
石榴MAPK家族基因鉴定及其响应冷胁迫的表达分析
2023-10-27 09:00:25陈利娜曹尚银唐丽颖李好先严琼李松开杨庆华鲁振华
果树学报 2023年10期
陈利娜 曹尚银 唐丽颖 李好先 严琼 李松开 杨庆华 鲁振华


DOI:10.13925/j.cnki.gsxb.20230136
摘 要:【目的】评价不同石榴种质资源籽粒硬度及抗寒性,筛选可能参与调控石榴抗寒性的MAPK家族基因。【方法】以31份石榴种质资源为试材,进行抗寒性及籽粒硬度评价;基于全基因组筛选石榴MAPK家族基因,对其进行进化、基因结构和蛋白理化性质分析,同时利用实时荧光定量PCR(real-time quantitative PCR,qRT-PCR)分析冷胁迫对石榴MAPK家族基因表达模式的影响。【结果】31个石榴品种籽粒硬度及半致死温度测定结果表明,峄城粉红牡丹、淮北六棱甜和鲁白榴2号等硬籽石榴抗寒性较强,华紫、以3和玛丽斯等软籽石榴抗寒性较弱。基于石榴全基因组鉴定出17个MAPK家族基因,广泛分布于8条染色体上;MAPK家族所有成员主要分为3个亚类,其中,A和B亚类成员主要包含PKc_MAPKK_plant_like和PTZ00024结构域,C亚类主要包含PLN00034结构域,所有成员均具有S_TKc结构域;各成员氨基酸残基数量分布在314~860 aa,外显子数目1~18个,蛋白分子质量为34 910.05~97 965.26 u,等电点4.94~9.35;PgMKK2、PgMPK6、PgMPK9、PgMPK16和PgMPK13在峄城粉红牡丹响应冷胁迫过程中表现为显著上调,PgMKK8、PgMPK1-1和PgMKK4在瑪丽斯响应冷胁迫过程中表现为显著上调;PgMKK2、PgMPK6、PgMPK9、PgMPK16和PgMPK13在峄城粉红牡丹响应冷胁迫过程中的表达量显著高于玛丽斯,PgMKK8和PgMPK1-1在玛丽斯响应冷胁迫过程中的表达量显著性高于峄城粉红牡丹;PgMKK3在峄城粉红牡丹不同时间均未检测到表达,在玛丽斯中表现为先升高后降低的趋势;PgMPK12-2在玛丽斯不同时间均未检测到表达,在峄城粉红牡丹中表现为逐渐升高的趋势。【结论】石榴MAPK家族基因响应冷胁迫信号,其中,PgMKK2、PgMPK6、PgMPK12-2和PgMPK9可能参与正调控石榴的抗寒性。
关键词:石榴;籽粒硬度;抗寒性;MAPK;表达分析
中图分类号:S665.4 文献标志码:A 文章编号:1009-9980(2023)10-2076-13
Identification of MAPK family genes and analysis of their expression patterns in response to cold stress in pomegranate
CHEN Lina1, CAO Shangyin1#, TANG Liying2, LI Haoxian1, YAN Qiong3, LI Songkai3, YANG Qinghua3, LU Zhenhua1*
(1Zhengzhou Fruit Research Institute, Chinese Academy of Agricultural Sciences, Zhengzhou 450009, Henan, China; 2Hebei Chengde Academy of Agriculture and Forestry Sciences, Chengde 067000, Hebei, China; 3Mengzi Fruit and Vegetable Technology Promotion Station, Mengzi 661100, Yunnan, China)
Abstract: 【Objective】 Cold stress is one of the most important factors limiting the progress of pomegranate production. The objectives of this study were to compare the cold hardiness among 31 pomegranate cultivars with wide distribution of seed hardness, and analyze the expression pattern of Mitogen-activated protein kinase (MAPK) family genes in response to cold stress. The results can pave the way for shedding light to the function of Mitogen-activated protein kinase cascades in pomegranate cod tolerance. 【Methods】Plant materials were cultivated in the National Horticulture Germplasm Resources Center of China (NHGRC), with conventional cultivation management. Juvenile branches of 31 pomegranate cultivars were sampled in mid-January, the relative electrolyte conductivity (REC) was measured, and the semi-lethal temperature (LT50) was calculated for the evolution of cold hardiness. Seed hardness was detected by the TA-XT texture apparatus. MAPK family genes of Arabidopsis were used as queries to search in the whole pomegranate genome database, and reference genome sequence of ‘Tunisia was obtained from NCBI database. Potential members of MAPK family were identified. Phylogeny relationship, gene structure and protein physicochemical properties were analyzed. Moreover, we performed real-time quantitative PCR (qRT-PCR) to analyze the expression pattern of 17 MAPK family genes in response to cold stress. 【Results】 The seed hardness and cold tolerance were evaluated. A total of 17 MAPK family genes were identified, which widely distributed on different chromosomes. All the members of the MAPK family could be mainly divided into three sub-classes, among which the members of subclass A and B mainly included PKc_MAPKK_plant_like and PTZ00024 Domain, subclass C mainly contained the PLN00034 domain, and all members contained a S_TKc domain. The results of analysis of physicochemical properties of proteins showed that the number of amino acid residues of each member distributed from 314 to 860 aa, the number of exons was from 1 to 18, the molecular weight of the protein was from 34 910.05 to 97 965.26 u, and the isoelectric point was from 4.94 to 9.35. Specific primer was designed for each PgMAPKs and PgMAPKKs, and their expression patterns were detected. The results showed that 12 out of 17 members were activated after low-temperature treatment. Yichengfenhongmudan that showed strong cold tolerance, and Malisi with weak cold tolerance, were both selected to compare the expression pattern of MAPKs. PgMKK2, PgMPK6, PgMPK9, PgMPK16 and PgMPK13 were all rapidly activated after low-temperature treatment in Yichengfenhongmudan. PgMKK8, PgMPK1-1 and PgMKK4 were rapidly activated in Malisi. PgMKK2, PgMPK6, PgMPK9, PgMPK16 and PgMPK13 showed significantly higher expression level in Yichengfenhongmudan than those in Malisi after low-temperature treatment. The expression of PgMPK3, PgMPK12-1, PgMPK20, PgMPK18 and PgMKK6 was not affected by low-temperature treatment. PgMKK8, PgMPK1-1 and PgMKK4 were up-regulated in Malisi than Yichengfenhongmudan. The expression of PgMPK12-2 was not activated after low-temperature treatment in Malisi, but showed gradual increase in Yichengfenhongmudan. The expression level of PgMKK3 increased at first and then decreased during low-temperature treatment in Malisi, whereas it was not detected in Yichengfenhongmudan. 【Conclusion】 Pomegranate MAPK family genes responded to cold stress signals. PgMKK2, PgMPK6, PgMPK12-2 and PgMPK9 might positively regulate cold tolerance.
