
山核桃丰产生理基础研究
2023-10-28 13:24丁之恩姚洪礼董书甲郭明蔚
安徽农业科学 2023年19期
丁之恩,姚洪礼,董书甲,郭明蔚,范 飞
(1.亳州学院药食同源功能食品亳州市重点实验室,安徽亳州 236800;2.安徽农业大学,安徽合肥 230036)
山核桃属胡桃科山核桃属落叶乔木,俗称小胡桃、小核桃。山核桃属约有21个种,我国现栽培的经济价值较高的有2个种,一个原产我国,分布于浙江、安徽等省的山核桃;另一个原产北美,在我国北京、河北、江浙、福建、四川、江西等地栽培的长山山核桃,又名美国薄壳山核桃[1-3],山核桃为国内单位面积产量最高的木本油料树种之一[4-5],净果仁含油率高达69.8%~74.0%,带壳出油率为28%,比油茶、油桐高数倍。山核油清澄透亮,色淡金黄胜似芝麻油,油味独特芳香,有抗菌、抗氧化、抗肿瘤的作用[6-7],同时有润肺滋补的功效,冷榨油具有降低血脂[8]、预防冠心病的功效。另外,山核桃果仁中的蛋白质含量高达18.3%[4],且容易加工成多种风味产品,其综合营养价值不亚于橄榄油[9],并且加工工艺对核桃油脂组成有较大影响[10]。山核桃外果皮烧灰后含碱率达20%~30%,其中碳酸钠含量达60%以上,可在化工、医药等领域使用。因此,发展山核桃产业、推广丰产技术,对促进山区经济发展具有重要意义。
山核桃属于深根树种,适宜凉爽湿润,弱光照,昼夜温差小的立地条件。山核桃不耐严寒、酷暑,花期和果实生长发育时期[11-12]对气候条件敏感,对立地条件要求苛刻。造林地的选择条件是年均温大于13 ℃;土壤要求深厚、肥沃,质地从砂壤到轻黏土之间,以石灰岩、紫砂岩、灰质岩、花岗岩上发育的土壤为好,pH 5~7。海拔200~900 m。坡向要求:海拔小于500 m,阴坡、半阴坡;海拔大于500 m,阳坡、半阳坡。孤立峰、迎风坡、山谷和积水处不适宜种植[13]。山核桃干物质中的90%~95%来源于光合作用所获得的物质,光合作用及其强度是山核桃生命活动中重要的生理指标,对产量、品质影响较大。针对美国薄壳山核桃的光合生理已有报道[14-18],但中国山核桃、大别山山核桃等鲜有针对光合生理特性所开展的研究。受山核桃树体特性及栽培环境条件等多种因素的影响,研究光合特性对山核桃的科学栽培管理措施、提高其产量和品质具有重要意义。
