
多酶一锅法合成硫酸基供体3′-磷酸腺苷-5′-磷酸硫酸(PAPS)
2023-12-11 03:04:06岳继恒储梦萍储建林何冰芳
生物加工过程 2023年6期
岳继恒,储梦萍,储建林,3,何冰芳,3
(1.南京工业大学 生物与制药工程学院,江苏 南京 211800; 2.宜兴市人民医院,江苏 宜兴 214200;3.南京工业大学 药学院,江苏 南京 211800)
硫酸化修饰是生物体内糖类和小分子等化合物的一种常见修饰方法,硫酸化寡糖在动物、植物、海洋生物以及人体的生长发育等方面发挥重要的作用[1-2]。以低分子肝素、肝素类似物、硫酸软骨素和硫酸皮肤素等为代表的硫酸化寡糖不仅能够与细胞表面和细胞外基质(ECM)内的相关因子(包括细胞表面受体、ECM蛋白、酶、生长因子(GFS)、细胞因子和形态因子等[3])相互作用,还可以与细胞核中的组蛋白和组蛋白修饰酶或转录因子等结合,在生物体中发挥不同的生物功能[4]。
为了研究硫酸化修饰寡糖类药物在生物体内的作用,往往需要高纯度、高度硫酸化修饰的硫酸寡糖类产物为研究基础。目前,硫酸寡糖的制备方法主要有3种:超低分子硫酸肝素可利用水解动物源大分子肝素[5]、全化学合成肝素寡糖[6]和生物工程合成肝素前体再进行硫酸化修饰[7]。硫酸化修饰生物工程合成肝素前体获得的肝素寡糖产物具有混合物少、结构较为明晰、硫酸化位点可控等优点,且质量较容易控制,已成为研究热点[8-10]。然而,在肝素前体的酶法硫酸化修饰的过程中,需要用到大量硫酸基供体,3′-磷酸腺苷5′-磷酸硫酸(PAPS)是该反应常用的硫酸基供体[11-13]。
PAPS为高能磷酸化合物,非常不稳定[14],同时,市售PAPS的价格昂贵。因此,生物酶法合成PAPS一定程度可解决这种困境。目前,酶法制备PAPS主要有2种方式。第1种是使用芳基硫磺基转移酶Ⅳ(AST Ⅳ),以对硝基苯酚硫酸盐(PNPS)为硫酸基供体,将化合物3′,5′-二磷酸腺苷(PAP)转化为PAPS[15-16]。由于硫酸基转移反应会生成产物PAP,PAP又作为AST Ⅳ的反应底物,这种方式可以实现PAPS的循环再生[17-18]。第2种方法是以三磷酸腺苷(ATP)和无机硫酸盐等为原料,在ATP硫酸化酶(ATPS)的催化作用下生成5′-腺苷磷酰硫酸(APS),进一步由APS激酶(APSK)催化APS与ATP发生磷酸化反应,生成PAPS[15,19]。
本研究以ATP和无机硫酸盐为原料,建立多酶催化反应体系制备PAPS,对多酶催化反应涉及的ATPS和 APSK进行优化表达。同时,在反应体系中加入铜绿假单胞菌(Pseudomonasaeruginosa)来源的无机焦磷酸化酶PPase,将多酶催化合成PAPS过程中的副产物焦磷酸(PPi)水解为磷酸(Pi),以利于PAPS的生成与累积,显著提高反应体系中PAPS的产量水平。
1 材料与方法
1.1 实验材料
1.1.1 质粒与菌株
大肠杆菌EscherichiacoliBL21(DE3)和E.coliDH5α、质粒pET-28a(+)和pET-28a-sumo等保存于何冰芳教授实验室。密码子优化后来源于乳酸克鲁维酵母(Kluyveromyceslactis)的ATPS基因和来源于产黄青霉菌(Penicilliumchrysogenum)的APSK基因由金唯智生物科技有限公司(苏州)合成并克隆至pMD18-T载体。
1.1.2 主要试剂