Key words: Pomegranate (Punica granatum L.); Seed hardness; Cold resistance; MAPK; Expression analysis
石榴(Punica granatum L.)属千屈菜科(Lythraceae)石榴属(Punica L.)落叶果树,是中国重要的经济作物。石榴抗寒性差,硬籽石榴一般遇-17 ℃以下低温会出现冻害,以突尼斯软籽为代表的软籽石榴遇-10 ℃以下低温即出现冻害[1-2]。软籽石榴商品价值高,约占石榴总市场的80%,已成为多个石榴主产区的主栽品种。但是,目前主栽的软籽石榴突尼斯软籽和中农红等品种在河南、山东、陕西等主产区频发冻害,给果农造成了极大的损失,这已成为制约石榴产业健康和可持续发展的主要问题。
为缓解石榴冻害问题,国内外学者通过冬季扣棚或埋土、抗寒砧木嫁接和冬季喷施防冻剂等栽培方式防寒[3],并在部分地区取得了缓解冻害的效果。但与此同时,培育优良抗寒软籽石榴新品种仍是解决冻害问题的根本途径。目前,国内软籽石榴资源较少,收集或创制并筛选抗寒性强的软籽石榴资源可加快优良抗寒软籽石榴培育进程。本研究团队对经过多年收集或创制获得的软籽石榴种质资源进行抗寒性评价,其结果可为亲本选择提供依据。石榴抗寒性评价方法主要包括组织褐变法、生理生化指标测定法和电导法等,其中电导法是石榴抗寒性最可行的评价方法[4-6]。
截至目前,已有大量报道植物抗寒调控关键基因的研究,其中CBF(C-repeat binding factors)基因是调控植物抗寒性的重要开关基因[7],而促分裂原活化蛋白激酶(Mitogen-activated protein kinases, MAPK)可通过级联反应调控CBF基因的表达從而调控植物抗寒性。MAPK是一类保守的丝氨酸-苏氨酸蛋白激酶,主要通过逐级磷酸化放大和传递细胞外来刺激[8],在植物生长发育和胁迫响应过程中发挥重要作用[9-12]。MEKK1-MKK1/2-MPK4级联反应可通过与MPK3和MPK6拮抗,激活CBF基因表达从而提高植物抗寒性[13-15];MPK3和MPK6等主要通过调控质膜H+-ATP酶活性,磷酸化ICE1蛋白,从而抑制CBF基因表达,负调控植物抗寒性[16-18]。
关于石榴抗寒性机制研究的相关报道较少。刘贝贝[19]研究表明,CBF1基因是参与调控石榴抗寒性的关键基因。同时,MAPK级联途径相关基因在硬籽群体内受强烈选择,这从进化角度初步解释了大部分硬籽石榴抗逆能力强于软籽石榴的原因,暗示了MAPK级联途径对石榴抗逆过程中的作用[20]。Ren等[21]的研究发现,MAPK家族基因可能参与石榴顶端分生组织、花和果实发育过程。但MAPK家族基因对石榴抗寒性的影响尚不明确。
因此,为研究MAPK家族基因在石榴冷胁迫过程中表达变化,笔者在本研究中对31份种质资源进行籽粒硬度及抗寒性评价。同时,基于全基因组挖掘MAPK家族基因,对MAPK家族基因成员进行系统进化、基因结构和蛋白理化性质分析,明确MAPK家族基因在石榴响应冷胁迫过程中的表达模式,进而为石榴抗寒机制的研究提供理论支持。
1 材料和方法
1.1 植物材料及试验地概况
试验以31个石榴品种的果实和枝条为试材,进行籽粒硬度和抗寒性检测。试验材料均取自国家园艺种质资源库(郑州)内7年生石榴树,株行距2 m × 4 m,树体主干开心形,管理方式采用常规栽培管理,土壤为壤砂土。
1.2 试验方法
1.2.1 不同品种石榴籽粒硬度的测定 成熟期取石榴果实籽粒,每个果实取上中下部各10粒种子混合测定(每个果实测定30个籽粒),每个品种取3个果实分别测定。测定时使用纱布去除籽粒外种皮,取光洁未被破坏的籽粒进行硬度测定,使用TA-XT质构仪(英国SMS)测定,选择P2探头(位移1.3~2.0 mm),测前测后速率5 mm·s-1,测中速率1 mm·s-1。
1.2.2 不同品种石榴枝条电导率的测定 2021年1月中旬,选取每个品种1年生健壮枝条20~30根,用去离子水洗净,吸干水分后剪成8~10 cm小段,每组5段,分成3组,捆绑并用保鲜膜包裹。将分组的枝条于4 ℃冰箱预冷24 h,之后用低温培养箱进行梯度冷处理,处理温度分别为-4、-8、-12、-16、-20 ℃,降温速率4 ℃·h-1,达到目的温度后保持24 h,以4 ℃为对照。
在50 mL离心管中加入25 mL去离子水,将低温处理后的枝条剪成0.5 cm的小段,各称取1 g,加入去离子水中,置于25 ℃摇床中,90 min,用电导率仪(雷磁 DDS-307型号)测量初始电导值,然后沸水浴20 min,自然冷却至室温,测量其终电导值。相对电导率=初始电导值/终电导值[6]。Logistic回归方程y=k/(1+ ae-bx),其中y为相对电导率,x为处理温度,k为当x趋于无穷大时的值,a和b为方程参数。计算半致死温度(LT50)。
