
动物的分节现象
2023-12-21 03:09:06李海云
生物学杂志 2023年6期
李海云,李 瀛
(1.华南农业大学 动物科学学院,广州 510642; 2.华南农业大学 生命科学学院,广州 510642)
动物学教材中仅简单明确地在环节动物章节中介绍了同律分节和异律分节,在节肢动物章节中介绍了异律分节。而诸如多节绦虫的链体与链涡虫的节裂体是怎么回事?脊索或脊椎动物的分节与无脊索动物的分节有什么区别?各类动物的分节主要涉及的胚层等问题常使教学陷入困境。动物的分节现象极为复杂并不仅限于同律分节和异律分节。动物分节现象吸引了不少发育生物学家和演化生物学家的关注和研究[1-6]。本文就动物分节现象的定义、国内外动物分节现象用词的混乱、动物分节的起源、分节的类型及其演化意义进行综述。旨在使读者对动物分节现象有较全面的理解,同时为动物学教学提供有益的参考。
1 分节现象 (segmentation phenomenon)的定义
关于分节现象的定义,传统认为是指动物身体沿纵轴分成许多相似的部分,每个部分称为一个体节[7-17]。这实际上是同律分节的定义。动物身体由许多体节、节段、节片或体环构成,均为分节现象。分节的体制有许多共同的形态特征,典型同律分节的环节动物由一系列重复的身体单位构成,每个单位可能都由来自不同器官系统如体腔、排泄、生殖、神经、循环、附肢和外部的环等元素组成,体表以节间沟为界或无分界。分节现象从两侧对称、三胚层的扁形动物开始出现,直至脊索动物。这些动物具有各种类型的分节现象,有些分节现象主要涉及外胚层,有些主要涉及中胚层,有些则既涉及外胚层也涉及中胚层,有的则主要涉及内胚层和中胚层。
2 国内外动物分节现象用词的混乱状况
