
喀斯特地区植物对石生高钙环境的生态适应研究进展
2023-11-23 01:25:12田玲李双智陈凯金兆麟牟凤娟
南方林业科学 2023年5期
田玲,李双智,陈凯,金兆麟,牟凤娟
(西南林业大学·林学院,云南 昆明 650224)
喀斯特地貌又称石灰岩地貌或岩溶地貌[1],全球陆地喀斯特地貌面积约2.2×107km2,占全球陆地总面积的15%[2]。世界喀斯特地貌集中分布于欧洲(阿尔卑斯山、法国中央高原、俄罗斯乌拉尔山地)、美洲(美国肯塔基州、印第安纳州以及古巴)、亚洲(中国西南部和越南、老挝北部)、大洋洲(澳大利亚南部)等地区[3],在我国的分布面积居世界首位。喀斯特地貌在中国的总面积达3.44×106km²,约占国土面积的1/3,其中广西、贵州、云南等省(区)是最集中也是最大的分布区[4]。石灰岩主要成分是碳酸钙,极易被溶蚀,常形成喀斯特地形,该地区的土壤层薄、钙含量高、保水能力差,生态系统敏感而脆弱,不合理的人类活动更是加剧了本地区的土地退化[5]。在石灰岩为主的地区,木本植物的覆盖度高、植被活动强,但生产力水平较低,该生境若遭遇破坏,30%的特色植物种类首先丧失,且恢复极为困难。
植物在受外界环境影响过程中,会逐渐演化形成各式各样的形态结构和生理结构以适应环境变化,植物特征与环境之间相互作用的关系历来备受关注[6]。植物的功能性状受非生物环境的影响较大,喀斯特地区土壤、水分及营养物质的异质性常促使植物功能性状产生异变[7]。近年来,大量研究发现植物形态与其生境间存在强烈的内在联系[8]。植物可通过调节营养器官的生物量来应对环境的变化,在形态上主要表现为根、茎、叶的相应变化。植物根系和生境的关系是密不可分的,常通过其大小、长度、根毛等变化以适应环境[9];植物茎的大小、茎密度、茎粗和茎高等形态特征也会为了适应环境变化而产生相应变化[10];叶片在应对环境变化时,其数量、大小、厚度、毛被等常发生相应变化[11]。分布在喀斯特生境下的植物常具有喜钙、耐旱以及石生性等特点,代表植物所属科有苦苣苔科(Gesneriaceae)、秋海棠科(Begoniaceae)、槭树科(Aceraceae)、樟科(Lauraceae)、胡桃科(Juglandaceae)等,而芸香科(Rutaceae)的拟九里香属(Bergera)植物仅分布于南方喀斯特地区[12],具有较强的耐旱性。
研究喀斯特地区植物适应环境的形态特征对于认识和保护其脆弱的生态系统,揭示植物与环境之间相互作用的关系,以及合理保护与持续开发利用喀斯特地区植物资源具有重要意义。本文对喀斯特地区植物形态对高钙和石质生境适应方面的相关研究进行了总结,以期为喀斯特地区的植物和生态保护及生态恢复提供参考。
1 石质生境下植物的生态适应策略
喀斯特石漠化(Karst stony desertification)指喀斯特地区由于人类活动破坏自然植被导致土壤流失、岩石裸露的土地退化现象,被称为“地球的癌症”[13]。由此可见,石漠化已成为喀斯特地区生物生存的“大敌”,一直是备受各界关注的重要环境问题[14]。由于土壤稀少、岩石裸露,碳酸盐岩长期经化学侵蚀,石灰岩地区常形成包括地上浅土层及地下岩溶裂隙和洞穴的双层结构[15]。且浅薄土层保存水分的能力有限,降水常从石缝渗入到地下岩溶系统中[16],因此干旱是该地区的典型特征;此外土层不连续以及大面积裸露的基岩等也导致石灰岩地区的土壤、水分、营养物质等具有极强的空间异质性[17],这种空间分布特征极大地限制了植物种群的分布和生长。喀斯特地区优势植物对独特的石生小生境具有明显的适应性。近年来石灰岩生态研究的重点偏向于对地衣、苔藓等低等植物的研究[13,18-19],而对其他类型植被的研究较少。