1.2.3 MAPK家族基因鉴定及生物信息分析 拟南芥MAPK家族的30个成员的氨基酸序列(包括10个MKK和20个MPK成员)来源于拟南芥基因数据库TAIR(https://www.arabidopsis.org/)[22]。以突尼斯软籽石榴基因组为参考基因组通过NCBI blast+-2.9.0进行本地blast获得。氨基酸序列通过在线软件emboss[23](https://www.ebi.ac.uk/Tools/st/emboss_transeq/)翻译获得。各编码蛋白的分子质量和理论等电点通过在线软件ProtParam(https://web.expasy.org/protparam/)进行预测[24]。MAPK家族基因系统发育树的构建和多序列比对使用本地软件MEGA 7.0[25],采用邻接法,bootstrap值为1000,进化树可视化通过Evolview在线软件[26]。氨基酸序列的保守结构域通过在线软件Batch CD-Search(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)进行预测[27],通过DNAMAN进行MAPK家族基因多序列比对。通过在线网站String(https://cn.string-db.org/)在线网站进行蛋白互作网络预测。
1.2.4 MAPK响应冷胁迫过程中表达模式分析 取峄城粉红牡丹、玛丽斯1年生枝条,8~10 cm小段,每组5段,分成3组。4 ℃开始降温,温度达到-12 ℃时,分别处理0(对照)、5、10、20、30、60、120 min后取出枝条剪成0.5 cm小块,液氮速冻,-80 ℃保存。每个处理设3次生物学重复。
以石榴PgActin为内参基因[28],实时荧光定量PCR(real-time quantitative PCR,RT-qPCR)测定各基因在不同胁迫条件下的相对表达量。利用NCBI在线软件Primer-BLAST设计引物(表1)。选用Universal SYBR Green Master荧光定量试剂盒(Roche)在LightCycler? 480 Ⅱ (Roche)进行检测。Real Time-qPCR反应程序:95 ℃ 5 min; 95 ℃ 10 s、56 ℃ 10 s、72 ℃ 10 s,共 45 个循环;95 ℃ 5 s、65 ℃ 1 min,10个循环;40 ℃冷却。
1.3 数据统计与分析
采用Excel 2013对枝条电导率进行计算和Logistic方程对31个石榴品种的电导率进行回归分析,荧光定量数据计算采用2-ΔΔCT法[29]。采用Excel 2013、SPSS 25.0对籽粒硬度与相对电导率相关性和荧光定量结果进行数据整理与分析。利用R语言gplots绘制不同基因表达热图。使用T检验和多重比较法进行差异显著性分析。
2 结果与分析
2.1 石榴籽粒硬度与抗寒性分析
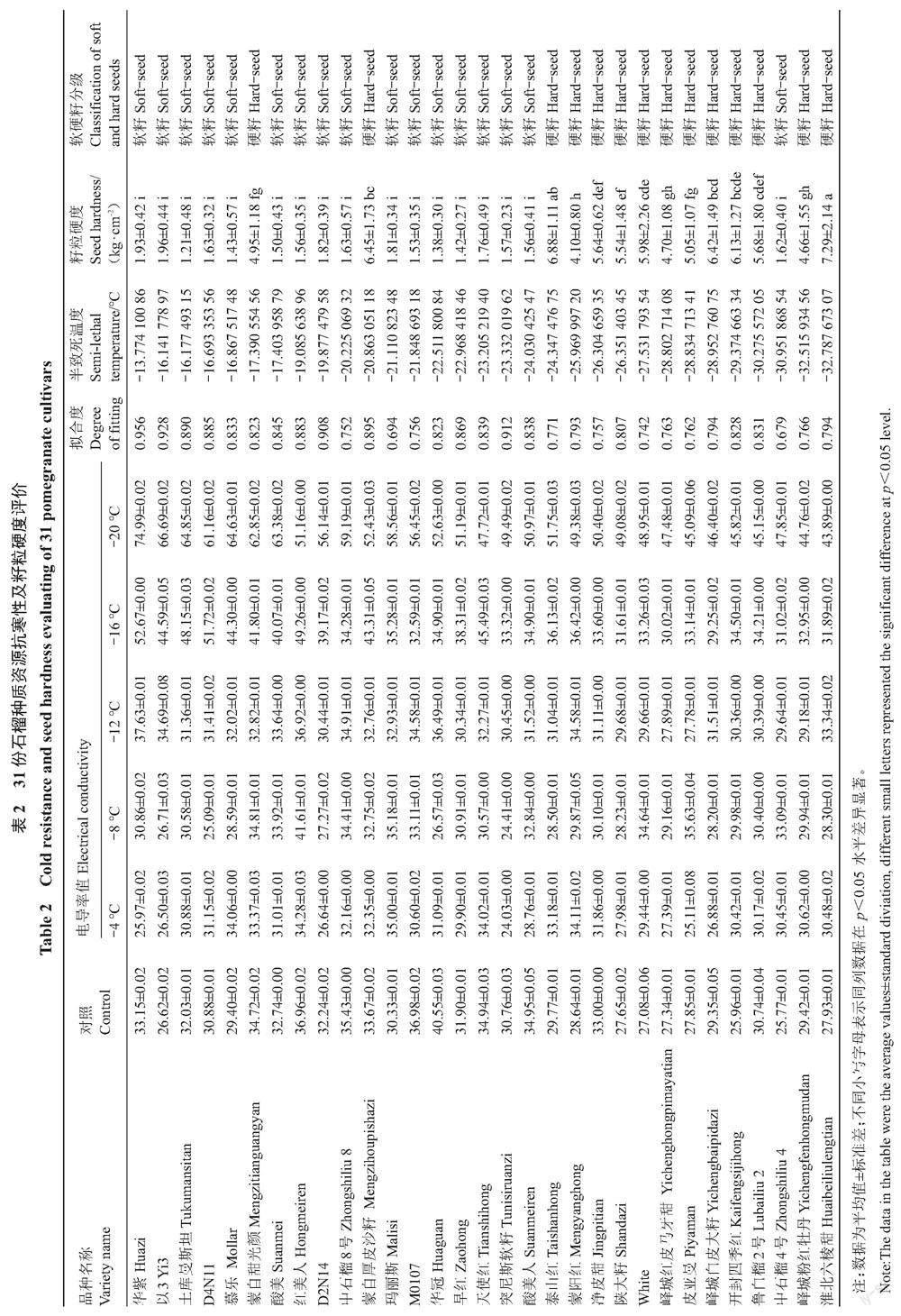
籽粒硬度测定结果表明,31个石榴品种籽粒硬度范围主要分布在(1.21±0.48)~(1.96±0.44) kg·cm-2和(4.10±0.80)~(7.29±2.14) kg·cm-2;依据Zarei等[30]分的籽粒硬度等级标准,将籽粒硬度分布在(1.21±0.48)~(1.96±0.44) kg·cm-2的品种归类为软籽,将籽粒硬度分布在(4.10±0.80)~(7.29±2.14) kg·cm-2的品种归类为硬籽;31份石榴种质资源共包括17份软籽和14份硬籽资源(表2)。
对31个石榴品种半致死温度测定结果表明,半致死温度分布范围在-13.78~-32.79 ℃;其中,峄城粉红牡丹、淮北六棱甜和鲁白榴2号等硬籽石榴抗寒性较强,华紫、以3和土库曼斯坦等软籽石榴抗寒性较弱;除蒙自甜光颜和蒙自厚皮沙籽外,硬籽石榴半致死温度低于大部分软籽石榴;同时,除中石榴4号外,软籽石榴半致死温度高于大部分硬籽石榴(表2)。
2.2 石榴MAPK家族基因的进化分析
基于拟南芥MAPK家族成员氨基酸序列从突尼斯软籽石榴基因组数据库中筛选出17个候选的MAPK同源基因(包括5个MAKK和12个MAPK成员)(表3),根据与拟南芥MAPK家族基因同源性进行命名,广泛分布于8条染色体上(图1)。通过拟南芥与石榴MAPK家族基因系统发育分析,石榴MAPK家族基因可分为3个亚类(A、B、C),其中,A亚类包含PgMPK12-1、PgMPK12-2、PgMPK6、PgMPK3、PgMPK13、PgMPK1-1和PgMPK1-2共7个成员,B亚类包含PgMPK18、PgMPK16、PgMPK20、PgMPK8和PgMPK9共5个成员,C亚类包含PgMKK2、PgMKK3、PgMKK4、PgMKK6和PgMKK8共5个成员(图2)。