1 材料与方法
1.1 试验地概况研究区位于安徽省金寨县,在5月中旬开展工作。金寨县位于安徽省西部,大别山北麓,位于115°221′~116°111′E、31°061′~31°481′N,为鄂、豫、皖三省交界处,其中天堂寨海拔达1 729.1 m。金寨属北亚热带季风型气候,年平均气温15 ℃,年降水1 400~1 600 mm,年日照时数2 060 h,其中山区在1 800~1 850 h,无霜期180~190 d,相对湿度年平均在80%左右,山场土壤大多由变质岩形成,主体由花岗岩、片麻岩构成,土壤多是黄棕壤或棕壤,呈弱酸性,pH 5.5~6.5。全县境内既无严寒,也无酷暑,气候温和湿润,土壤肥沃且深厚,山高林密,森林覆盖率高达70%,其地理位置和气象因子为山核桃生长发育创造了得天独厚的条件。
1.2 试验材料供试材料为多年生山核桃成年树(15年及其以上树龄),选择自然分布的山核桃成片林,管理水平正常,生长结果良好,处于盛果生育期。试验地设置6片,每片林地选择15株山核桃树木,树高1 m以上,在树冠外围观察、测量、分析。
1.3 山核桃生物学特性观察在试验区域内,选取长势良好的山核桃成年树20株和幼树20株,观察其生物学特性和物候期(年周期),其中包括树皮颜色、芽、花、幼枝、新梢、叶、果实等器官。为了调查结果准确,入选树处于不同地段和地势,且具有一定间隔距离,确保相互间不受干扰,影响结果分析,同时,分别对入选树外围叶片、内膛叶片进行观察比较。
1.4 净光合速率日变化的测定使用美国Licor-6400光合作用测定系统,采用透明叶室测定。利用自然光照,直接输出数据包括叶温、进气CO2浓度、叶室CO2浓度、胞间CO2浓度、光合速率、蒸腾速率、气孔导度等。为了减少试验误差,自始至终由同一人读取测量数据。测定时间为08:00至15:30,自动记录。
