
葡萄COMT基因家族的鉴定与表达分析
2023-10-27 09:00:25许雯雯高换超韩菲菲李凯薇贾山毅李桂荣
果树学报 2023年10期
关键词:功能分析
许雯雯 高换超 韩菲菲 李凯薇 贾山毅 李桂荣

DOI:10.13925/j.cnki.gsxb.20230113
摘 要:【目的】咖啡酸O-甲基轉移酶(COMTs)是木质素合成过程中一种多功能酶,参与多种初生代谢和次生代谢途径,在植物木质素次生物质合成及植物抗逆胁迫反应中起着重要作用。旨在鉴定葡萄COMT基因家族成员,探究其对葡萄真菌病害胁迫的响应,为葡萄抗病育种提供基因资源。【方法】基于拟南芥COMT基因搜索葡萄COMT基因家族,运用生物信息学方法研究葡萄COMT蛋白质理化性质、基因染色体定位、motif分析和启动子顺式作用元件等;利用荧光定量PCR法分析COMT基因在抗病品种摩尔多瓦和感病品种夏黑葡萄上接种霜霉病的表达模式。【结果】从葡萄中鉴定出26个COMT基因,主要位于第10、12和15号染色体上;蛋白质分子质量差异较大,属于不稳定的两性蛋白;亚细胞定位显示其蛋白主要位于细胞质和细胞外。从构建的系统进化树中发现,该家族分为Ⅰ和Ⅱ 2个亚组,在进化过程中比较保守,与拟南芥和水稻亲缘关系较近。启动子分析表明,葡萄COMT基因启动子包含丰富的植物激素响应和胁迫响应的顺式元件,接种葡萄霜霉病后,在抗病品种摩尔多瓦中,除COMT2基因外其余25条COMT基因在接种24 h后均显著上调,而在感病品种夏黑中只有63%的COMT基因出现显著上调,且抗病品种的表达量显著高于感病品种,其中以VvCOMT1、5、6、7、8、9和19上调最为显著。【结论】共鉴定了26个葡萄COMT基因家族成员,同时发现COMT基因在葡萄抗病品种中受霜霉病的强烈诱导,表明木质素在葡萄抗病中起重要作用,可为后期研究葡萄抗霜霉病分子机制提供候选基因。
关键词:葡萄霜霉病;咖啡酸-O-甲基转移酶;生物学分析;功能分析
中图分类号:S663.1 S436.631 文献标志码:A 文章编号:1009-9980(2023)10-2061-15
Identification and expression analysis of grape COMT gene family
XU Wenwen1, GAO Huanchao1, HAN Feifei1, LI Kaiwei1, JIA Shanyi1, LI Guirong1, 2*
(1School of Horticulture and Landscape Architecture, Henan Institute of Science and Technology, Xinxiang 453003, Henan, China; 2Henan Province Engineering Research Center of Horticultural Plant Resource Utilization and Germplasm Enhancement, Xinxiang 453003, Henan, China)
Abstract: 【Objective】Caffeic acid O-methyltransferase (COMTs) is a multifunctional enzyme in lignin synthesis. It participates in some primary and secondary metabolic pathways, and plays an important role in the synthesis of lignin secondary substances and the responses to stresses in plants. The European grape genome was used in the study to search for genes and proteins homologous to members of the COMT gene family, and identification of the final grape COMT gene family and analysis of their expression levels under grape downy mildew stress were carried out to provide a molecular basis and genetic resources for grape resistance breeding. 【Methods】 Firstly, the identified O-methyltransferase gene sequence was searched from the Arabidopsis database. Based on the Arabidopsis COMT gene, the grape COMT gene family was searched by BLASTP tool, and candidate genes of the VvCOMT family were obtained by search and comparison, and duplicate and redundant sequences were deleted. The gene structure, protein physicochemical properties, protein secondary structure, chromosome location and promoter cis-acting elements were obtained using various biological online analysis software; Moldova and Summer Black were selected as materials to inoculate the pathogen spores of grape downy mildew on the leaves respectively. The samples were taken at 0, 6, 24, 48 h and 120 h after inoculation, and the expression patterns of the COMT gene family in Moldova and Summer Black after inoculation of downy mildew pathogen were analyzed by real-time fluorescence quantitative method. 