胰蛋白胨和酵母粉,OXOID公司(英国);限制性核酸内切酶、T4 DNA连接酶、Taq酶等,TaKaRa Bio宝日医生物技术(北京)有限公司;小批量质粒提取试剂盒和凝胶回收试剂盒,AXYGEN公司;ATP等其他化学试剂均为市售国产分析纯试剂。
LB培养基:胰蛋白胨10 g/L、酵母粉5 g/L、NaCl 10 g/L;pH 7.0,121 ℃灭菌 20 min,加入过滤除菌的硫酸卡那霉素(Kana)使其终质量浓度为50 mg/L。
固体培养基是在LB培养基中加入琼脂20 g/L,相同灭菌条件。
1.2 实验方法
1.2.1 重组酶ATPS、APSK和PPase的表达载体构建方法
乳酸克鲁维酵母来源的ATPS基因(GenBank登录号:CR382125.1),根据大肠杆菌的密码子偏好性,利用https:∥www.novopro.cn/tools/codon-optimization.html网站进行密码子优化,由金唯智生物科技有限公司(苏州)进行全基因合成。以引物(F′-CCCGGATCCGCATGCCGAGCCCGCAT (BamH Ⅰ)、R′-GATCAAGCTTCTTCCAGTTCCTCGAG (HindⅢ))为模板进行PCR,产物和载体质粒使用限制性内切酶BamH Ⅰ和HindⅢ进行双酶切,再用T4 DNA连接酶连接,将目的基因构建到含有sumo标签蛋白的pET-28a-sumo载体上。将重组质粒导入E.coliBL21(DE3)感受态细胞得到重组菌株E.coliBL21(DE3)/pET-28a-sumo-ATPS。
根据大肠杆菌的密码子偏好性优化产黄青霉菌来源的酶APSK的基因(GenBank登录号:8313362),并由金唯智公司进行全基因合成获得目的基因。以引物(F′-CGCGGATCCA ̄T ̄G ̄T ̄C ̄T ̄A ̄C ̄C ̄A ̄A ̄C ̄A ̄T ̄C ̄A ̄C ̄C ̄T ̄T ̄C ̄C ̄A (BamH I)和R′-CCCAAGCTTT ̄T ̄A ̄C ̄T ̄C ̄C ̄T ̄T ̄C ̄T ̄T ̄G ̄G ̄C ̄A ̄G ̄G ̄C ̄A (HindⅢ))进行PCR,限制性内切酶BamH I和HindⅢ进行酶切和T4 DNA连接酶连接,将目的基因构建到pET-28a(+)载体上。将重组质粒导入E.coliBL21(DE3)感受态细胞,得到重组菌株E.coliBL21(DE3)/pET-28a-APSK。
提取何冰芳教授实验室保存的菌株铜绿假单胞菌的基因组DNA。根据NCBI数据库中铜绿假单胞菌来源的无机焦磷酸化酶PPase的氨基酸序列和基因序列,克隆获得酶PPase的基因。以引物(F′-GCGGATCCCATATGGGTCTGGA (BamH Ⅰ)和R′-CCAAGCTTGGATCCTTATTTTTTGG (HindⅢ))进行PCR,以上述同样方式构建到pET-28a(+) 载体上。将重组质粒导入E.coliBL21(DE3) 感受态细胞中,得到重组菌E.coliBL21(DE3)/pET-28a-PPase。
1.2.2 重组酶ATPS、APSK和PPase的诱导表达条件优化方法