2.3 石榴MAPK家族的结构分析
对石榴和拟南芥MAPK家族的功能域进行分析,发现这些基因均具有促分裂原活化蛋白激酶(Pkinase)的功能结构域,其中,A和B亚类成员主要包含PKc_MAPKK_plant_like、PTZ00024结构域,C亚类主要包含PLN00034结构域(图2)。基于石榴MAPK家族基因氨基酸多序列比对结果,发现其具有MAPK家族特有的保守结构域S_TKc(图3)。
石榴MAPK家族结构分析结果显示,石榴MAPK家族各成员氨基酸残基数量分布在314~860 aa(表3),外显子数目1~18个,同一亚类成员间外显子数目、位置及大小类型相近;A亚类成员外显子数目主要为6个,PgMAK1-1和PgMAK1-2外显子数目为1个,PgMAK12-1外显子数目为18个;B亚类成员外显子数目主要为10个;C亚类成员外显子数目主要为6~9个,PgMKK8外显子数目为1个(表3)。利用ProtParam对石榴MAPK家族基因蛋白理化性质进行分析,结果显示,石榴MAPK家族蛋白分子质量为34 910.05~97 965.26 u,等电点4.94~9.35(表3)。
2.4 石榴MAPK家族基因冷胁迫过程中表达模式分析
在抗寒性强的峄城粉红牡丹石榴响应冷胁迫的过程中,随着冷胁迫时间推移,MAPK家族基因表达模式主要分为3类(Ⅰ、Ⅱ和Ⅲ类)。Ⅰ类基因随着冷胁迫时间推移表达量表现为先升高后降低的趋势,主要包括PgMKK2、PgMPK13、PgMPK1-2、PgMPK8、PgMKK4、PgMPK20、PgMPK18共7个基因;Ⅱ类基因随着冷胁迫时间推移表达量表现为逐渐升高的趋势,主要包括PgMPK3、PgMKK6、PgMPK1-1、PgMPK16共4个基因;Ⅲ类基因表达量表现为逐渐降低的趋势,主要包括PgMPK12-1、PgMPK12-2、PgMPK6、PgMPK9和PgMKK8共5个基因;PgMKK3在峄城粉红牡丹石榴冷胁迫不同时间均未检测到表达(图4-A)。
在抗寒性差的玛丽斯石榴响应冷胁迫过程中,随着时间推移,MAPK家族基因表达模式同样主要分为3类(Ⅰ、Ⅱ和Ⅲ類)。Ⅰ类主要包括PgMKK3、PgMKK2、PgMPK20、PgMPK13和PgMPK1-1共5个基因;Ⅱ类主要包括PgMPK6、PgMKK6、PgMPK9、PgMPK16、PgMPK18和PgMPK3共6个基因;Ⅲ类主要包括PgMPK8、PgMKK4、PgMKK8、PgMPK12-1和PgMPK1-2共5个基因;PgMPK12-1在玛丽斯响应冷胁迫过程中未检测到表达(图4-B)。
其中,PgMKK2、PgMPK20和PgMPK13在峄城粉红牡丹石榴和玛丽斯石榴冷胁迫过程中表达量均表现为先升高后降低的趋势;PgMPK3、PgMKK6和PgMPK16表达量均表现为逐渐降低的趋势;PgMPK12-1和PgMKK8表达量均表现为逐渐升高的趋势(图4)。PgMPK9和PgMPK6在峄城粉红牡丹石榴响应冷胁迫过程中表现为逐渐升高的趋势,在玛丽斯石榴中表现为逐渐降低的趋势;PgMPK18在峄城粉红牡丹石榴不同时间表现为先升高后降低的趋势,在玛丽斯石榴中表现为逐渐降低的趋势;PgMPK1-2、PgMPK8和PgMKK4在峄城粉红牡丹石榴不同时间表现为先升高后降低的趋势,在玛丽斯石榴中表现为逐渐升高的趋势;PgMPK1-1在峄城粉红牡丹石榴不同时间表现为逐渐降低,在玛丽斯石榴中表现为先升高后降低;PgMKK3在峄城粉红牡丹石榴不同时间均未检测到表达,在玛丽斯石榴中表现为先升高后降低的趋势;PgMPK12-2在玛丽斯石榴不同时间均未检测到表达,在峄城粉红牡丹石榴中表现为逐渐升高的趋势(图4)。
MAPK家族基因在峄城粉红牡丹石榴和玛丽斯石榴间的相对表达量差异结果表明,PgMKK2、PgMPK6、PgMPK9、PgMPK16在响应冷胁迫过程中,在峄城粉红牡丹石榴中的相对表达量显著高于玛丽斯石榴;PgMPK13相对表达量在响应胁迫后5和10 min时在峄城粉红牡丹石榴和玛丽斯石榴间不存在显著性差异表达,20、30和60 min时在峄城粉红牡丹石榴中的相对表达量显著高于玛丽斯石榴;PgMKK8和PgMPK1-1在玛丽斯石榴中的相对表达量显著性高于峄城粉红牡丹石榴;PgMKK6、PgMPK12-1和PgMPK3的相对表达量在玛丽斯和峄城粉红牡丹石榴间无显著差异;PgMKK4、PgMPK8、PgMPK1-2响应冷胁迫前期在峄城粉红牡丹石榴中的相对表达量显著高于玛丽斯石榴、后期在玛丽斯石榴中的相对表达量显著高于峄城粉红牡丹石榴;PgMPK20和PgMPK18的相对表达量与石榴对冷胁迫的响应无显著相关性(图5)。