2 结果与分析
2.1 山核桃生物学特性及物候期山核桃是落叶乔木,树皮光滑,幼树呈青褐色,老树灰白色,裸芽较瘦小,芽、幼枝、新梢、叶背面、外果皮均密被褐黄色腺鳞。奇数羽状复叶5~7小叶(7小叶较多),椭圆状披针形,长10~17 cm,宽2.5~5.0 cm,先端渐尖,基部楔形,复叶边缘锯齿形、细尖。单性花,雌雄同株异熟;雄花呈柔荑花序,每3条花序成一束,腋生于花序总梗之上,雄花包含有1个大苞片和2枚小苞片,雄蕊2~7枚,无花丝,花药有毛;雌花为顶生穗状,直立雌花1~3枚。果实核果状,卵球形,同一雌花上结1个果实时无果柄,结2个果实时有1个果实无果柄,另1个果有约2 cm长的果柄;外果皮有4个棱脊,干燥后革质四瓣裂开;山核桃核果隔膜内及内壁无空隙。幼苗时,主根发达,树体达到成年时,由于土壤中砾石阻挡作用导致主根没有侧根生长发达。根部富含单宁物质,受伤后容易变黑色。
年周期(物候期)调查结果见表1。从表1可以看出,山核桃约在每年4月初开始萌动出芽,4月中、上旬小叶展开,4月底—5月上旬中开花,5—7月是山核桃种实膨大期,8月—9月上旬是种实及油脂形成期,9月上、中旬种实成熟,11月上、中旬出现落叶,然后树体进入休眠状态,生长期在200~210 d。雄花期4~5 d,雌花期稍长,为5~10 d,当雌花柱头顶端呈现红褐色,且有少量黏液时,说明雌花已经成熟,可以进行人工授粉。
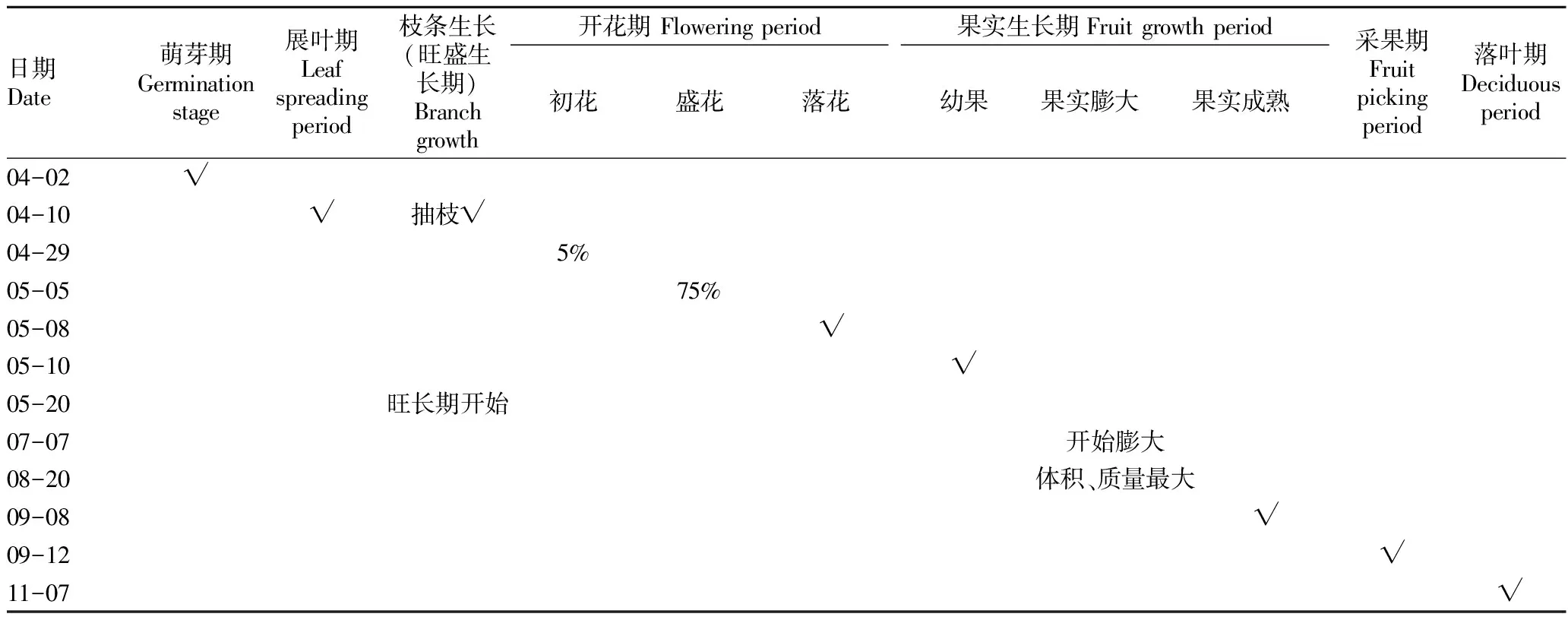
表1 山核桃物候期
果实动态生长调查发现:单果质量总体上不断增长,生长曲线呈“S”型(“快—慢—快”趋势),初果后20 d内果实(只有0.071~0.095 g/果)增长极慢,此后的40 d单果质量增长较为缓慢,到7月上旬果实开始快速生长,且维持30 d以上(果实纵横径发育趋势与单果质量生长类似)。在果实快速膨大时,可溶性糖从最高含量(65%)降低至10%以下,同时,脂肪含量快速增加,达70%,说明物质转化的主要方向是油脂形成。蛋白质表现为果实膨大前一直维持一定水平,到果实膨大开始时积累到最高(达15.8%),此后稍有下降。