将E.coliBL21(DE3)/pET-28a-sumo-ATPS、E.coliBL21(DE3)/pET-28a-APSK和E.coliBL21(DE3)/pET-28a-PPase在含有50 μg/mL卡那霉素的LB平板上复苏2次。分别将3个重组菌接种于含50 μg/mL卡那霉素的LB液体培养基中,于37 ℃、180 r/min摇床培养12 h。以体积分数2%的接种量转接至含有培养基的250 mL三角瓶中,重组菌在相同条件下培养至OD600为0.6~0.8时加入诱导剂进行诱导表达。分别设置不同的诱导温度、诱导剂种类和浓度以及诱导后培养时间来考察不同的培养条件对重组酶的表达影响。以诱导剂IPTG终浓度为0.1 mmol/L,分别考察不同诱导温度(16、20、25和30 ℃)对重组酶的表达影响,培养条件为180 r/min和24 h。发酵液于8 000 r/min离心20 min获得菌体,以Tris-HCl(pH 8.0)重新悬浮菌体,超声破碎(超声破碎细胞的功率50%,每次超声3 s,间隙5 s;超声总时间为20 min)获得含重组酶的溶液;再于12 000 r/min的条件下离心20 min获得含重组酶的上清液和沉淀部分。含重组酶的上清液和沉淀部分进行SDS-PAGE凝胶电泳分析,使用Image J软件对蛋白质的SDS-PAGE电泳分析,采集图像进行灰度扫描,计算目标酶的蛋白相对含量。在优化后的诱导温度下,分别加入不同种类诱导剂(IPTG、乳糖)和不同浓度的诱导剂IPTG(0.05~0.4 mmol/L),培养24 h,再分析获得较优的诱导剂种类和浓度。在最佳诱导温度和诱导剂的条件下,重组菌分别在诱导后培养不同时间(2、3、6、8、10、12、24和36 h)取样,计算目标酶的蛋白相对含量。
1.2.3 重组酶ATPS、APSK和PPase的纯化与酶学性质研究
收集菌体超声破碎后的上清液,重组酶ATPS、APSK和PPase(C端带有6×His标签)分别使用Ni-NTA柱纯化。重组酶纯化时,先在中压柱(φ1 cm×25 cm)中装入5 mL填料,再以10倍柱体积量的buffer A(20 mmol/L Tris-HCl (pH 8.0)、0.5 mmol/L NaCl、20 mmol/L咪唑)平衡,加样量为5 mL。在buffer A中加入体积分数20%的buffer B(20 mmol/L Tris-HCl (pH 8.0),0.5 mmol/L的NaCl、250 mmol/L的咪唑),先洗脱杂蛋白,再以体积分数100%的buffer B洗脱目的蛋白,收集洗脱液。纯化后的酶蛋白经透析、超滤后,置于含体积分数20%甘油的Tris-HCl(20 mmol/L,pH 8.0)缓冲溶液中保存。
分别考察pH、反应温度和Mg2+浓度对重组酶ATPS和PPase的酶活影响。分别将2种酶置于不同pH的缓冲体系(pH分别为3.0、4.0、5.0、6.0、7.0、8.0、9.0和10.0)中,测定相应的酶活。在pH 8.0的条件下,分别将2种酶置于不同的温度(25、30、35、37、40、45、50和60 ℃),测定相应的酶活。在最适pH和温度的条件下,在反应体系中加入不同终浓度的MgCl2(0.5、1、2、4、6、8和10 mmol/L),测定相应的酶活。
1.2.4 重组酶PPase的酶活测定方法
重组酶PPase的酶活测定使用钼酸铵显色法[14,20],重组酶PPase可以与PPi反应,生成磷酸(Pi),具体反应见式(1)。
(1)
Pi可以与钼酸铵反应生成能稳定显色的磷钼蓝,符合朗伯-比尔定律。在酸性条件下,正磷酸盐与钼酸铵反应生成黄色的磷钼杂多酸,具体反应见式(2)。
(2)
在室温条件下,磷钼杂多酸可以立即被柠檬酸或者氯化亚锡还原,转化成稳定的蓝色络合物——磷钼蓝(H3PO4·10MoO3·Mo2O5或H3PO4·8MoO3·2Mo2O5),以此显色反应测定重组酶PPase的酶活。