3 讨 论
3.1 枝条取样时间不同影响检测的半致死温度
笔者在本研究中测定的不同石榴品种半致死温度范围在-13.77~-32.79 ℃,与罗华等[31]发表的石榴半致死温度存在范围差异(-9.04~-11.77 ℃)。Ghasemi等[4]对不同取样时间的石榴枝条半致死温度检测的差异进行分析,发现同一品种1月份取石榴枝条进行测定获得的半致死温度显著低于3月份取枝条所获得的半致死温度。笔者取样时间为石榴枝条休眠期(1月中旬),罗华等[31]取样时间为萌芽前(3月中旬),这一结论解释了本研究测定的石榴半致死温度范围显著低于其发表范围的原因。但是,半致死温度的范围不影响品种间抗寒性强弱的差异比较。
3.2 石榴抗寒性可能与籽粒硬度呈正相关
大部分硬籽石榴果实内种皮次生细胞壁厚度显著高于软籽石榴,枝条较软籽石榴硬。石榴抗寒性和籽粒硬度性状可能存在连锁遗传现象。例如,NAC家族基因通过调控木质素、纤维素和半纤维素合成从而参与调控石榴籽粒硬度形成[28],在硬籽石榴中的表达量显著高于软籽石榴。同时,NAC家族基因参与植物对抗寒性等非生物胁迫的响应[32]。同时,已有研究报道生长环境温度对植物抗寒性具有显著影响,来源于云南产区的蒙自甜光颜石榴和蒙自滑皮沙籽石榴抗寒性、显著低于新疆、山东、河南和安徽产区的石榴,长期自然环境驯化对石榴抗寒性具有显著影响[33]。
3.3 石榴MAPK家族基因响应冷胁迫
目前,水稻[34]、玉米[35]、葡萄[36]、枣[37]、麻风树[38]等多个物种上MAPK家族基因被挖掘,在植物生长发育、抗逆等过程中发挥重要功能。笔者在本研究中分析石榴MAPK家族基因功能性结构域,发现在不同物种间MAPK家族基因功能性结构域具有保守性。在拟南芥和玉米中MAPK家族基因MPK3、MPK4、MPK6和MKK2等在冷处理30 min内表达量显著上调[34-35]。MKK2基因在马铃薯响应冷胁迫过程中显著上调表达[39]。MPK5[40]和MPK3[41]响应香蕉冷胁迫处理过程,并通过调控NAC042和ICE1基因表达从而参与调控香蕉抗寒性。石榴17个MAPK家族基因中12个均能不同程度地响应石榴冷胁迫信号。PgMKK2、PgMPK6、PgMPK9、PgMPK16、PgMPK13在峄城粉红牡丹石榴响应冷胁迫过程中表现为显著上调,PgMKK8、PgMPK1-1和PgMKK4在玛丽斯石榴响应冷胁迫过程中表现为显著上调,MAPK家族基因参与石榴响应冷胁迫过程。
3.4 PgMKK2-MPK6级联反应途径可能参与调控石榴抗寒性
已有研究表明,低温诱导MKK2蛋白磷酸化[15],MKK2激发MPK4/MPK6蛋白磷酸化[14],MPK4/MPK6通过调控ICE1和CBF基因表达从而调控植物抗寒性[14];同时MKK2-MAPK4/7通过调控ICE1基因表达从而参与调控植物抗寒性;MEKK1-MKK2-MPK4通过调控CBFs基因表达从而正调控植物抗寒性。而低温处理可显著激活石榴PgMKK2和PgMPK6基因表达,冷胁迫处理后PgMKK2和PgMPK6在峄城粉红牡丹石榴中的表达量显著高于玛丽斯石榴,同时,PgMPK6表达趋势与PgMKK2一致,推测PgMPK6基因表达上调可能与PgMKK2级联反应相关,PgMKK2-MPK6级联反应途径可能参与正调控石榴抗寒性。
3.5 PgMPK12-2可能参与正调控石榴抗寒性
油菜、擬南芥等物种中MPK12与MPK9功能冗余,正向调控脱落酸、茉莉酸、水杨酸[42-43]、低温和盐[44]等逆境胁迫过程。而石榴PgMPK12-2在玛丽斯石榴不同时间均未检测到表达,在峄城粉红牡丹石榴中表现为逐渐升高的趋势。PgMPK9在响应冷胁迫过程中在峄城粉红牡丹石榴中的表达量显著高于玛丽斯石榴。PgMPK12-2和PgMPK9可能参与石榴响应低温胁迫的过程。
4 结 论
笔者在本研究中基于石榴全基因组共挖掘到17个MAPK家族基因成员,分布于不同染色体,根据系统发育树将其分为3个亚类,所有成员均含有S-TKc结构域,同一亚类成员间保守结构域、外显子数目具有保守性。在响应冷胁迫过程中,PgMKK2、PgMPK6、PgMPK9、PgMPK16和PgMAPK13在峄城粉红牡丹石榴中的表达量显著高于玛丽斯石榴,PgMKK8和PgMPK1-1在玛丽斯石榴中的表达量显著高于峄城粉红牡丹石榴,PgMKK2、PgMPK6、PgMPK12-2和PgMPK9可能正调控石榴抗寒性。
参考文献 References:
[1] 杨雪梅,苑兆和,尹燕雷,冯立娟,招雪晴. 不同石榴品种抗寒性综合评价[J]. 山东农业科学,2014,46(2):46-51.