国内动物学教材中仅在环节动物门中提及同律分节(homonomous segmentation[7]或homonomous metamerism[8-9]或两者混用[9])和异律分节(heteronomous segmentation[7]或heteronomous metamerism[8-9]或两者混用[4]),在节肢动物门中提及异律分节[7-9]。
国外的分节现象相关术语比较混乱,使用过的有“metamery”[10]“segmentation”[11-14]“metamerism”[15],有时也有混用[16-17]。也有学者认为“segmentation”适用于环节动物、节肢动物和脊索动物(或脊椎动物)的分节,并把这些动物的身体称为分节的身体体制(segmented body plan),其他动物的分节则使用“metamerism”[18-19]。有的将动物的分节区分为真分节“true segmentation”和表面分节“superficial segmentation”[20];也有的区分为分节“metameric”和假分节“pseudometameric”;有的则将“segmentation”用于描述环节动物、节肢动物和脊椎动物的分节,将“metamerism”用于描述绦虫的分节,其他动物的分节则采用“serial repetition”或“series”[19,21]。
国外文献中体节“segments”常被当作“metameres”[14]。然而,一些作者认为“metamere”仅指体腔的细分,并将其与术语“segment”区分开来,认为“segmentation”是“metamerism”的特例[22]。近期“segmentation”被广泛应用,甚至有人将钵水母水螅体无性繁殖的横裂体也当作“segmentation”[23]。