【Results】A grape COMT gene family consisting of 26 COMT members was obtained through the identification, and all COMT genes had a Methyltransfer_2. The C-terminal catalytic domain (PF00891) was named VvCOMT1-26 according to the chromosome position, 84.6% of the grape COMTs had an amino acid length of over 300 aa, and their amino acid numbers ranged from 189 to 395. Among them, VvCOMT24 (VIT_215s0048g02460) had the longest sequence with 395 amino acids, while VvCOMT3 (VIT_208s0032g01130) has the shortest sequence with only 189 amino acids. The relative molecular weight of the proteins were between 21 179.45 (VvCOMT3) and 43 521.88 (VvCOMT24); The isoelectric points were distributed between 5.18 (VvCOMT22) and 6.23 (VvCOMT3), and the isoelectric points were all less than 7; The protein instability coefficients ranged from 27.21 (VvCOMT23) to 43.30 (VvCOMT5), among them 6 instability coefficients over 40 belonged to unstable proteins; All proteins with a hydrophilicity index between -0.158 (VvCOMT16) and 0.079 (VvCOMT7) were amphoteric proteins; The subcellular localization results showed that the final localization was in cytoplasm and extracellular space. By constructing the phylogenetic trees of the different species, it could be observed that the grape COMT gene was relatively conservative and highly similar in the evolutionary process, which was closely related to Arabidopsis and rice, and was farthest related to apple. From the prediction of the secondary structure of the proteins, it could be seen that the grape COMT gene family contained α-helix, β-corner, irregular curl and extended chain four configurations, the total proportion of α-helix and irregular curl was 70%. The Motif analysis yielded 19 more conservative motifs, and each VvCOMT gene was distributed with 4-14 motifs, the motif 6 present in all COMT genes, indicating that the motif 6 was highly conservative. At the same time, it was found that the COMT genes in the same group contained the same motifs. For example, the closely related VvCOMTs in Group I contained motifs 5, 7, 8, 17 and 19, while those in Group Ⅱ were different. Most COMT genes contained motifs 13, 14 and 16, while the VvCOMT4 only contained motifs 2, 3, 6 and 10. The difference in the motifs contained in different branches might be one of the reasons for functional differentiation in the evolution of the VvCOMT. It was found that 26 VvCOMT genes were irregularly distributed on 7 chromosome skeletons, among them the 12 chromosome had the most genes, including 13 VvCOMT genes. The promoter visualization showed that 227 cis-acting elements were found in the grape COMT gene family, including 184 hormone related elements, 19 fungal induction related elements and 29 defense and stress response related elements. By analyzing the expression of the COMT genes in the two cultivars after inoculation with downy mildew pathgen, it was found that 25 VvCOMTs were significantly upregulated under downy mildew stress in the resistant cultivar Moldova, with 42% of the COMT genes significantly upregulated at 48 h after inoculation, VvCOMT1, 5, 6, 7, 8, 9 and 19 were most significantly upregulated after inoculation with the pathogen in the susceptible cultivar Summer Black. Among them, the VvCOMT2 was not significantly upregulated in both cultivars, indicating that it did not respond to the infection of downy mildew. In addition, the expression of the COMT genes in resistant cultivar was significantly higher than that in susceptible cultivars, indicating that the VvCOMT genes played a certain role in the process of resistance to downy mildew. 【Conclusion】 This study showed that the grape COMT gene family responded to grape downy mildew and played an important role in the process of resistance to downy mildew infection.