1.1 植物根对石质生境的生态适应
植物主要通过根系从土壤中吸收水分和养分并固定植物体,对水分的吸收能力主要取决于根系的形态和生理特征,其形态的改变与生境含水量直接相关。岩石深部的溶蚀裂隙可为木本植物根系发育提供更大的空间,以获取深层地下水[20]。在这种生长环境下,裂隙中会截留一部分土壤资源,裂隙水也是近地表层植物生长的重要水源之一,这些水分不易蒸发。在这种土壤浅薄的石生环境中,植物往往具有发达、强壮的根和茎,根系可以爬过岩石,穿透裂缝,并从裂缝中生长到较深的地方获得水分和营养[21],如茂兰喀斯特山地上很多榕属(Ficus)植物根部常会沿石缝或岩壁延伸至数十米深处[22]。一些优势木本植物主要通过发达根系吸收土壤水分或者伸入较深的岩石孔隙中汲取水分,如火棘(Pyracantha fortuneana)、齿叶黄皮(Clausena dunniana)、香叶树(Lindera communis)等[23],而黄连木(Pistacia chinensis)、乌桕(Sapium sebiferum)最长的根可达29 m[24]。因此植物根的形态受裂隙类型及裂隙水含量影响较大,生长在裸露岩石上的植物根部在连接的裂缝中生长,不但可以在根部储存营养物质和水分,还能通过发达的根部堵塞和拦截降水[25],说明裂隙对植物根系的生长有一定的促进作用;贵州喀斯特地区植物根系常沿着岩石和土壤的交界面弯曲或螺旋式生长,在有限的土壤条件中通过弯曲缠绕岩石的方式生长可以获取更多的水分和营养物质[26],这是喀斯特地区植物的重要生存策略。如任豆(Zenia insignis)的侧根系粗壮发达,穿透力强,可沿着岩石表面、岩石间隙和石缝中生长,根可长达数米[27]。此外大量研究表明还有一些植物根系可以和丛枝菌根真菌(Arbuscular mycorrhizal fungi,AMF)建立一种互利共生的关系[28],有利于植物扩大根系、促进对营养物质的吸收,以适应石灰岩生态系统养分缺乏的环境。
调查发现很多喀斯特地区的优势种一般以灌木和草本植物为主,这可能与它们自身的习性和生理形态特征有关[29]。灌木深根系的特征使其在贫瘠和空间异质性极强的喀斯特地区具有较强的适应能力,如紫穗槐(Amorpha fruticosa)根系会伸长寻找更多空间,而胡枝子(Lespedeza bicolor)根系会横向生长尽量占据有限空间,这两种植物都会通过增加根系生物量来适应石质生境[30]。而浅根系草本植物通常更矮小,对水分需求也更小,主要利用表层带水和浅层土壤水,具有极强的适应能力;1 年生草本植物虽然寿命短但繁殖能力强[31],广泛传播的种子使其后代可以占据更多的环境资源。在同一群落中植物的根系结构和分布差异在水分吸收和再分配中具有重要作用,这些深根系和浅根系的植物在生态位上形成一种互补关系,可以充分利用有限资源缓解种间竞争,从而可以维持自然生态系统中的物种多样性和群落稳定性(表1)。
1.2 植物茎对石质生境的生态适应
植物茎是根和叶之间起支持作用的重要营养器官,可通过木质部和韧皮部传输水分和营养物质。早期有学者指出,相比较于根而言,茎受环境影响的表现更为显著[32]。Liu 等[33]研究发现常分布在喀斯特地区岩石缝上的藤本植物地果(Ficus tikoua),其匍匐茎上会产生不定根扎入岩石缝吸收土壤中的养分和水,茎叶则会蔓延到石质化的周边区域进行光合作用;Ziemmer 等[32]通过对野牡丹科(Melastomataceae)植物茎进行解剖,发现该科植物茎具有含水的髓腔,这与其节水策略有关。与其他生境相比,生于喀斯特悬崖上的尾囊草属(Urophysa)植物的根状茎更长、更粗壮,并深深扎根于岩石裂缝中,这有助于其汲取水分和养分[34]。此外有大量研究表明石灰岩地区植物茎的木质化现象普遍存在,如适生植物雏菊(Bellis perennis)的茎木质化,可有利于攀援和保持水分与营养[35]。大多数藤本植物的形态可塑性都有助于其在石质环境下寻找有利的生根点,以利用有限的资源[36],说明发达而强壮的茎、茎的木质化和茎的含水髓腔等也是植物适应石质生境的有效特征(表1)。