重组酶PPase酶活测定的反应体系(总体积500 μL):Tris-HCl(100 mmol/L,pH 8)、MgCl2(2 mmol/L)和PPi (2 mmol/L);加入重组酶PPase溶液 10 μL。置于35 ℃反应10 min后,加入AAM溶液(2.5 mmol/L的(NH4)6Mo7O24·4H2O,0.1 mol/L的H2SO4,50%丙酮)4 mL,混合均匀后再加入0.5 mL柠檬酸溶液(1.0 mol/L),再次混匀并静置30 s后测定405 nm处的吸光度。配制H3PO4浓度分别为0.2、0.4、0.6、0.8、1.0、1.5、2.0和2.5 mmol/L的磷酸盐溶液,依次加入AAM溶液和柠檬酸溶液。测定吸光度,制作标准曲线(拟合曲线:y=0.440 8x+0.017 9,R2=0.999 2),计算Pi的浓度。酶活定义1 U为在上述条件下每分钟催化PPi生成1 μmol的Pi所需要的酶量。
1.2.5 重组酶ATPS的酶活测定方法
通过PPi的生成量来测定重组酶ATPS的酶活,以酶PPase催化产生Pi的量来计算PPi的生成量[14]。配制反应体系总体积为1 mL的溶液:Tris-HCl (100 mmol/L,pH 8.0)、MgCl2(6 mmol/L)、Na2MoO4·2H2O (5 mmol/L)和ATP (2 mmol/L),加入重组酶ATPS溶液10 μL;置于37 ℃水浴中反应15 min后,沸水浴10 min终止反应。冷却至室温并稀释2倍,再向终止的反应液中加入1 U 的PPase酶溶液,置于37 ℃水浴反应10 min。取500 μL反应液,测定405 nm处的吸光度并根据标准曲线计算Pi的浓度。酶活定义(1 U)为在上述反应条件下每分钟催化ATP生成1 μmoL的PPi所需要的酶量。
1.2.6 多酶催化合成PAPS的反应条件优化方法
多酶催化合成PAPS的反应体系(1 mL):Tris-HCl (0.1 mol/L)、LiCl (0.05 mol/L)、Na2SO4(0.1 mol/L)、MgCl2(4 mmol/L)、ATP (20 mmol/L),加入重组酶ATPS (10 U)、PPase (10 U)和APSK (10 μL,蛋白质量浓度19.60 mg/mL)。35 ℃条件下反应一定时间后,沸水浴10 min终止反应,再以12 000 r/min离心10 min,并稀释和过滤后测定PAPS产物。单因素实验优化:考察不同的底物浓度(5~35 mmol/L)、温度(25~50 ℃)、pH(4~10)、反应时间(10~105 min)等条件对多酶催化反应合成PAPS产率的影响,并研究批次补料方式对多酶催化反应过程中产物PAPS累积的影响。
1.2.7 产物PAPS的检测与分析方法
采用高效液相色谱法对产物PAPS进行分析[3]。使用C18色谱柱(Agilent,ZORBAX SB-Aq 4.6 mm×250 mm,5 μm,填料粒径12 nm),流动相为体积分数5%甲醇/甲酸铵溶液(50 mmol/L,甲酸调节pH至4.5),流速为1.0 mL/min,进样量为10 μL,检测波长为UV 254 nm。进一步利用液质联用色谱仪(LC-MS)对产物进行分析,负离子模式,扫描范围为100~3 000m/z。
2 结果与讨论
2.1 酶ATPS、APSK和PPase的重组质粒构建结果