由于初期人们认为连续排列的体腔对分节现象概念起着至关重要的作用,这对当时动物学者的思维有巨大的影响,体腔的存在被解释和用作基本的形态、系统和演化属性。体腔被认为是识别更复杂的身体组织和更高的演化发展的标志。因此,在很长一段时间内,连续体腔的存在与否是动物学术语中真体节“true segments”和其他序列结构(serial structures,被称为假体节pseudometameres)区别的基本标准[24-25]。即有体腔动物的分节被认为是真分节,无体腔动物的分节被认为是假分节。
绦虫链体的节被称为节片(proglottid)[26];动吻动物躯干部的体节则被称为节带(zonite);扁形动物和环节动物无性繁殖产生的个体被称为个员“zooid”[27-28]。事实上,无论是绦虫的节片还是动吻动物的节带,实际都是动物的体节。而无性繁殖所产生的个员不属于动物的分节现象,只能算作是假分节(图1)。
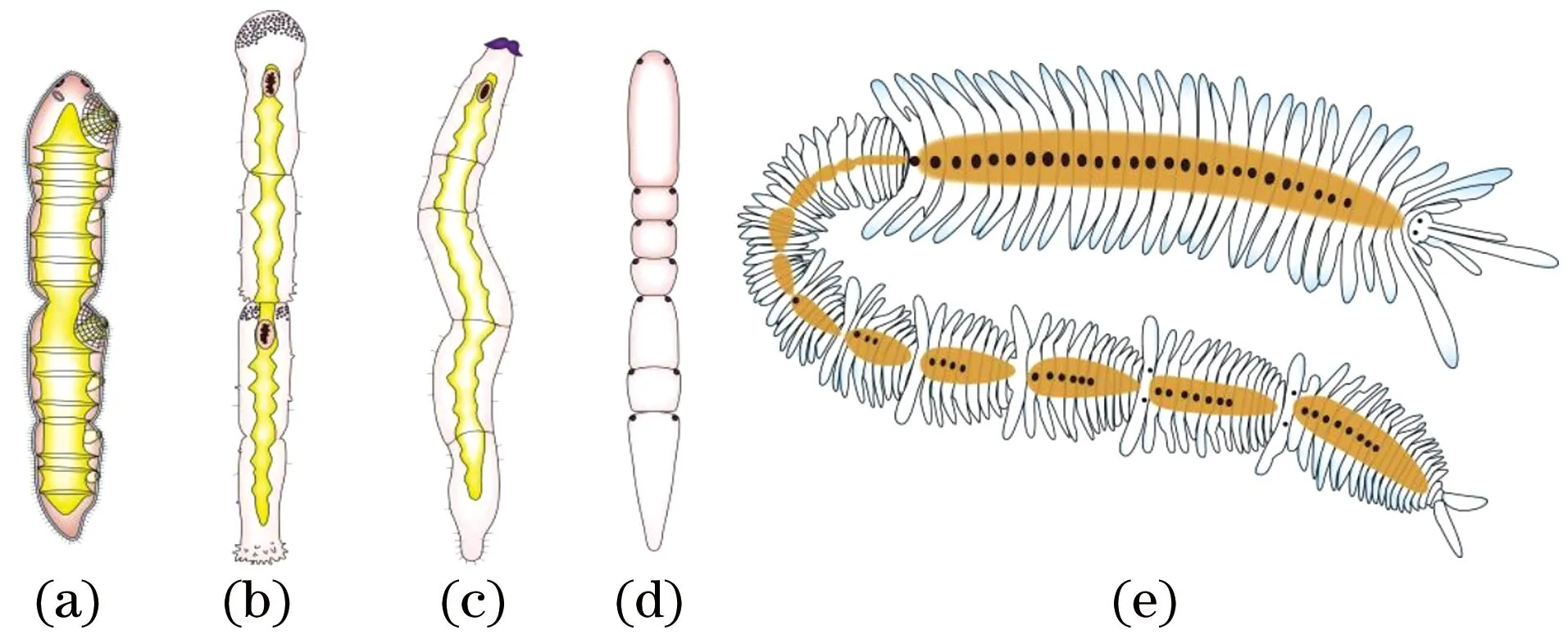
(a)~(d)扁形动物涡虫;(a)微口涡虫(Microstomum)的出芽过程;(b)红斑微口涡虫(M. rubromaculatum)的异分体(paratomy)繁殖;(c)乳头微口涡虫(M. papillosum)的原分(architomy)繁殖; (d)直口涡虫(Stenostomum)的虫链。(e)环节动物多毛纲沙蚕目裂虫科多链虫属多链虫(Myrianida pachycera)的无性繁殖。(a)~(d)根据文献[28]修改;(e)根据照片绘图。
3 分节的起源
分节的起源在不同动物的个体发育中有明显的区别,主要表现有3种起源方式:后生长区起源、前生长区起源及轴向同时起源。
3.1 后生长区起源(origin of the posterior growth zone)