1.3 植物叶对石质生境的生态适应
植物叶片是对环境变化最为敏感的营养器官,其形态表现为多变[37],环境会通过影响植物叶片的形态功能从而影响其生长发育,这也是植物适应环境所形成的生存策略[38]。Xie 等[34]研究发现喀斯特悬崖上的尾囊草属植物叶片表皮具有明显的蜡质和大量表皮绒毛,而生长在森林或路边的个体的叶片均无表皮蜡质,表皮蜡具有阻止水分流失等作用,而表皮绒毛也能在防御和保护中发挥重要作用;同样有研究指出潺槁木姜子(Litsea glutinosa)的叶片性状在石面、土面、石沟、石窝和石缝等不同微地形条件下具有较为明显的分异特征,石缝中的潺槁木姜子比叶质量最大,叶片更厚[39]。植物通过增加叶片厚度来增加储水能力,从而减小叶面积并减少蒸腾,以提高植物对水分的利用效率[40]。总的来说,石质生境下的植物,尤其是干旱条件更为严重的山顶植物,其叶片形态表现为数量少、叶面积减小、革质叶或较厚的纸质叶、表皮有蜡质或绒毛、气孔指数降低,发达的表皮绒毛、栅栏组织[41],这些特征均有助于植物保存营养物质和水分(表1)。
植物在长期的适应进化过程中,可以通过调整自身的生理过程和结构来应对各种环境压力,植物倾向于将有限的资源分配到最受限制的器官,具体表现为生物量的增加,当环境胁迫程度加剧时,植物会倾向于增加根和叶生物量来降低茎的生物量[15]。根系是响应石质生境和岩溶裂隙的主要器官,其形态和特征可直接反映植物对环境资源的利用情况,在土壤异质性高的石灰岩地区,植物通过调整根系的空间分布与功能性状以适应生境[42]。发达的根系、灵活匍匐的茎、叶表皮具蜡质和绒毛等表型特征都可能有助于植物更好地适应石灰岩环境[43];植物通过不同的形态来适应生境,形成了对环境的协同进化,反映了植物对环境的不同适应策略(表1)。
2 高钙胁迫下植物的生态适应策略
Ca2+是植物细胞中的第二信使,可将环境中收到的信号传递给植物,使其产生相应的生理反应[44];钙也是植物必需的营养元素,在细胞壁和细胞膜的各种结构以及植物的生长发育调节中均具有重要作用。Ca2+通过质膜中的渗透通道进入植物细胞,其浓度变化是植物细胞内部对环境胁迫的一种协调反应[45];此外,钙还有利于植物对其他营养元素的吸收,能增强植物的抗逆能力,植物体的钙含量过高或过低均会抑制其生长甚至造成死亡。不同植物对钙的需求不同,导致他们对高钙环境具有不同的适应机制。石灰岩的主要化学成分是碳酸钙,钙含量高是喀斯特地区土壤的主要特征之一,也是影响当地植被类型、植物生理特征和植物分布的最重要环境因子之一[46],然而喀斯特地区植物适应高钙环境的机制尚不清楚。我国分布于石灰岩地区的植物多具有喜钙的特点,如该地区的先锋树种复羽叶栾树(Koelreuteria bipinnata)[47],喜钙植物紫花苜蓿(Medicago sativa)、甘草(Glycyrrhiza uralensis)等。
2.1 植物根对高钙胁迫的生态适应