分别构建了酶APSK、ATPS和PPase的表达载体,将重组质粒分别导入E.coliBL21(DE3)感受态细胞,菌落PCR和基因测序验证了结果(图1);成功获得了重组菌株E.coliBL21(DE3)/pET-28a-sumo-ATPS、E.coliBL21(DE3)/pET-28a-APSK和E.coliBL21(DE3)/pET-28a-PPase。
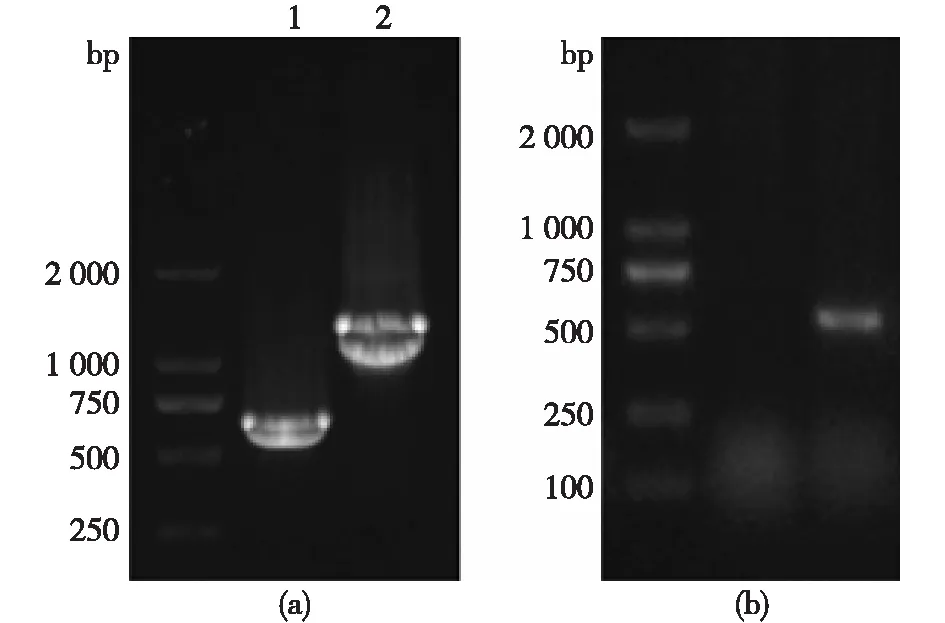
图1 APSK、ATPS和PPase 3种酶基因片段的凝胶电泳验证
图1为3种酶基因片段的电泳检测结果。图1(a)中的条带1为APSK,条带2为ATPS,图1(b)为PPase的扩增条带。3组条带分别在1 509、647和537 bp处有对应条带,与预期结果相符。同时基因测序的结果与目的基因相同,证明重组表达质粒构建成功。
2.2 重组酶ATPS、APSK和PPase的诱导表达条件优化
重组菌株在摇瓶中培养后,可实现3个酶(ATPS、APSK和PPase)在大肠杆菌中的表达。图2显示了重组菌E.coliBL21(DE3)/pET-28a-sumo-ATPS诱导表达酶ATPS的优化结果。由图2可知:大肠杆菌表达sumo标签融合的酶ATPS的最佳诱导温度为25 ℃,最适的诱导剂IPTG的终浓度为0.3 mmol/L。在摇瓶培养温度为25 ℃和培养基中添加0.3 mmol/L 的IPTG条件下连续培养36 h,考察不同的诱导后培养时间对重组酶ATPS的表达影响。诱导后培养12 h内,重组酶ATPS的累积明显;随后,重组蛋白的含量增加缓慢。在诱导后培养24 h时,重组酶ATPS的蛋白含量水平达到最大值。

图2 温度、诱导剂和诱导时间对酶ATPS的表达影响
图3显示了重组菌E.coliBL21(DE3)/pET-28a-APSK表达酶APSK的优化结果。由图3可知:在培养温度为16、20和25 ℃的条件下,重组酶APSK的可溶性部分相对较高,表达的可溶性蛋白含量的差异较小;当培养温度为30 ℃时,可溶性蛋白含量显著降低。因此,选择25 ℃为大肠杆菌表达重组酶APSK的最佳诱导温度。
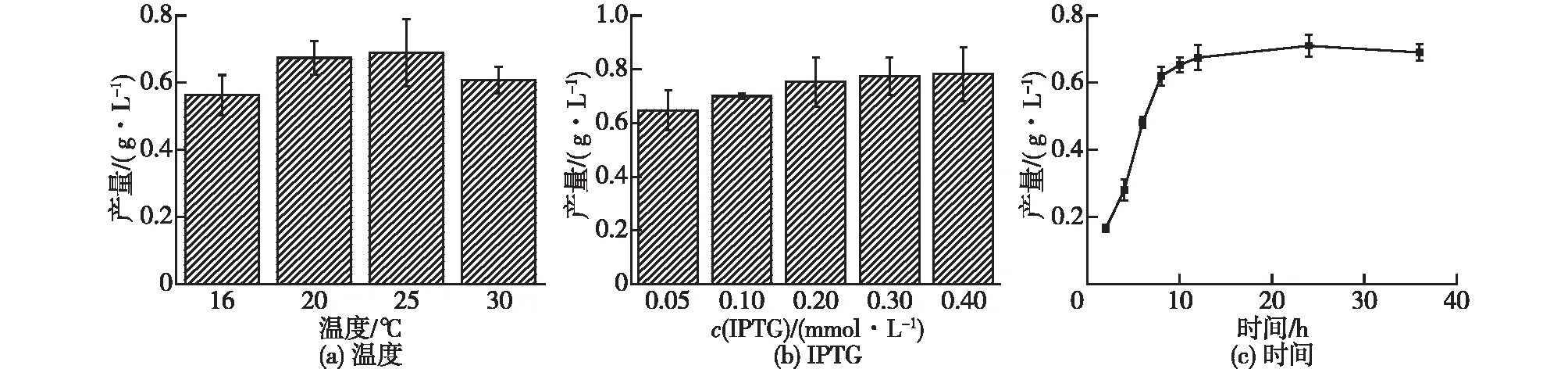
图3 温度、诱导剂和诱导时间对酶APSK的表达影响
添加诱导剂IPTG的终浓度为0.1 mmol/L时,表达的重组酶APSK的蛋白量最高。当诱导剂IPTG的终浓度大于0.1 mmol/L时,酶蛋白的表达量略有降低,且可溶性蛋白的比例也明显降低。因此,选择IPTG终浓度为0.1 mmol/L为最佳的诱导剂浓度。
在培养温度为25 ℃和诱导剂IPTG的终浓度为0.1 mmol/L的条件下,培养24 h,重组酶APSK的表达量最高。
图4为重组菌E.coliBL21(DE3)/pET-28a-PPase表达酶PPase的优化结果。由图4可知:当诱导温度为20 ℃时,酶PPase的表达水平较高,实现了重组酶PPase在大肠杆菌中的高效可溶性表达;当在培养基中添加不同种类的诱导剂(乳糖、IPTG)时,以诱导剂IPTG对重组酶PPase的表达效果最佳;加入阿拉伯糖反而会降低PPase的表达量。因此,培养基中添加0.1 mmol/L的IPTG为最佳的诱导剂浓度。在培养温度为25 ℃和诱导剂IPTG的终浓度为0.1 mmol/L的条件下,诱导后培养12 h,重组酶PPase的表达水平最高。

图4 温度、诱导剂和诱导时间对酶PPase的表达影响