YANG Xuemei,YUAN Zhaohe,YIN Yanlei,FENG Lijuan,ZHAO Xueqing. Comprehensive evaluation on cold resistance of different pomegranate cultivars[J]. Shandong Agricultural Sciences,2014,46(2):46-51.
[2] 姚方,王宁,曹尚银,马贯羊,司守霞,姚海雷.不同软籽品种石榴抗寒性综合评价[J]. 森林与环境学报,2016,36(3):373-379.
YAO Fang,WANG Ning,CAO Shangyin,MA Guanyang,SI Shouxia,YAO Hailei.Comprehensive evaluation on cold resistance of different pomegranate cultivars[J]. Journal of Forest and Environment,2016,36(3):373-379.
[3] 焦其庆,冯立娟,尹燕雷,崔洪涛. 石榴冻害及抗寒评价研究进展[J]. 植物生理学报,2019,55(4):425-432.
JIAO Qiqing,FENG Lijuan,YIN Yanlei,CUI Hongtao. Research progress on evaluation of freezing injury and cold resistance of pomegranate[J]. Plant Physiology Journal,2019,55(4):425-432.
[4] SOLOKLUI A A G,ERSHADI A,FALLAHI E. Evaluation of cold hardiness in seven Iranian commercial pomegranate (Punica granatum L.) cultivars[J]. HortScience,2012,47(12):1821-1825.
[5] 毕润霞,郝兆祥,侯乐峰,王庆军,李昭慧,王艳琴. 电导法评价石榴抗寒性方法的探讨[J]. 山东农业科学,2015,47(2):38-41.
BI Runxia,HAO Zhaoxiang,HOU Lefeng,WANG Qingjun,LI Zhaohui,WANG Yanqin. Evaluation on cold resistance of pomegranate by conductivity method[J]. Shandong Agricultural Sciences,2015,47(2):38-41.
[6] 劉贝贝,陈利娜,牛娟,李好先,张杰,曹尚银. 6个石榴品种抗寒性评价及方法筛选[J]. 果树学报,2018,35(1):66-73.
LIU Beibei,CHEN Lina,NIU Juan,LI Haoxian,ZHANG Jie,CAO Shangyin. Selection of methods for evaluation on cold tolerance of six pomegranate varieties[J]. Journal of Fruit Science,2018,35(1):66-73.
[7] ZHANG F,JIANG Y,BAI L P,ZHANG L,CHEN L J,LI H G,YIN Y,YAN W W,YI Y,GUO Z F. The ICE-CBF-COR pathway in cold acclimation and AFPs in plants[J]. Middle-East Journal of Scientific Research,2011,8(2):493-498.
[8] TIAN K,LI Q,ZHANG X M,GUO H Y,WANG Y H,CAO P L,XU S Y,LI W Y. Analysis of the expression and function of the CBL-CIPK network and MAPK cascade genes in Kandelia obovata seedlings under cold stress[J]. Frontiers in Marine Science,2023,10:1113278.
[9] PITZSCHKE A,SCHIKORA A,HIRT H. MAPK cascade signalling networks in plant defence[J]. Current Opinion in Plant Biology,2009,12(4):421-426.