喀斯特地区植物对钙的吸收能力主要受土壤钙含量的影响[48]。植物适应高钙环境最直接的方式是控制根系对钙的吸收和运输,根系从土壤溶液中获得的钙通过木质部转移到嫩枝及其他部位,因此植物根系的不同形态结构会直接影响植物对钙的吸收能力。喀斯特地区一些优势种在根组织中结合和固定过量吸收的钙,通过形成钙化根,限制钙素向上运输[49];另一些植物则会形成草酸钙晶体细胞来固定多余的Ca2+,如野生毛葡萄(Vitis quinquangularis)会在根中储存大量的草酸钙晶体[50];还有一些植物通过调节Ca2+在根质膜上的亲和力或Ca2+在根际细胞中的沉淀来限制根对Ca2+的吸收,高钙环境下会在根的皮层细胞中沉淀钙[51]。高钙环境中云贵鹅耳枥(Carpinus pubescens)的根细胞结构(如液泡、细胞壁)中会出现较多的钙质[52],这类植物均通过根部的特殊形态结构将过量钙储存在钙库而保持植物的正常生理活动。Chun 等[53]指出拟南芥(Arabidopsis thaliana)根系细胞可能通过细胞壁的增厚来适应高钙胁迫,认为根细胞壁的物理强化是该物种长期适应胁迫的重要原因,这可能是因为细胞壁是最早感受到外界环境刺激的细胞结构并且可以决定细胞的大小和形状,形成一种保护屏障。土壤中含有高浓度Ca2+时会显著抑制初生根的生长,高钙胁迫下圆叶乌桕(Sapium rotundifolium)总根长和根表面积会明显下降[1],可能是由于Ca2+可通过植物根部的钙离子通道直接进入表皮细胞和根毛,植物根部为适应高钙环境主动减弱根系生长以减少对土壤中Ca2+的吸收。总之,喀斯特地区植物通常会主动减少根系生长或形成特殊形态的根来适应高钙环境,此外植物还会通过改变根部的分泌物来影响相关的微生物群落,通过调节营养物质的吸收和代谢来适应高钙环境。
2.2 植物茎对高钙胁迫的生态适应
有关高钙胁迫下植物茎的形态变化的研究甚少,可能是植物茎适应高钙环境而产生的形态变化不如根和叶表现明显。研究指出当土壤中的Ca2+浓度发生变化时,对柠檬黄金花茶(Camellia limonia)茎中钙含量影响最大[54],其茎长、茎鲜重和茎干重均会显著降低[55];石华[56]也指出,在钙浓度较高时植物的茎秆干重会明显下降,这说明通过茎积累钙或降低茎生长以减少对Ca2+的吸收,是喀斯特地区植物对高钙环境的一种有效适应机制(表2)。
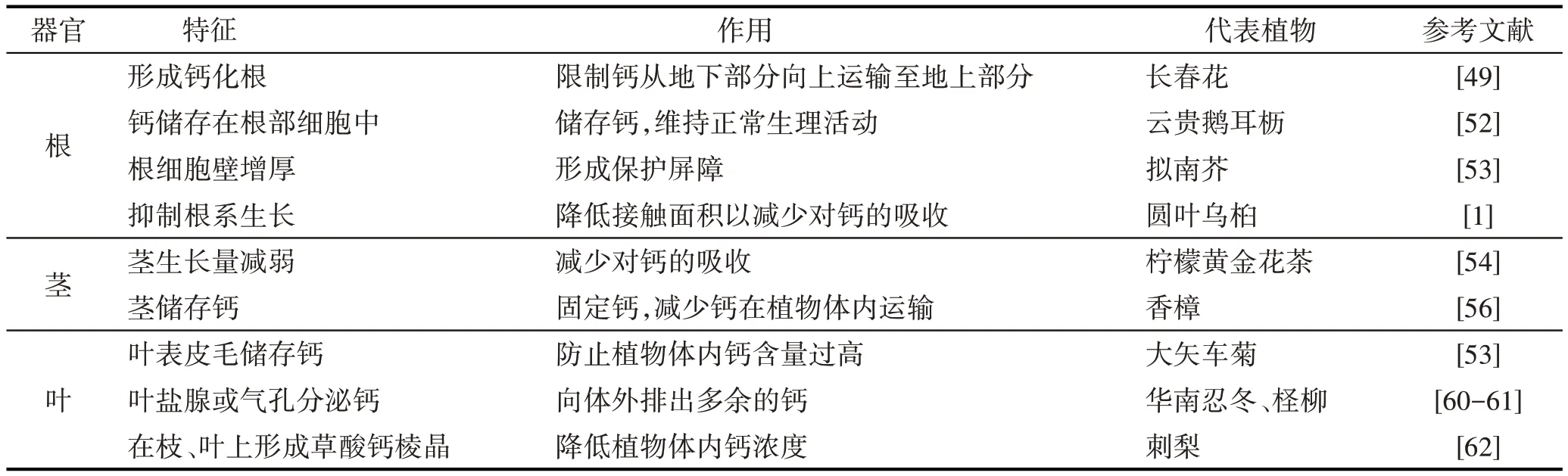
表2 喀斯特地区植物对高钙胁迫的生态适应特征表现Tab.2 The ecological adaptive characteristics of plants to high calcium stress in karst regions
2.3 植物叶对高钙胁迫的生态适应
叶是植物进行光合作用、气体交换和水分蒸腾的重要器官,喀斯特地区的优势种可以通过叶片的特殊结构排出多余的钙或限制Ca2+的输送[57]。如Li等[58]指出叶片是植物积累钙的主要器官,海绵组织是叶片中钙的主要储存部位,如吊石苣苔(Lysionotus pauciflorus)通过减少气孔导度和降低蒸腾作用等机制来限制Ca2+从茎向叶片的运输,且有利于植物保持水分;曹建华等[59]研究发现在岩溶区植物叶片中钙的形态主要以果胶酸钙为主,植物可以将细胞质中的钙运送出细胞,并储存在细胞壁中,实现对高钙环境的适应,这些生理机制对植物适应喀斯特地区高钙环境具有重要作用。此外,一些喜钙植物对环境的适应表现为通过气孔分泌钙盐,如华南忍冬(Lonicera confusa)可通过气孔分泌钙盐,并在叶腺和毛状体中储存Ca2+[60];柽柳(Tamarix ramosissima)可通过叶盐腺排出叶片中多余的钙[61];大矢车菊(Centaureaamericana)叶片表皮绒毛可以作为钙库,从而减轻高钙对植物气孔的伤害[53];野生毛葡萄叶片中钙较多,有大量的草酸钙和果胶酸钙储存于体内,并且可通叶片中的过气孔排出草酸钙[50],喀斯特地区的野生刺梨(Rosa roxburghii)也具有Ca2+的生物矿化贮钙功能和通过叶片气孔排钙的特性[62]。说明植物可以通过在细胞内或在外质体中与有机阴离子结晶以化学方式隔离过量钙,这是其对高钙环境的特殊生理响应机制(表2)。