2.3 酶ATPS、APSK和PPase的纯化结果
图5为ATPS、APSK和PPase的表达及纯化结果。由图5可知:使用带sumo标签的质粒pET-28a-sumo构建了酶ATPS的表达载体,融合表达策略显著提高了重组酶ATPS的异源表达水平,并且表达的可溶性蛋白占总蛋白的相对含量较高。在大肠杆菌中,表达的重组酶APSK大部分为可溶性蛋白,仅有少量的包涵体。在大肠杆菌中,也实现了重组酶PPase的高效可溶性表达。重组酶APSK、ATPS和PPase的可溶性表达水平较高,3种酶的异源表达效果较好。

图5 重组酶APSK、ATPS和PPase的表达和纯化的SDS-PAGE电泳分析
同时发现,纯化后的酶APSK、ATPS和PPase条带较为单一,纯度较高。进一步以BCA试剂盒测定了纯化后的酶ATPS、APSK和PPase的蛋白质量浓度,分别是9.47、19.60和16.33 mg/mL。使用钼酸铵法测定了纯化后的酶PPase和ATPS的酶活,计算得到酶PPase和ATPS的比酶活分别是518.72和 12.75 U/mg。同样地,测定了摇瓶中发酵产酶PPase和ATPS的酶活,酶PPase和ATPS的酶活分别是124 492.70 和 1 811.78 U/L。摇瓶发酵液中目标重组酶的蛋白水平为酶PPase 1.225 g/L、酶ATPS 0.752 g/L和酶APSK 1.470 g/L。
2.4 重组酶PPase和ATPS的酶学性质研究
2.4.1 温度、pH和Mg2+浓度对重组酶PPase酶活的影响
考察不同的温度、pH和Mg2+浓度对铜绿假单胞菌来源的酶PPase酶活的影响,结果见图6。由图6可知:当缓冲液pH为8.0时,酶PPase的酶活最高;当pH为3.0~7.0和pH>8.0时,酶活有所降低。当温度为25~40 ℃时,酶PPase具有相对较高的酶活。当温度为35 ℃时,酶PPase的酶活最高。因此,重组酶PPase的最适温度为35 ℃,最适pH为8.0。当溶液中的Mg2+浓度为1~4 mmol/L时,酶PPase具有相对较高的酶活(相对酶活>70%)。添加2 mmol/L Mg2+时,酶PPase的酶活最高。
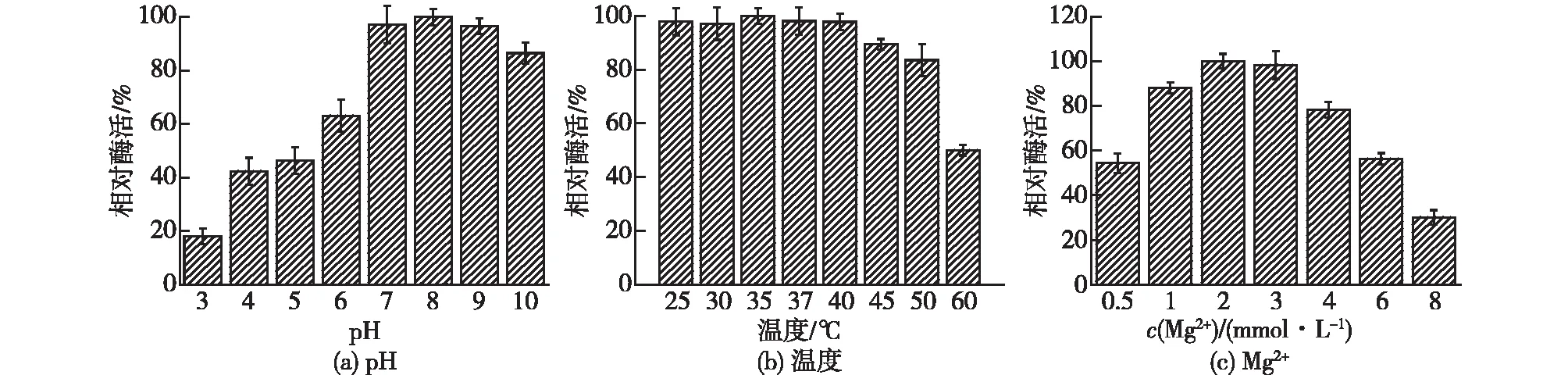
图6 温度、pH和Mg2+浓度对重组酶PPase的酶活影响
2.4.2 温度、pH和Mg2+浓度对重组酶ATPS的影响
考察不同的温度、pH和Mg2+浓度对重组酶ATPS酶活的影响,结果见图7。由图7可知:当pH为8.0时,sumo标签融合表达的酶ATPS的酶活最大;当pH<8.0 和pH>8.0时,重组酶ATPS的酶活降低明显。当温度为30~45 ℃时,融合表达的酶ATPS具有较高的酶活;当温度为35 ℃时,融合表达的酶ATPS的酶活力最大。当Mg2+浓度为2~8 mmol/L时,融合表达的酶ATPS具有相对较高的酶活性(相对酶活>80%)。因此,融合表达的酶ATPS的最适温度为35 ℃,最适pH为8.0。Mg2+浓度为4 mmol/L时,融合表达的酶ATPS大约具有90%的相对酶活; Mg2+浓度为6 mmol/L,融合表达的酶ATPS酶活最高。
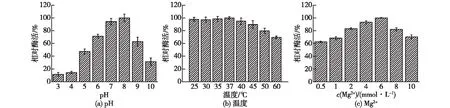
图7 温度、pH和Mg2+浓度对重组酶ATPS的酶活影响
2.5 多酶催化反应合成PAPS的条件优化
2.5.1 多酶催化反应合成PAPS产物的HPLC和质谱结果