此类分节现象可能起源于担轮幼虫样祖先,经原始的分节或不分节的两侧对称动物(Urbilateria)[29-31]向原口和后口动物方向发展(图2)。环节动物、脊索动物及大部分节肢动物的分节均为后生长区起源,即这些动物均是从靠近身体末端的后生长区向前长出体节。
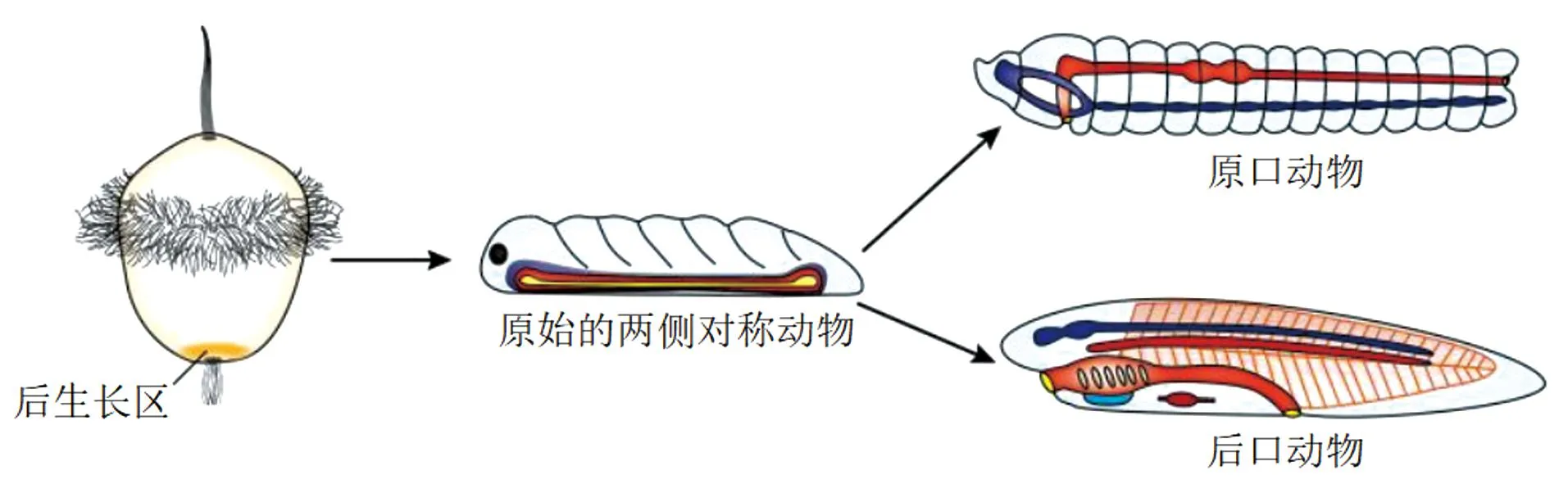
图2 分节的后生长区起源示意(根据文献[6]修改)
3.2 前生长区起源(origin of the anterior growth zone)
前生长区起源以扁形动物多节绦虫亚纲动物为例,其链体是从其头节后的颈区向后生长形成的。近颈区新形成的节片尚未分化形成生殖器官等构造,为未成节,未成节经发育分化出生殖器官等构造原基,再进一步形成成熟的生殖器官便成为成节,成节繁殖后生殖器官等构造退化,节片被充满虫卵的子宫填充成为孕节(图3)。

图3 分节的前生长区起源示意(根据文献[8]修改)
3.3 轴向同时起源(axial simultaneous origin)
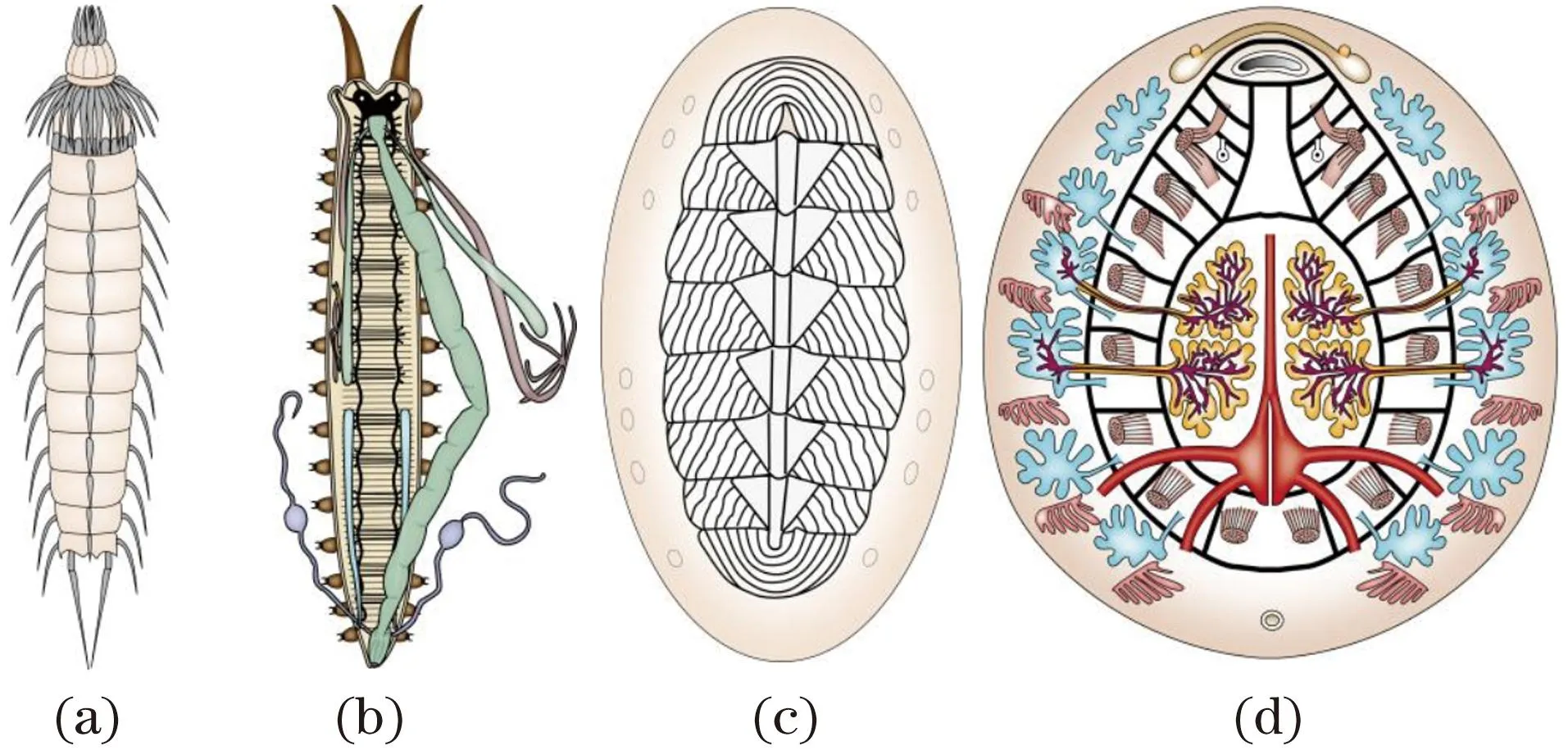
分节的轴向同时(同步)起源是指体节是同时形成的,没有先后顺序。长胚带胚胎发育昆虫,如果蝇便是实例。这类分节的轴向同时起源可能由低等蠕虫的系列重复现象逐渐演变形成。它们的消化、生殖、神经等内脏器官成对按体节重复排列,如三肠目涡虫[图4(a)]和纽虫[图4(b)]等,当动物体左右蠕动时,各器官之间的体壁处可能产生褶缝,随后在前后褶缝间分化出肌肉群,最终形成体节。
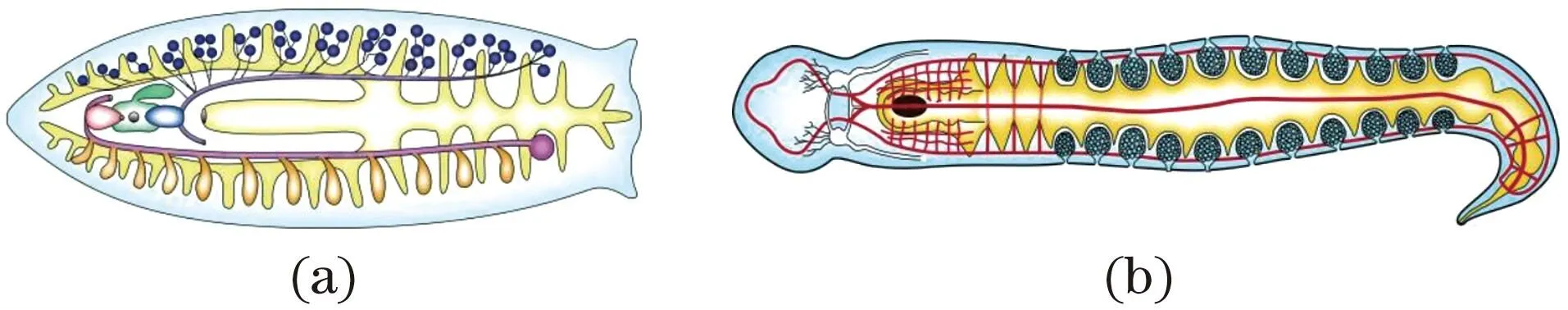
系列重复的三肠目涡虫(a)和纽虫(b)内部结构示意。(a)根据文献[28]修改;(b)根据文献[33]修改。