2.2 光合生理指标变化
2.2.1净光合速率日变化。从图1可以看出,在一天中,环境中的光照强度、温度、土壤和大气的水分状况、空气中CO2浓度及植物体当时的水分含量、光合中间产物含量、气孔开度等生理状况都在不断变化,且这些变化的结果又会使光合速率发生进一步的日变化。从图1可以看出,在13:00以前的光合速率为负值,说明当天天气状况不是十分良好,上午的光照强度不能使山核桃进行充分的光合作用,导致光呼吸和暗呼吸的速率大于光合作用的速率,从而造成上午的净光合速率为负值。13:00以后光照逐渐加强,净光合速率随之迅速上升,在13:56达到最大值。山核桃有明显的光合“午休”现象,“午休”的主要因素是大气干旱和土壤干旱。午间气温升高,叶片蒸腾失水加剧,若此时土壤水分也亏缺,山核桃的失水大于吸水,就会引起萎蔫和气孔导度降低,使CO2吸收减少。另一方面,中午及午后的强光照、高温、低CO2浓度等环境条件都会促进光呼吸的激增,呈现光抑制现象,同样也会使光合速率下降。所以,山核桃光合速率的“午休”现象表明当地土壤中缺少水分。从理论角度,“午休”现象直接影响光合产物的生产,甚至达到30%或更多,因此,实际生产中,及时灌溉,缓解“午休”程度是增强树木光合能力的一个途径。山核桃光合速率在14:56—15:01呈现另外一次负值,这可能是光照不足造成的。一般而言,山核桃的光合速率在上午高于下午,这主要是因为山核桃体内的酶在经过一段时间(夜间)的自我调整后,恢复和保持一段时间内的高光合速率现象,也从另一个角度证明了山核桃体内的酶在长期进化过程后形成了“光周期”。但上午的光合速率并不高,可能与当日的天气有关。
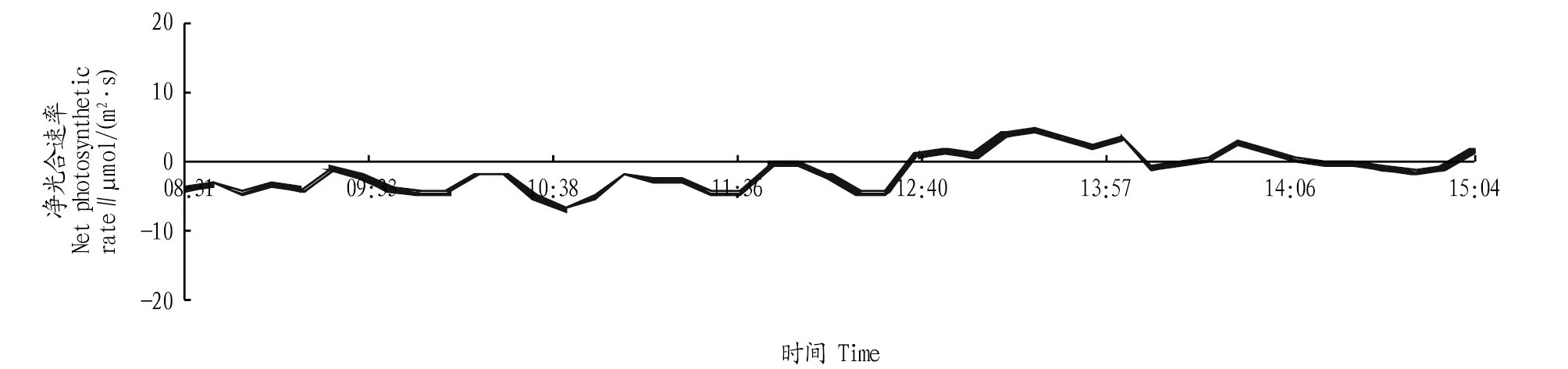
图1 净光合速率日变化Fig.1 Daily variation of net photosynthetic rate