以大肠杆菌表达的重组酶APSK、PPase和sumo标签融合表达的酶ATPS建立多酶催化反应体系,对反应后的样品进行HPLC检测,分析是否有PAPS产物的生成,结果如图8(a)所示。图8(a)HPLC分析表明,在1.34 min左右出现一个新的尖峰,通过与文献[14]比较底物ATP的保留时间,推测可能是PAPS的产物峰。进一步以LC-MS检测和验证,结果如图8(b)所示。图8(b)质谱分析显示:在m/z=505.115 8[M-H]-处出现离子峰,与计算得到的PAPS的分子量506.0相一致[3],可以说明该产物为PAPS。
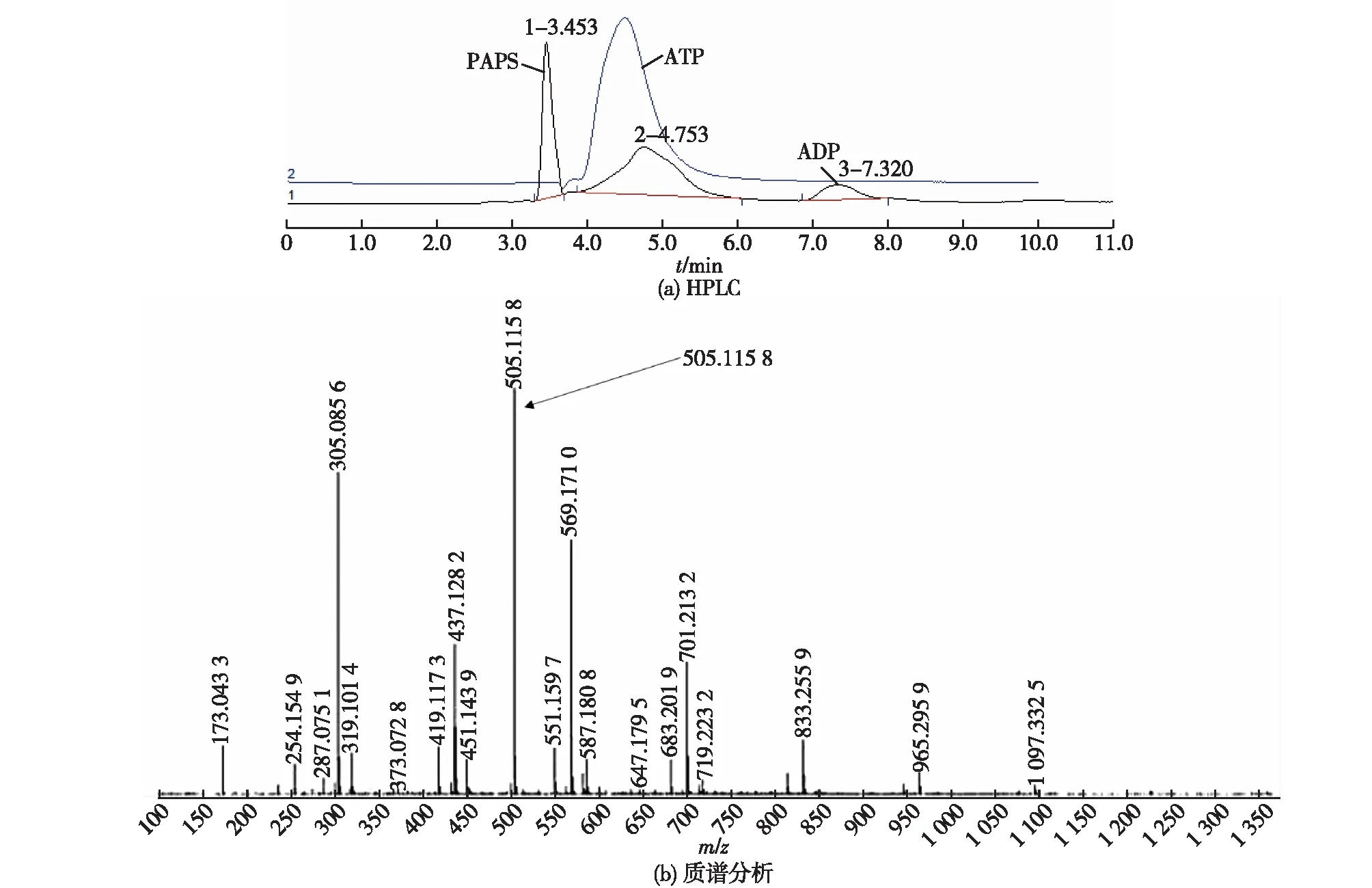
图8 产物PAPS的HPLC和质谱分析图
2.5.2 多酶催化反应合成PAPS的条件优化结果
以ATP(初始浓度20 mmol/L)、Na2SO4为底物,重组酶APSK和融合酶ATPS能催化合成PAPS。在反应体系中额外加入酶PPase,可以使得酶法合成PAPS过程中产生的中间产物PPi水解为Pi,从而有利于驱动反应向合成PAPS进行[19]。酶法合成的PAPS的反应条件优化结果如图9所示。由图9(a)可知:与不加酶PPase组相比,在重组酶APSK和ATPS催化合成PAPS的反应体系中加入酶PPase,PAPS的累积量从4.79 mmol/L增加到6.85 mmol/L,明显提升了PAPS产物的浓度水平。因此,在酶APSK和ATPS催化合成PAPS的反应体系中加入铜绿假单胞菌来源的无机焦磷酸水解酶PPase可以对产物PAPS的累积起到明显的促进作用。

图9 酶法合成的PAPS的反应条件优化结果
不同pH对多酶催化反应的转化率影响显著。当pH为8.0时,多酶催化反应的转化率最高。反应温度从25 ℃升高到35 ℃,多酶催化反应的转化率逐渐升高;在35 ℃时,达到最大。随着温度的升高(>35 ℃),转化效率迅速降低,并不利于PAPS的合成。酶学性质研究也表明,在温度为35 ℃时,融合酶ATPS和酶PPase具有相对最高的酶活性。另外,融合酶ATPS和酶PPase最适的pH均为8.0。在多酶催化反应体系中,不同的酶具有相同或相近最适pH和最适温度能增强多酶催化反应的协调性,可能更加有利于多酶催化反应合成PAPS产物。因此,优化后的多酶催化反应条件为反应温度35 ℃和pH 8.0。
当底物ATP的浓度为20 mmol/L时,转化率最高。当ATP浓度升高为25~35 mmol/L,转化率明显降低,这可能是过高浓度的底物ATP抑制了酶的活性,导致多酶催化反应的转化率下降[19]。
2.5.3 多酶催化合成PAPS的反应进程曲线
在ATP初始浓度为20 mmol/L、pH 8.0和反应温度为35 ℃的条件下,酶法合成PAPS的反应进程曲线如图10所示。

图10 酶法合成PAPS的反应进程曲线
由图10可知:酶APSK、融合酶ATPS和酶PPase构成的多酶催化反应体系中生成的产物PAPS浓度水平增加明显;当反应时间为60 min时,反应体系中的PAPS产物达到峰值,产物PAPS的浓度为8.33 mmol/L。继续延长反应时间,反应体系中PAPS并不增加,反而略有降低,可能是PAPS不稳定所导致的。
2.6 分批补料原料ATP对多酶催化反应合成PAPS的影响