Key words: Grape downy mildew; Caffeic acid O-methyltransferase; Biological analysis; Functional analysis
葡萄作为四大水果之一,因其营养丰富,适应性强,普遍受到消费者和生产者的青睐,在世界范围内被广泛栽培。然而,葡萄生产过程中经常遭受霜霉病、白腐病等真菌病害的危害,严重影响葡萄的产量和品质。葡萄霜霉病是全世界范围内严重危害葡萄的真菌病害之一[1],该病原菌主要以卵孢子形态在病残组织内越冬,5月份通过气流或雨滴溅散传播,典型的症状为叶片背面出现白色霜状霉层,即病菌的孢子囊和孢子梗,同时侵染嫩梢、卷须、叶柄和幼果等部位,严重危害葡萄的营养器官和生殖器官[2-3]。
木质素是一种多酚聚合物,被木纤维、其他维管束细胞和厚壁细胞包围[4-5],是植物中重要的次生物质,不仅可以增强植物细胞和组织的强度,有利于植物组织中的水分运输,同时也能提高植物抵抗病虫害的能力,其总量是仅次于纤维素的第二大有机物[6-7]。木质素的生物合成是苯丙氨酸或酪氨酸在一系列酶的催化下逐渐转化为木质素单体,继而形成木质素的过程[8-9],该过程由3个途径组成:苯丙烷途径、木质素合成的特定途径以及木质素单体向木质素的糖基化转运和聚合的途径。咖啡酸O-甲基转移酶(COMT)是苯丙烷代谢途径中重要的甲基化酶[10-11],COMT有多种功能,如催化咖啡酸的甲基化,5-羟基苯基醛生成阿魏酸、芥末醛等,还能催化S-腺苷L-蛋氨酸(SAM或AdoMet)的甲基基团形成阿魏酸和S-腺苷L-同型半胱氨酸(SAH或AdoHcy)来调节木质素的合成,且其N端在没有金属离子的环境下就能进行同源二聚化,除参与木质素合成外还在类黄酮和芥子酸酯等物质中发挥催化作用[12-13]。
前人研究表明,COMT基因家族包含多个成员,如毛杨中有25个[14],拟南芥和甘蓝型油菜中分别有14个[15]和42个[16]。在其他物种中也有相关的研究,如烟草[17]、燕麦[18]、松树[19]、水稻[20]、大麦[21]和蓝莓果实[22]。植物病原菌侵染和植株果实发育包含木质素的积累过程,而COMT基因已被证明在木质素积累过程中起着关键作用。Petitot等[23]发现非洲水稻COMT3在根结线虫侵染过程中表达量明显升高;Fornalé等[24]抑制玉米COMT基因的表达后发现其总木质素含量和S单位/G单位比降低;Wang等[25]过表达COMT-3D基因使得转基因小麦的耐病性与木质素含量得到了提高。