4 分节的类型
文献中分节的类型不统一,为了不使问题复杂化,我们在此使用同律分节、异律分节与系列重复概念。
4.1 同律分节
同律分节(图5)指相同规律的分节现象,指大部分体节在形态结构和功能上基本一致的分节现象,以环节动物多毛纲的砂蚕为典型代表。砂蚕非繁殖期的大部分体节为同律分节,其每节均有体腔室、神经节及其分支、后肾管、侧血管及成对的附肢(疣足)等构造。动物学教材环节动物门内容的概述以蚯蚓为代表动物,而蚯蚓实际上没有典型的后肾管(大肾管),而是具有小肾管(咽头小肾管、隔膜小肾管和体壁小肾管)结构,同时蚯蚓的生殖腺集中分布于身体前端的部分体节内。同律分节主要涉及中胚层和外胚层衍生结构的分节。
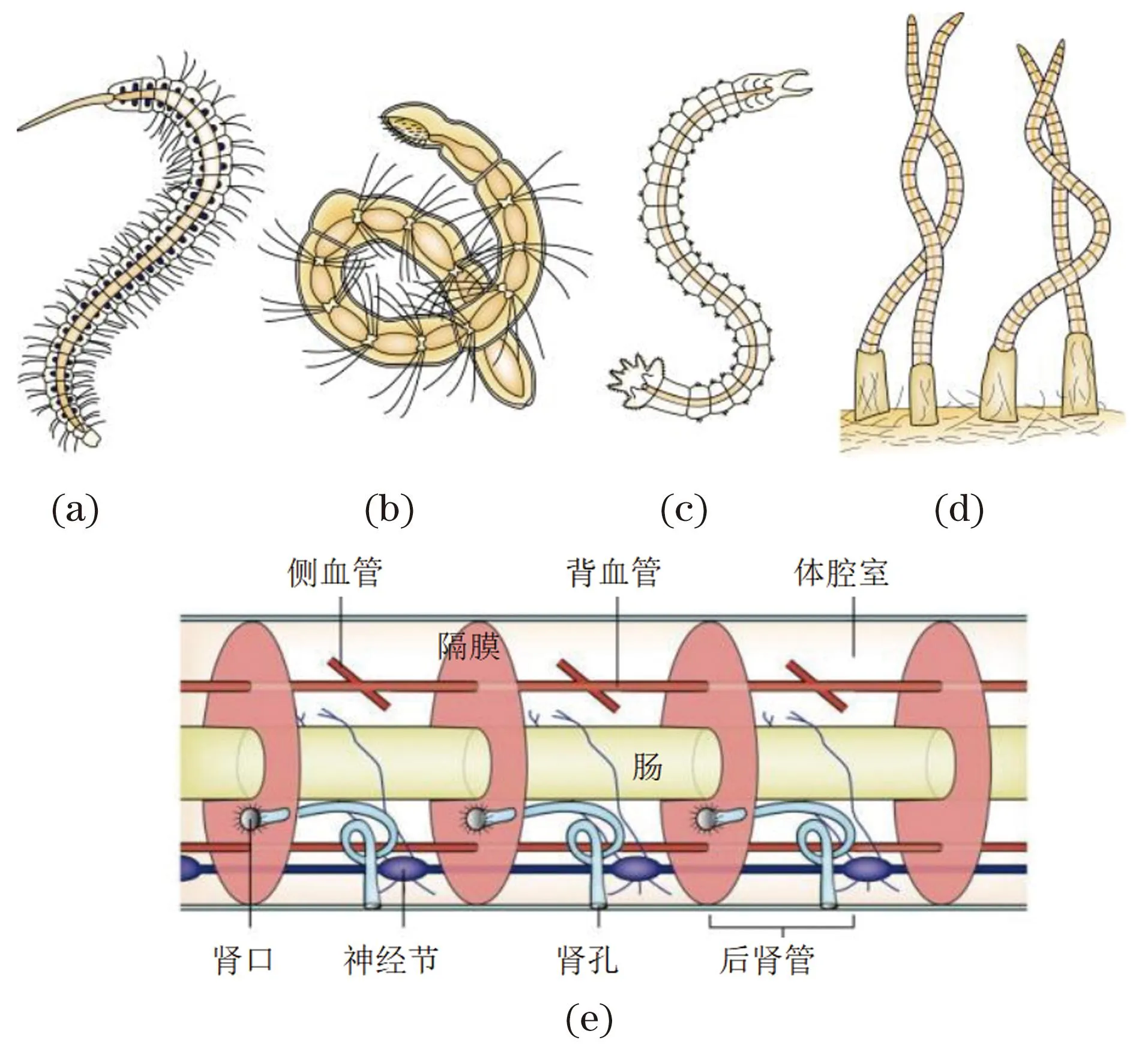
(a)杆吻虫(Stylaria sp.);(b)顠体虫(Aeolosoma sp.);(c)尾盘虫(Dero sp.); (d)水丝蚓(Tubifex sp.);(e)砂蚕体节结构示意。
4.2 异律分节
异律分节指不同规律的分节现象,指体节在形态结构和功能上不同(有分化)的分节现象,在异律分节基础上,功能相近或相关联的体节组合到一起形成动物身体的结构和功能明显不同的单位,称为分部(tagmata),如虾和蟹等的头胸部和腹部[图6(a)],蝗虫[图6(b)]等昆虫的头部、胸部和腹部等。虾和蟹等的头胸部主要具有感觉、摄食和运动功能,腹部主要具有营养代谢和繁殖等功能,由于游泳足的存在,腹部亦具有一定的运动功能。蝗虫等昆虫的头部主要具有感觉和摄食功能,胸部主要具有运动功能,腹部主要具有营养代谢和繁殖功能。节肢动物的分节主要涉及外胚层衍生结构的分节。

图6 沼虾(a)和蝗虫(b)在异律分节基础上的分部(引自文献[8])