在多酶催化反应体系中,较高浓度的底物ATP(>20 mmol/L)会显著抑制PAPS的合成。在反应过程中,以分批补料方式添加ATP原料,不仅可以维持体系中一定的ATP浓度,还能一定程度地解除过高浓度的底物对酶活性的抑制作用[19],以进一步提高多酶催化反应合成PAPS的产量水平。在总体积为100 mL的反应体系中,ATP初始浓度为20 mmol/L,考察分批补料底物ATP对多酶催化反应合成PAPS产物的累积影响,结果如图11所示。反应过程中,分别在反应时间为1、3和 6 h时添加适量的底物ATP至20 mmol/L。由图11可知:经3次补料后,反应体系中产物PAPS的积累量依然在持续增加。在反应时间为7 h(第3次补料1 h后)时,反应体系中获得的产物PAPS积累量达到最大(28.60 mmol/L),相当于产物PAPS的质量浓度水平(14.47 g/L)。
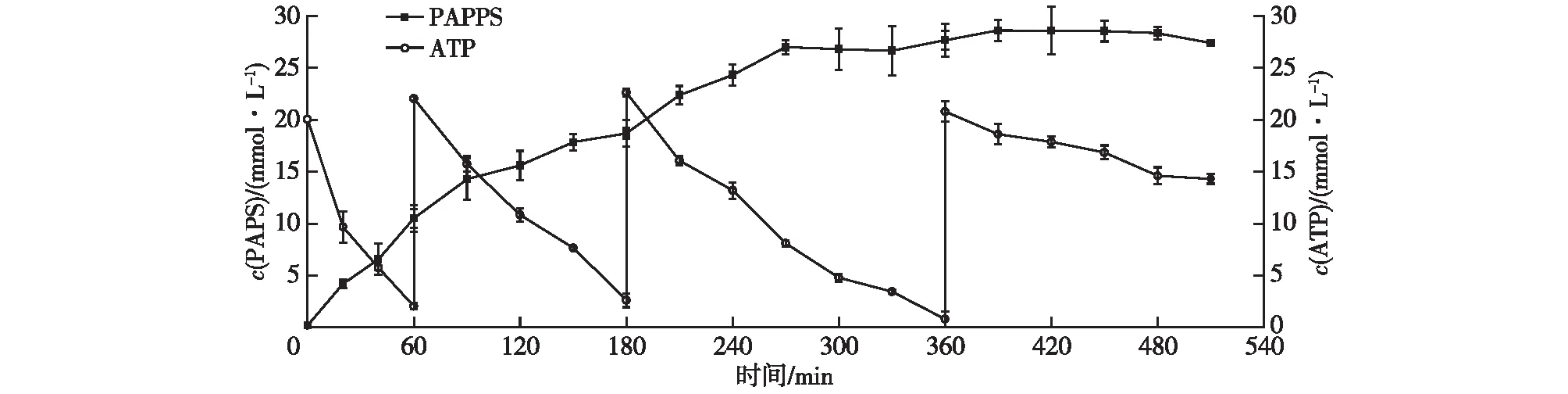
图11 分批补料ATP的对酶法合成PAPS的累积影响
3 结论
采用密码子优化后的ATP硫酸化酶和APS激酶的基因,并利用sumo标签融合表达ATP硫酸化酶ATPS,显著提高了重组酶ATPS和APSK在大肠杆菌中的异源表达水平。克隆并表达了铜绿假单胞菌来源的无机焦磷酸水解酶PPase。优化诱导表达条件实现了重组酶ATPS、APSK和PPase在大肠杆菌中高效可溶性表达,摇瓶发酵液中产生的PPase为1.225 g/L、ATPS为0.752 g/L和APSK为1.470 g/L。对重组酶进行分离纯化并研究了酶ATPS和PPase的酶学特征,发现融合表达的酶ATPS与酶PPase具有相同的最适pH和最适温度。
以融合表达的酶ATPS、重组酶APSK和酶PPase建立多酶催化反应体系,HPLC和LC-MS验证了PAPS产物的生成。多酶催化反应体系中加入铜绿假单胞菌来源的无机焦磷酸水解酶PPase消耗中间副产物,有效地驱动反应进行,对产物PAPS的累积起到了明显的促进作用。融合表达的酶ATPS与酶PPase具有相同或相近最适pH和最适温度,能增强多酶催化反应合成PAPS的协调性,更加有利于多酶催化反应高效合成PAPS产物。多酶催化反应中,批次补料反应不仅可以维持体系中一定的ATP浓度,还能一定程度地解除过高浓度的底物对酶活性的抑制作用,显著提升了多酶催化反应体系中累积PAPS的产量水平,最终使PAPS的累积量达到28.60 mmol/L(14.47 g/L)。
多酶催化反应合成PAPS可通过较低的成本获得PAPS,为后续研究提供了极大的便利。多酶催化反应合成PAPS仍有很多需要改进和探索的方面,比如PAPS的分离纯化以及更高效地利用酶,使得PAPS的制备更加简便高效,适用范围更加广泛,生产成本更加低廉。
猜你喜欢
特产研究(2024年1期)2024-03-12 05:40:56
江苏农业科学(2022年2期)2022-02-15 01:02:02
矿产综合利用(2020年1期)2020-07-24 08:51:20
中国化肥信息(2020年2期)2020-01-20 07:53:15
天然产物研究与开发(2019年10期)2019-11-05 10:12:44
中国化肥信息(2019年4期)2019-01-17 18:47:06
江西农业大学学报(2018年5期)2018-11-22 09:21:00
中学生理科应试(2017年2期)2017-04-01 00:05:46
上海医药(2016年23期)2016-12-22 15:21:07
现代农业(2016年6期)2016-02-28 18:42:51