2.2.2气孔导度日变化。山核桃叶片气孔导度的日变化见图2。由图1和图2对比发现,山核桃叶片气孔导度与叶片光合速率的日变化趋势呈正相关关系,趋势也一致。这与许大全等[19-20]提出的光合速率对气孔导度具有反馈调节的作用观点相一致即在利于植物叶肉细胞光合作用时气孔导度增大;不利于光合作用进行时,气孔导度减小。同时,从图2和图3比较可以看出,细胞内CO2浓度受气孔导度的影响较大。
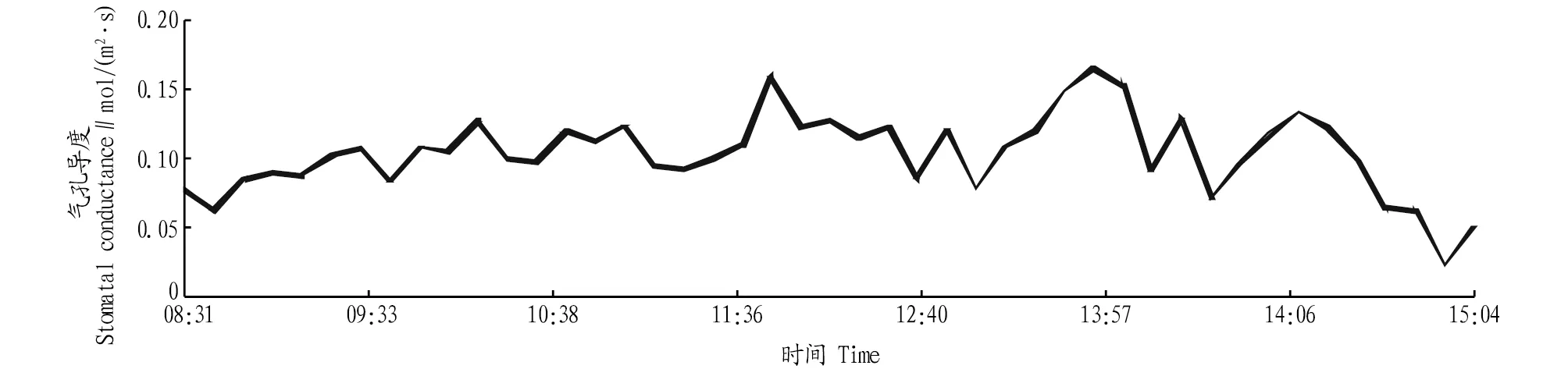
图2 气孔导度日变化Fig.2 Daily variation of stomatal conductance
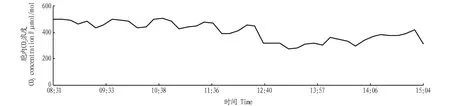
图3 胞内CO2浓度日变化Fig.3 Daily variation of intracellular carbon dioxide concentration
2.2.3胞内CO2浓度日变化。从图3可以看出,当山核桃光合速率出现较大值时,细胞内CO2浓度较小,表明光合作用是利用光能消耗CO2,从而合成有机产物的过程,所以山核桃细胞内CO2浓度在某种意义反映了山核桃光合速率的快慢。将图3、4与图1进行比较发现,胞内CO2浓度日变化和大气中CO2浓度日变化与光合速率日变化呈负相关,胞内CO2浓度和大气中CO2浓度在早晚均显示较高值。大气中CO2浓度在14:56时有一个小的峰值,细胞内CO2浓度也保持一致的变化趋势,这对应了山核桃的光合“午休”现象。
2.2.4空气中CO2浓度日变化。从图4可以看出,空气中CO2浓度在一天中的早晚较高,说明山核桃在该时段的光合速率较低。空气中的CO2浓度在13:00—14:00较低,与图1比较,该段时间正是山核桃光合速率在一天中较高的一段时间,应及时补充山核桃光合作用所必需的CO2,提高山核桃的产量。
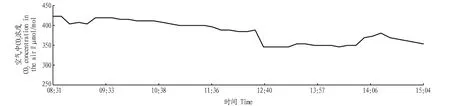