因此,COMT基因对植物抵抗生物和非生物胁迫尤为重要。但在葡萄中尚未对COMT基因家族的特征和功能进行全面研究。笔者在本研究中参考乃国洁等[26]的生物信息学方法,鉴定了葡萄COMT基因家族,分析其蛋白质理化性质、染色体定位、保守结构域和基序分析,同时研究不同抗性葡萄品种COMT基因接种霜霉病的表达模式,旨在挖掘葡萄霜霉病响应的关键COMT基因,有利于进一步明确该基因家族在葡萄抗病反应过程中的作用,为葡萄抗病品种的选育奠定基础。
1 材料和方法
1.1 葡萄COMT基因的鉴定及理化性质分析
葡萄基因组数据库(v2.1)来自phytozome 13(https://Phytozome-next.igi.doe.gov),首先在拟南芥数据库(https://www.arabidopsis.org)搜索O-甲基转移酶基因,基于Pfam数据库的隐马尔可夫模型(PF00891),利用phytozome 13非冗余蛋白质数据库中BLASTP工具搜索比对,获取葡萄COMT家族候选基因,删除重复和冗余序列,确定VvCOMT基因家族。利用在线ExPASy(http://web.expasy.org/)和Plant-PLocserve(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)工具预测VvCOMT蛋白的理化性质,包括蛋白长度、分子质量、等电点等,并预测亚细胞定位,利用线上分析软件ProtParam(http://web.expasy.org/protparam)进行蛋白质二级结构预测。
1.2 系统发育树的构建及染色体定位
运用MEGA 11软件中的Clustal W程序将拟南芥、水稻、玉米、大豆和番茄等物种蛋白序列进行多序列比对,并通过邻接法(neighbour-joining,NJ)和最大似然法(maximum likelihood,ML)构建系统发育树,Bootstrap检验设定1000次重复,以评价系统发育树的统计可靠性。利用iTOL(http://iTOL.embl.de)在线软件对进化树进行美化。
为了解VvCOMTs基因在基因组内的分布,通过JGI数据库获得基因组注释文件中提供的位置信息,利用TBtools软件[27]将葡萄COMT基因定位到相应的染色体上。
1.3 VvCOMT家族基序与启动子顺式作用分析
为更好地理解和调控VvCOMTs的基因功能,利用在線程序MEME(v4.3)(http://meme.nbcr.net/meme/)分析VvCOMTs序列特征蛋白保守模块(motif),查找的motif数量设置为20,运行参数为默认。