异律分节除了主要存在于节肢动物中,微颚动物[图7(a)]、须腕动物[图7(b)]、半索动物[图7(c)]和环节动物[图8(a)]中亦有存在。
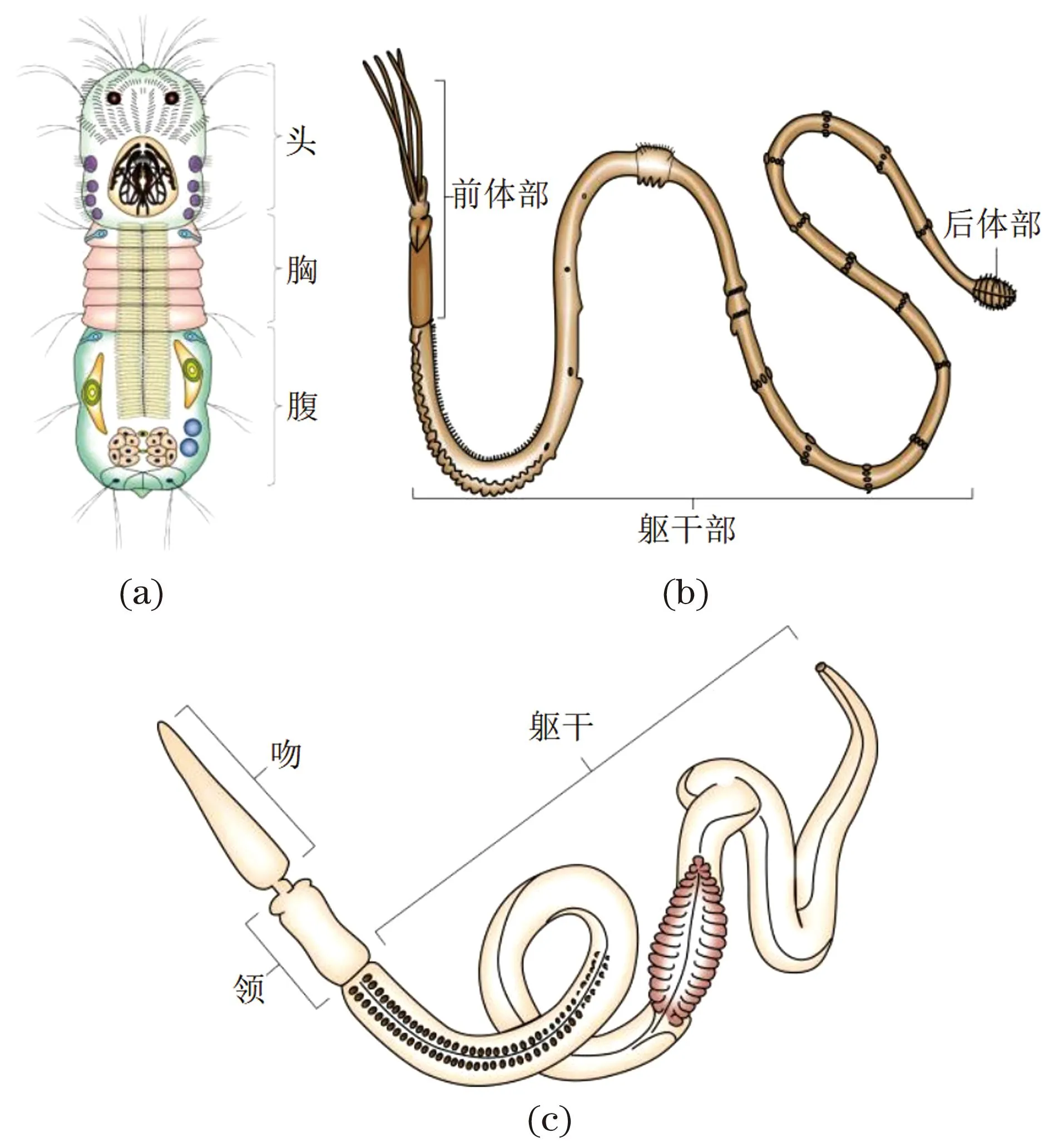
(a) 微颚动物; (b)须腕动物; (c)半索动物柱头虫。
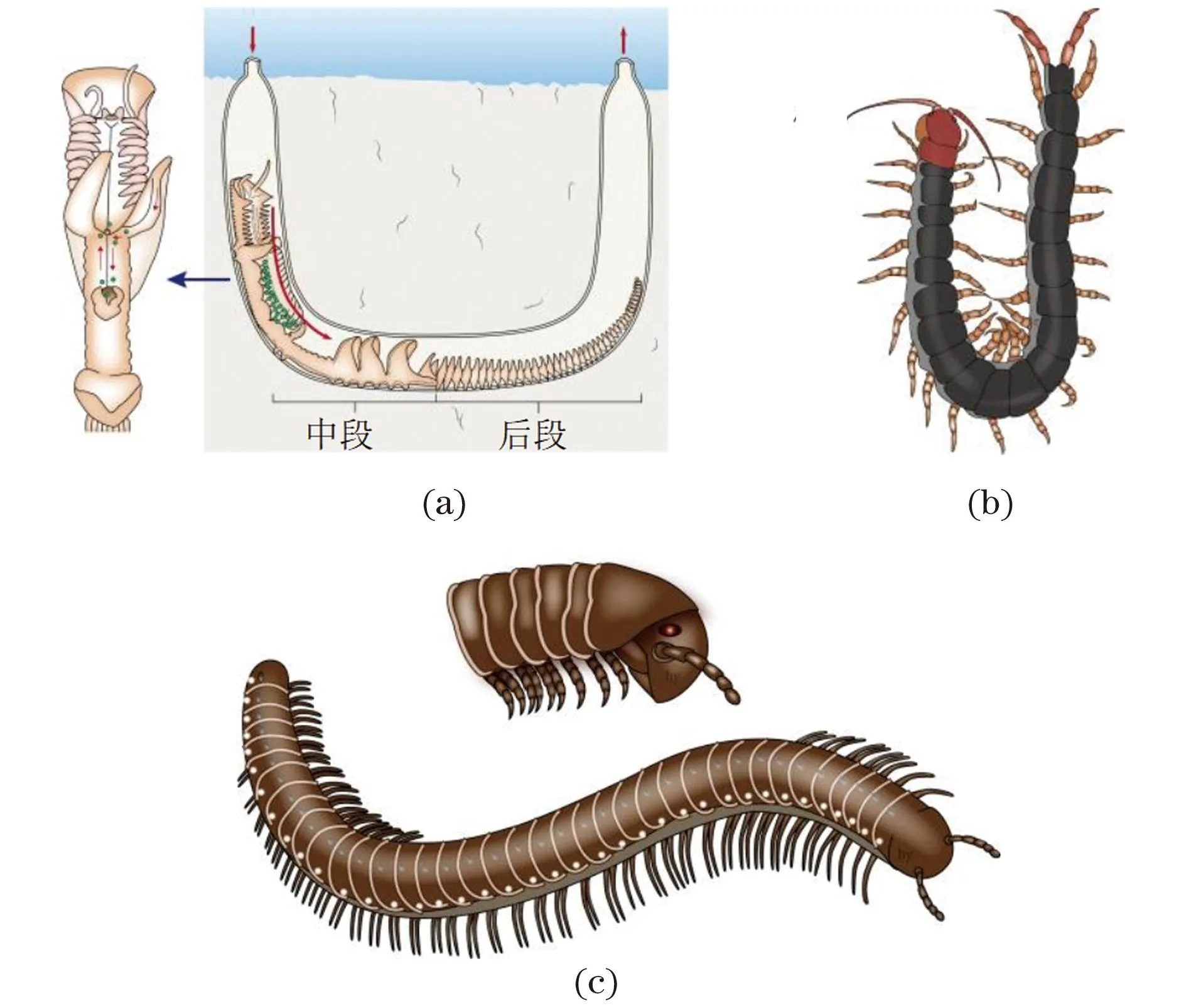
(a)毛翼虫; (b)蜈蚣; (c)马陆。
环节动物中存在异律分节的物种如隐居目的种类,如毛翼虫科(Chaetopteridae)的毛翼虫(磷沙蚕)(Chaetopterussp.)[图8(a)]。毛翼虫身体明显区段化:前段由11节组成,各节愈合,由两侧退化的圆锥状的疣足突起可辨识,前段以感觉和掘穴功能为主;中段由5节组成,其第1节(相当于第12体节)的疣足特别发达,变为翼状体,并分泌一个长长的黏液网,一直延伸到中段扇状疣足前方的小食物杯里,第2节(相当于第13体节)较细长,有食物杯,含许多腺体,用以团聚食物,第3~5节(相当于第14~16体节)的疣足变为扇状结构,以扇动水流完成摄食功能。后段较长,由13~60个体节所组成。疣足相似,背叶发达,为伸向两侧的圆锥状突起,腹叶退化为2个小突起。后段主要完成营养代谢和生殖功能。
节肢动物中的多足亚门动物身体分为头部和躯干部[图8(b)和(c)],其躯干部由同型的体节组成,应当归入同律分节的范畴。
其他应归入同律分节范畴的动物如:扁形动物多节绦虫纲的动物(图3)、动吻动物[图9(a)]、有爪动物[图9(b)]、软体动物多板纲[图9(c)]和单板纲[图9(d)]动物及脊索动物部分组织器官的分节等。
(a) 动吻动物外形; (b)有爪动物内部结构; (c)多板纲动物石鳖背面观; (d)单板纲动物新碟贝内部结构。(a)和(b)根据文献[33]修改;(c)和(d)根据文献[32]修改。