[10] BERGMANN D C,LUKOWITZ W,SOMERVILLE C R. Stomatal development and pattern controlled by a MAPKK kinase[J]. Science,2004,304(5676):1494-1497.
[11] MOUSTAFA K,ABUQAMAR S,JARRAR M,AL-RAJAB A J,TR?MOUILLAUX-GUILLER J. MAPK cascades and major abiotic stresses[J]. Plant Cell Reports,2014,33(8):1217-1225.
[12] ZHANG M M,SU J B,ZHANG Y,XU J,ZHANG S Q. Conveying endogenous and exogenous signals:MAPK cascades in plant growth and defense[J]. Current Opinion in Plant Biology,2018,45:1-10.
[13] BANERJEE G,SINGH D,SINHA A K. Plant cell cycle regulators:Mitogen-activated protein kinase,a new regulating switch?[J]. Plant Science,2020,301:110660.
[14] TEIGE M,SCHEIKL E,EULGEM T,D?CZI R,ICHIMURA K,SHINOZAKI K,DANGL J L,HIRT H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis[J]. Molecular Cell,2004,15(1):141-152.
[15] FURUYA T,MATSUOKA D,NANMORI T. Phosphorylation of Arabidopsis thaliana MEKK1 via Ca2+ signaling as a part of the cold stress response[J]. Journal of Plant Research,2013,126(6):833-840.
[16] LIU Y K,ZHOU J. MAPping kinase regulation of ICE1 in freezing tolerance[J]. Trends in Plant Science,2018,23(2):91-93.
[17] ZHAO C Z,WANG P C,SI T,HSU C C,WANG L,ZAYED O,YU Z P,ZHU Y F,DONG J,TAO W A,ZHU J K. MAP kinase cascades regulate the cold response by modulating ICE1 protein stability[J]. Developmental Cell,2017,43(5):618-629.
[18] PONCE-PINEDA I G,CARMONA-SALAZAR L,SAUCEDO-GARC?A M,CANO-RAM?REZ D,MORALES-CEDILLO F,PE?A-MORAL A,GUEVARA-GARC?A ? A,S?NCHEZ-NIETO S,GAVILANES-RU?Z M. MPK6 kinase regulates plasma membrane H+-ATPase activity in cold acclimation[J]. International Journal of Molecular Sciences,2021,22(12):6338.
[19] 劉贝贝. 石榴抗寒品种筛选及转录因子PgCBF1功能分析[D]. 北京:中国农业科学院,2018.
LIU Beibei. Screening cold-tolerante cultivars of pomegranate and analyzing the function of PgCBF1 transcription factor[D]. Beijing:Chinese Academy of Agricultural Sciences,2018.
[20] LUO X,LI H X,WU Z K,YAO W,ZHAO P,CAO D,YU H Y,LI K D,POUDEL K,ZHAO D G,ZHANG F H,XIA X C,CHEN L N,WANG Q,JING D,CAO S Y. The pomegranate (Punica granatum L.) draft genome dissects genetic divergence between soft-and hard-seeded cultivars[J]. Plant Biotechnology Journal,2020,18(4):955-968.
[21] REN Y A,GE D P,DONG J M,GUO L H,YUAN Z H. Genome-wide identification and expression analysis of MAPK and MAPKK gene family in pomegranate (Punica granatum L.)[J]. Agronomy,2020,10(7):1015.
[22] LAMESCH P,BERARDINI T Z,LI D H,SWARBRECK D,WILKS C,SASIDHARAN R,MULLER R,DREHER K,ALEXANDER D L,GARCIA-HERNANDEZ M,KARTHIKEYAN A S,LEE C H,NELSON W D,PLOETZ L,SINGH S,WENSEL A,HUALA E. The Arabidopsis Information Resource (TAIR):Improved gene annotation and new tools[J]. Nucleic Acids Research,2012,40(Database issue):1202-1210.
[23] MADEIRA F,PEARCE M,TIVEY A R N,BASUTKAR P,LEE J,EDBALI O,MADHUSOODANAN N,KOLESNIKOV A,LOPEZ R. Search and sequence analysis tools services from EMBL-EBI in 2022[J]. Nucleic Acids Research,2022,50(W1):276-279.
[24] ARTIMO P,JONNALAGEDDA M,ARNOLD K,…,STOCKINGER H. ExPASy:SIB bioinformatics resource portal[J]. Nucleic Acids Research,2012,40(Web Server issue):597-603.
[25] KUMAR S,STECHER G,TAMURA K. MEGA7:Molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution,2016,33(7):1870-1874.
[26] HE Z L,ZHANG H K,GAO S H,LERCHER M J,CHEN W H,HU S N. Evolview v2:An online visualization and management tool for customized and annotated phylogenetic trees[J]. Nucleic Acids Research,2016,44(W1):236-241.
[27] LU S N,WANG J Y,CHITSAZ F,DERBYSHIRE M K,GEER R C,GONZALES N R,GWADZ M,HURWITZ D I,MARCHLER G H,SONG J S,THANKI N,YAMASHITA R A,YANG M Z,ZHANG D C,ZHENG C J,LANCZYCKI C J,MARCHLER-BAUER A. CDD/SPARCLE:The conserved domain database in 2020[J]. Nucleic Acids Research,2020,48(D1):265-268.
[28] QIN G H,LIU C Y,LI J Y,QI Y J,GAO Z H,ZHANG X L,YI X K,PAN H F,MING R,XU Y L. Diversity of metabolite accumulation patterns in inner and outer seed Coats of pomegranate:Exploring their relationship with genetic mechanisms of seed coat development[J]. Horticulture Research,2020,7:10.
[29] LIVAK K J,SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method[J]. Methods,2001,25(4):402-408.