由此可见,植物对高钙环境的适应过程很复杂,喀斯特地区生境的异质性使得植物的适应机制也存在多样性,但喜钙植物对土壤中钙的吸收具有一定的调节能力,植物根的排斥机制和叶的分泌机制使植物能够在高钙环境中保持其内部钙含量的大致恒定,这都是其对高钙环境的特殊适应机制。钙在喜钙植物体内的分布特征为叶>茎>根[63],根据喀斯特地区植物适应高钙环境的策略将其分为3 种类型:(1)积累型,当土壤钙水平过高时,植物体较多的钙储存在钙库而不影响其自身的生长发育;(2)回流型,当植物体钙含量过高时,这类植物会通过根部或叶片盐腺渗出植物体内过量的钙以维持正常生长;(3)限制吸收型,这类植物通过根或茎的特殊结构来限制植物体对Ca2+的吸收,或限制钙从地下部分向地上部分传输,使植株钙含量保持在正常水平。
3 喀斯特植物生态适应研究展望
喀斯特地区土壤存在极大的空间异质性[64],该地区植物在生理和环境因素等综合作用下会形成特定的生态适应策略,不同程度的环境胁迫对植物的生长发育以及生态适应性的影响不同。在异质生境下植物可能会产生趋异适应,如增长或变粗的根、木质化或长毛的茎、变厚或变小的叶,往往具有不同的形态特征却有相似的功能来适应环境。因此对喀斯特地区不同程度的高钙胁迫以及石质生境下植物的生态适应策略进行深入研究具有极为重要的意义。对此提出以下6个方面的展望:
(1)除了根、茎、叶等营养器官外,花、果实、种子等繁殖器官对喀斯特高钙及石质生境的形态适应策略还有待深入研究;(2)石质生境除了土壤瘠薄,常伴随着干旱胁迫,目前多数研究均为室内控制实验,可加强对喀斯特地区代表植物的野外耐旱适应策略研究;(3)石灰岩地区的生态环境特性使得该地区的植物同时面临干旱胁迫、高钙胁迫与石质生境,有必要开展对其相互作用以及对植物的共同影响等研究;(4)喀斯特植物种群中不同优势种功能性状的联系与差异以及不同种或个体之间对营养物质吸收的相互影响;(5)在植物对高钙胁迫适应方面,更多研究倾向于研究植物生理过程的变化,而对于植物解剖结构的研究相对较弱,可加强在高钙环境下植物的微形态适应策略的研究;(6)加强植物在不同生长发育阶段对特殊生境的分子调控基础研究,以揭示植物不同生长发育阶段对不同环境的适应机制。
猜你喜欢
石材(2022年4期)2022-06-15 08:55:38
落叶果树(2021年6期)2021-02-12 01:29:10
阅读(低年级)(2020年8期)2020-11-06 06:23:19
石材(2020年8期)2020-10-28 07:53:12
乡村地理(2019年2期)2019-11-16 08:49:32
华人时刊(2019年23期)2019-05-21 03:31:28
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26 13:44:20
体育科技(2016年2期)2016-02-28 17:06:07
黑龙江史志(2015年13期)2015-12-07 06:01:10
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