动吻动物[图9(a)]及有爪动物如栉蚕[图9(b)]的躯干部外胚层与中胚层组织存在明显的同律分节。软体动物多板纲石鳖的8块背板[图9(c)]为外胚层组织的分泌物,与外胚层组织的节段性有关,与脑区域化基因相关[34];梯状的神经系统、成对的鳃则为外胚层组织的分节。单板纲新碟贝成对的鳃及神经系统亦为外胚层组织的分节,肾管、肌肉则为中胚层组织的分节[图9(d)]。脊索动物的分节主要表现为肌肉、脊椎骨的分节,肋骨的按节排布,鳃裂孔的成对排布等,其中,肌肉、脊椎骨的分节和肋骨的按节排布涉及中胚层组织的分节,而鳃裂孔的成对排布则涉及3个胚层组织的分节。
因此,同律分节及异律分节普遍存在于动物界众多类群而非局限于环节动物、节肢动物及脊索动物或脊椎动物。
4.3 系列重复
系列重复指动物体的有些系统出现规则重复现象。系列重复可以涉及内胚层组织的重复,如扁形动物三肠目涡虫[图4(a)]和纽虫[图4(b)]等,它们体表没有呈现出明显的分节,但其内部身体纵轴向的消化道常分出规则成对的肠盲囊,生殖腺亦在体两侧成对规则排布,神经系统及纽虫循环系统的血管均呈现为规则重复。
此外,线虫具有与F-肌动蛋白分布相一致的角质环(系列凝结物环)及连续排列的运动神经元;线形动物幼虫具有环状角质层结构;鳃曳动物具有环状角质层结构和相应的环状肌肉,一系列均匀间隔的环状神经束;棘头虫躯干的环状收缩和相应的环状肌肉及一系列环状通道;缓步动物胚体中的躯干节段和中胚层囊,成对的附肢和少数相应的重复的肌肉组和躯干神经节等一系列结构;棘皮动物腕上的骨板和管足的规则排布等均可以看作是系列重复[27]。
5 动物分节的演化意义