[30] ZAREI A, ZAMANI Z, FATAHI R, MOUSAVI A, SALAMI S A, AVILA C , C?NOVAS F M. Differential expression of cell wall related genes in the seeds of soft- and hard-seeded pomegranate genotypes[J]. Scientia Horticulturae, 2016, 205: 7-16.
[31] 羅华,王庆军,郝兆祥,赵丽娜,陈颖,赵登超,毕润霞,孟健,马敏. 石榴抗寒种质筛选研究[J]. 中国果树,2018(4):46-52.
LUO Hua,WANG Qingjun,HAO Zhaoxiang,ZHAO Lina,CHEN Ying,ZHAO Dengchao,BI Runxia,MENG Jian,MA Min. Study on cold resistance of pomegranate germplasms[J]. China Fruits,2018(4):46-52.
[32] SINGH S,KOYAMA H,BHATI K K,ALOK A. The biotechnological importance of the plant-specific NAC transcription factor family in crop improvement[J]. Journal of Plant Research,2021,134(3):475-495.
[33] DUNN S. Factors affecting cold resistance in plants[J]. Plant Physiology,1937,12(2):519-526.
[34] RAO K P,RICHA T,KUMAR K,RAGHURAM B,SINHA A K. In silico analysis reveals 75 members of mitogen-activated protein kinase kinase kinase gene family in rice[J]. DNA Research,2010,17(3):139-153.
[35] KONG X P,L? W,ZHANG D,JIANG S S,ZHANG S Z,LI D Q. Genome-wide identification and analysis of expression profiles of maize mitogen-activated protein kinase kinase kinase[J]. PLoS One,2013,8(2):e57714.
[36] ?AKIR B,KILI?KAYA O. Mitogen-activated protein kinase cascades in Vitis vinifera[J]. Frontiers in Plant Science,2015,6:556.
[37] LIU Z G,ZHANG L M,XUE C L,FANG H,ZHAO J,LIU M J. Genome-wide identification and analysis of MAPK and MAPKK gene family in Chinese jujube (Ziziphus jujuba Mill.)[J]. BMC Genomics,2017,18(1):855.
[38] WANG H B,GONG M,GUO J Y,XIN H,GAO Y,LIU C,DAI D Q,TANG L Z. Genome-wide identification of Jatropha curcas MAPK,MAPKK,and MAPKKK gene families and their expression profile under cold stress[J]. Scientific Reports,2018,8:16163.
[39] CHEN Y,CHEN L,SUN X M,KOU S,LIU T T,DONG J K,TU W,ZHANG Y L,SONG B T. The mitogen-activated protein kinase kinase MKK2 positively regulates constitutive cold resistance in the potato[J]. Environmental and Experimental Botany,2022,194:104702.
[40] TAK H,NEGI S,RAJPUROHIT Y S,RAJPUROHIT Y S,MISRA H S,GANAPATHI T R. MusaMPK5,a mitogen activated protein kinase is involved in regulation of cold tolerance in banana[J]. Plant Physiology and Biochemistry,2020,146:112-123.
[41] GAO J,DOU T X,HE W D,SHENG O,BI F C,DENG G M,GAO H J,DONG T,LI C Y,ZHANG S,YI G J,HU C H,YANG Q S. MaMAPK3-MaICE1-MaPOD P7 pathway,a positive regulator of cold tolerance in banana[J]. BMC Plant Biology,2021,21(1):97.
[42] JAMMES F,SONG C,SHIN D,MUNEMASA S,TAKEDA K,GU D,CHO D S,LEE S M,GIORDO R,SRITUBTIM S,LEONHARDT N,ELLIS B E,MURATA Y,KWAK J M. MAP kinases MPK9 and MPK12 are preferentially expressed in guard cells and positively regulate ROS-mediated ABA signaling[J]. Proceedings of the National Academy of Sciences of the United States of America,2009,106(48):20520-20525.
[43] KHOKON M A R,SALAM M A,JAMMES F,YE W X,HOSSAIN M A,OKUMA E,NAKAMURA Y,MORI I C,KWAK J M,MURATA Y. MPK9 and MPK12 function in SA-induced stomatal closure in Arabidopsis thaliana[J]. Bioscience,Biotechnology,and Biochemistry,2017,81(7):1394-1400.
[44] 李萍,聶亭亭,张腾国,郑晟,王娟,毛玉珊. 白菜型油菜MPK12基因克隆及表达分析[J]. 西北农林科技大学学报(自然科学版),2017,45(5):26-33.
LI Ping,NIE Tingting,ZHANG Tengguo,ZHENG Sheng,WANG Juan,MAO Yushan. Clone and characterization of MPK12 gene from Brassica campestris[J]. Journal of Northwest A & F University (Natural Science Edition),2017,45(5):26-33.
收稿日期:2023-04-12 接受日期:2023-06-27
基金项目:国家重点研发计划项目(2021YFD1600802);中国农业科学院科技创新工程(CAAS-ASTIP-2023-ZFRI);河南省科技攻关项目(212102110421)
作者简介:陈利娜,女,助理研究员,研究方向为果树遗传育种。Tel:13283811852, E-mail:1571863765@qq.com。#为共同第一作者。
通信作者 Author for correspondence. Tel:0371-65330990,E-mail:luzhenhua@caas.cn
猜你喜欢
音乐天地(音乐创作版)(2021年7期)2021-10-13 08:11:12
草原歌声(2021年1期)2021-07-16 06:58:06
少先队活动(2021年1期)2021-03-29 05:27:06
小天使·一年级语数英综合(2018年4期)2018-06-22 10:33:26
中国中药杂志(2016年20期)2016-11-19 12:27:25
新疆农垦科技(2016年2期)2016-08-21 13:50:26
中国果菜(2016年9期)2016-03-01 01:28:40
中国马铃薯(2015年5期)2016-01-09 06:11:11
江苏农业科学(2015年5期)2015-10-20 21:08:26
江苏农业科学(2015年9期)2015-10-20 21:04:29
