顾问(2014年11期)2014-12-01