分节现象源于原始的三胚层、两侧对称动物,是动物演化过程中的一个重要标志。分节有其美学意义,分节的规律性,被称为“平移对称”,成为关键的美学标准之一。分节不仅增强了动物的运动机能,也是生理分工的开始。使动物体向更高级形式发展。动物定向运动,逐渐分化出头部、胸部、腹部或头胸部、腹部或头部、躯干部等体制。运动机能的加强,必然伴随摄食的增加、新陈代谢的强化,相应出现了排泄器官系统等的分化。因此,分节现象是动物发展的基础,是高等无脊索动物的标志,其对促进动物体的新陈代谢,增强动物对环境的适应能力等均有重要的意义。
6 结论
传统上仅把环节动物、节肢动物和脊索动物或脊椎动物当成分节的动物,把其他动物当作不分节或假分节的动物是不全面的。动物的分节现象并不仅是教材中介绍的同律分节与异律分节那么简单。系列体腔不能作为分节的关键元素。分节现象广泛存在,不同类群的动物分节涉及不同的胚层、也可能是不同的区域。这是动物界长期演化适应的结果。
猜你喜欢
大自然探索(2024年1期)2024-03-19 19:01:03
快乐作文(1.2年级)(2022年8期)2022-05-30 10:48:04
小学科学(学生版)(2021年12期)2021-12-31 03:22:22
家园·建筑与设计(2021年3期)2021-12-30 21:16:45
建材发展导向(2021年15期)2021-11-05 08:21:20
科学大众(中学)(2021年3期)2021-05-17 17:07:01
小学科学(学生版)(2020年7期)2020-07-28 08:00:54
疯狂英语·初中天地(2019年4期)2019-10-17 02:07:12
课外生活(小学1-3年级)(2018年10期)2018-11-19 07:26:10
中国漫画(2017年4期)2017-06-30 13:06:16