图4 空气中CO2浓度日变化Fig.4 Daily variation of CO2 concentration in the air
2.2.5蒸腾速率日变化。从图5可以看出,蒸腾速率和光合速率的日变化趋势类似,呈同步变化趋势。表明蒸腾速率和光合速率均受到叶片气孔开启的调节作用。图5的变化趋势与图2大致相当,表明当气孔阻力增大,气孔导度变小时蒸腾速率也会随之而降低。图4和图5进行比较发现,在高CO2浓度条件下,蒸腾速率迅速下降。蒸腾速率随CO2浓度增加而减小的趋势与气孔导度随CO2浓度变化而变化相同[21]。
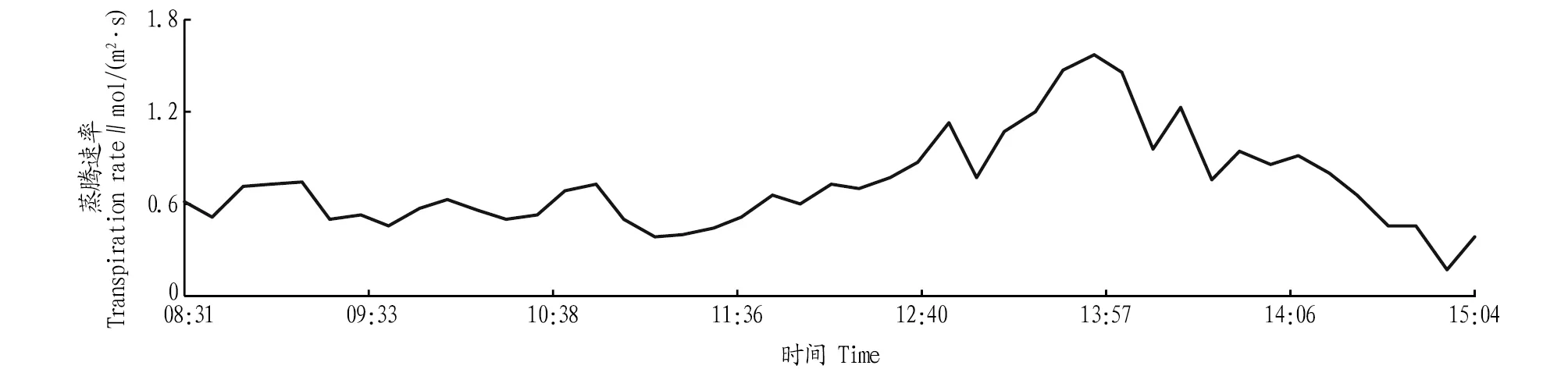
图5 蒸腾速率日变化Fig.5 Daily variation of transpiration rate
2.2.6叶温日变化。从图6可以看出,一天中随气温和光强的增加,叶温增加,变化趋势相同。在13:00左右叶温最高,此后的1 h叶温变化平稳,在24.5 ℃左右变动。由图5和图6可知,随着蒸腾速率的增大,叶温呈下降趋势,反之若蒸腾速率减小,叶温会随之升高。因此,当蒸腾作用受阻时,叶温升高,在叶温高于37 ℃时会影响叶肉细胞中与光合作用关联酶的活性,当叶温过高时,会灼伤叶片,影响叶肉的光合机能[22]。
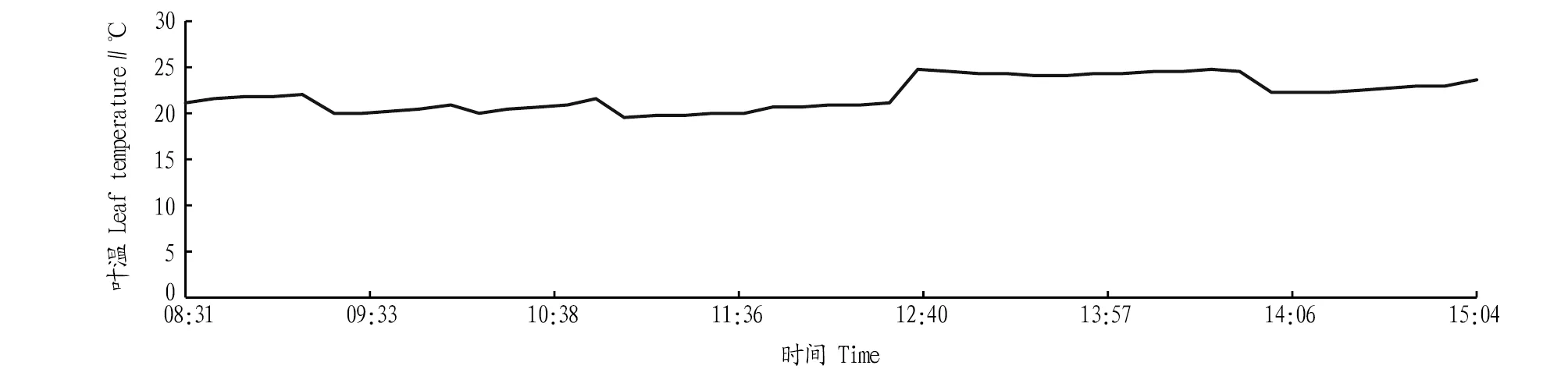
图6 叶温日变化Fig.6 Daily variation of leaf temperature
2.2.7光合作用后CO2浓度日变化。图1和图7呈明显的负相关关系,上午山核桃光合速率呈负值时CO2浓度较高,随着山核桃光合速率的增高,光合作用后的CO2浓度降低。图4和图7的变化趋势基本一致,这说明CO2被山核桃利用,进行光合作用,形成了有机物,此时应及时补充CO2以保证山核桃光合作用顺利进行,为山核桃的丰产奠定基础。
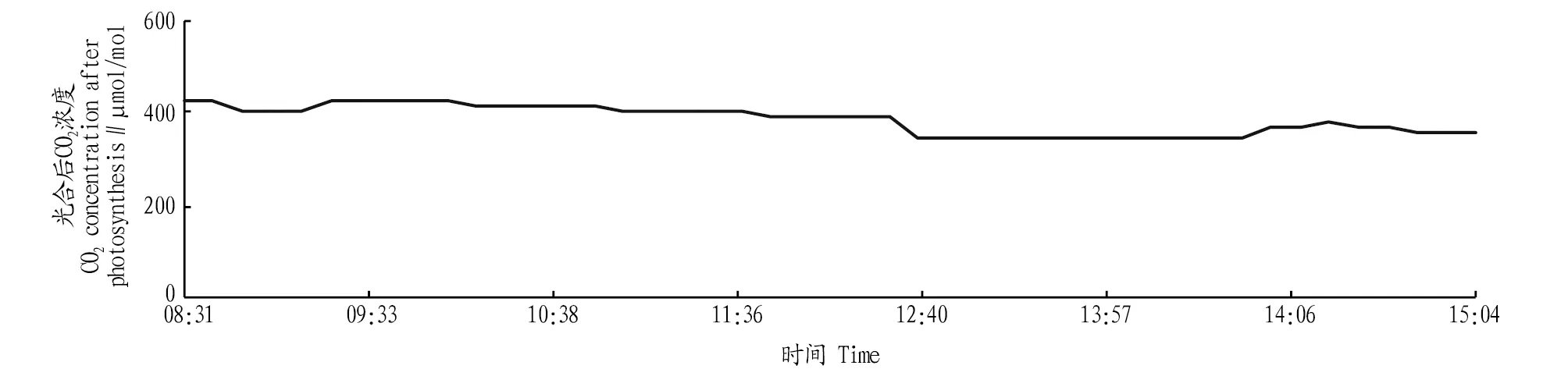
图7 光合后CO2浓度日变化Fig.7 Daily changes in CO2 concentration after photosynthesis
3 结论与讨论
(1)该研究结果进一步证实了山核桃的喜温特性与光合性能密切相关。提高光合性能,或从品种到栽培措施来改善光合物质的积累是山核桃丰产的基础工作;8—9月上旬是山核桃种实及油脂形成期,9月上、中旬种实成熟,11月上、中旬出现落叶,然后树体进入休眠状态,生长期在200~210 d。
(2)通过山核桃净光合速率日变化的研究,结果表明,在自然条件下,山核桃的日光合速率有明显光“午休”现象,光合作用呈双峰曲线。蒸腾速率和光合速率在一天内的变化趋势相似。叶片气孔导度的日变化趋势与光合速率的日变化趋势有很强的正相关(平行变化趋势),叶片气孔导度的日变化也与空气中的CO2浓度的日变化呈负相关,随着空气中CO2浓度的增高叶片的气孔导度降低,在气孔导度降低的同时又影响到蒸腾速率,使蒸腾速率随气孔导度的降低而降低,蒸腾速率的降低则造成叶温的升高,过高的叶温影响光合作用酶活性的发挥致使光合速率也随之下降。
(3)山核桃90%~95%的干物质是通过光合作用获取,因此,研究光合作用,且发挥最大效益时,对山核桃的丰产和质量形成具有重要意义。另外,山核桃的栽培技术不容忽视,要根据物候期生长特点(年周期),更好地利用光合产物,积累营养物质,同时,制定相应的措施;山核桃对立地条件要求苛刻,因此在选择宜林地上要重视小气候环境。山核桃是雌雄花异熟,花期不一致也是导致山核桃低产的一个重要因素。此外还应加强山核桃的栽培管理,对病虫害的防治采用“以防为主,综合防治”的方针,适当时可以增加山核桃林地的CO2浓度,以促进光合作用。
猜你喜欢
生态学报(2024年1期)2024-01-25
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
食品科学(2013年14期)2013-03-11
赤峰学院学报·自然科学版(2012年19期)2012-10-14
植物营养与肥料学报(2011年2期)2011-10-26