从欧洲葡萄数据库中查找VvCOMTs起始密码子ATG上游2 kb的序列,提交使用在线程序PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测VvCOMT启动子区域的顺式作用元件,将获得的顺式作用元件提交至TBtools软件中进行启动子可视化。
1.4 VvCOMT基因家族成员在葡萄霜霉病中的表达分析
为了解VvCOMTs在抗霜霉病侵染中的作用,采集病叶制作霜霉病病原孢子(浓度为1×105个·mL-1),材料选择抗病品种摩尔多瓦和感病品种夏黑接种制备的霜霉病菌,接种后0、6、48、96、120 h分別采样,液氮速冻于-80 ℃保存。样品RNA提取采用试剂盒法(OMEGA,美国),利用UEIris RT mix with DNase(All-in-One)合成cDNA,使用实时荧光定量PCR(qRT-PCR)技术分析基因表达水平,荧光定量反应体系10 μL:TB Green?Premix Ex Taq?(TaKaRa,大连)5 μL,模板0.5 μL,上、下游引物1 μL,ddH2O 3.5 μL。荧光定量反应程序为:50 ℃ 2 min,95 ℃ 2 min,然后95 ℃ 15 s,60 ℃ 30 s,39次循环。Thresh值按PCR仪默认为30,分别记录每个反应荧光信号由本底进入指数增长阶段的拐点所对应的循环数(threshold cycle,Ct),然后用2-△△CT法[28]以未接种霜霉病菌的叶片为对照,对不同时间点VvCOMTs基因的相对表达量进行分析。所有样本使用3次生物重复进行分析。使用葡萄Actin作为内参引物,本研究中使用的所有引物均列于表1。
1.5 统计分析
采用SPSS Stantistics v.26.0软件对数据进行方差分析(ANOVA)。采用最小显著性差异(p<0.05)进行显著性分析。
2 结果与分析
2.1 葡萄COMT基因的鉴定和蛋白质理化性质分析
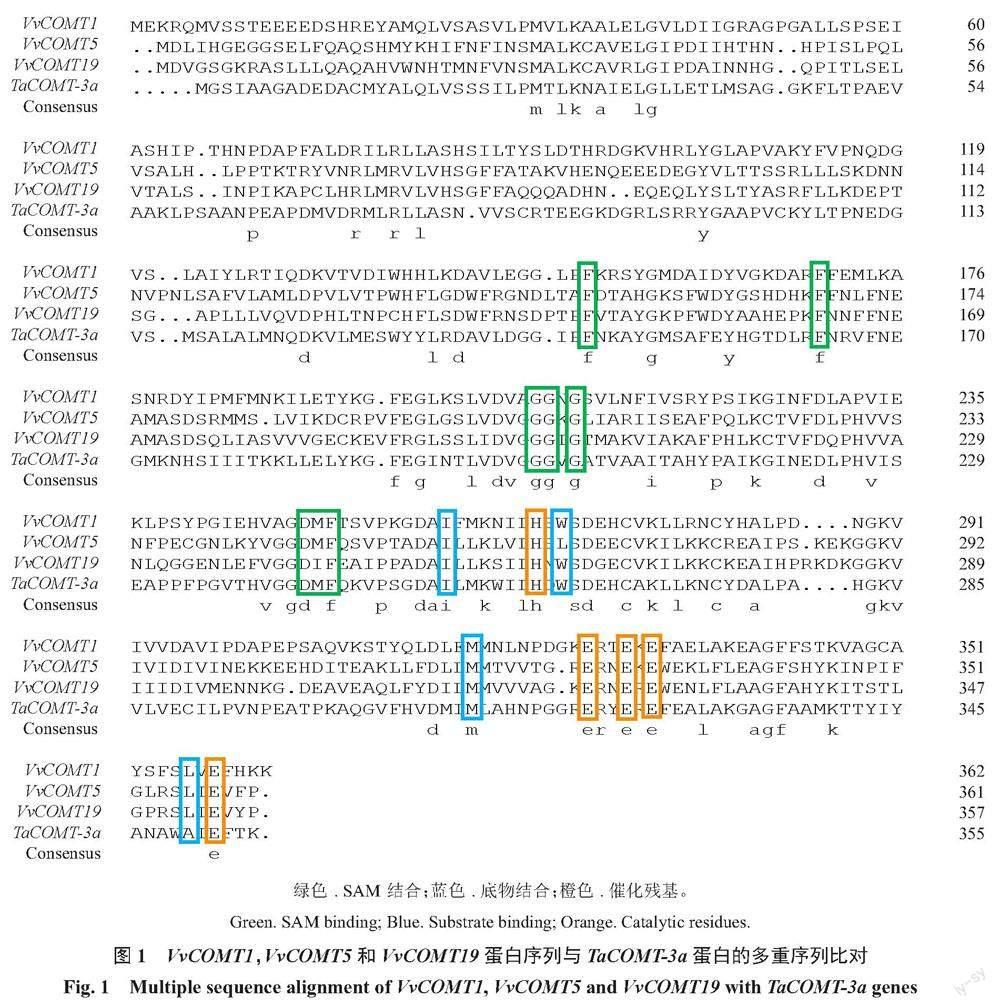
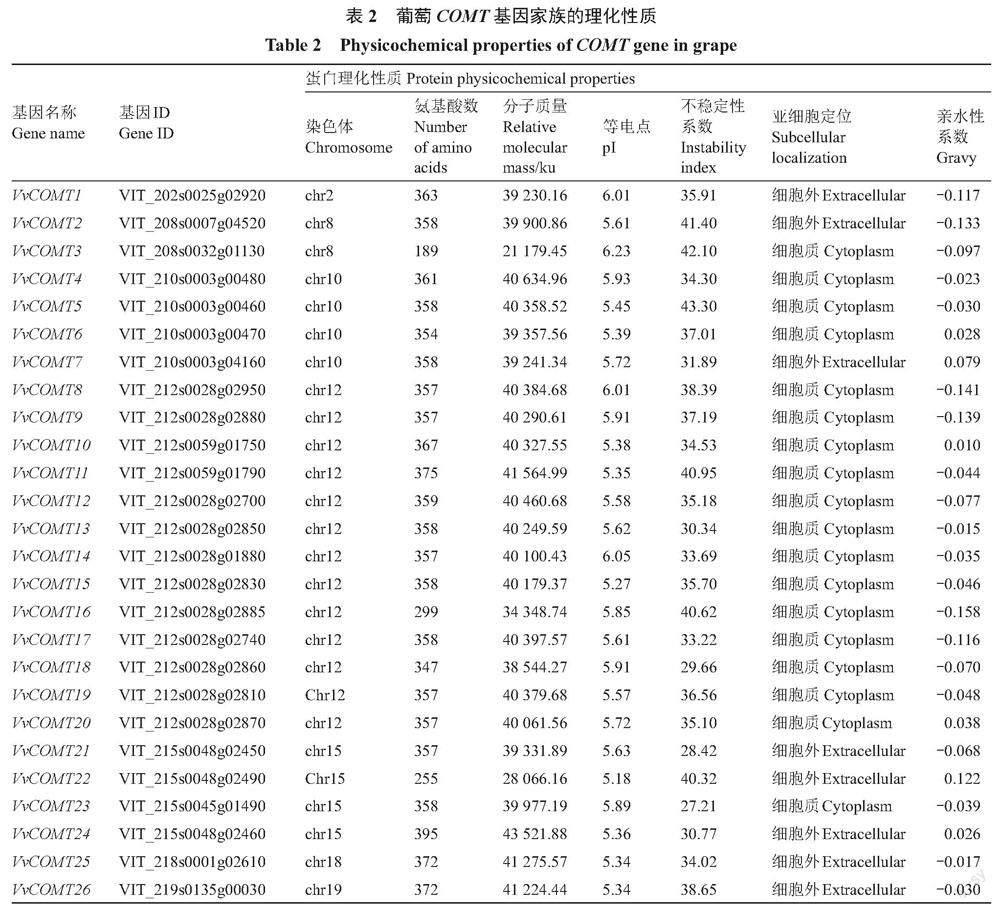
通过生物信息学分析获得26条VvCOMT基因,按照染色体位置分别命名为VvCOMT1~26。基因结构和保守结构域分析表明,26个葡萄COMT都具有一个名为Methyltransf_2结构域的C端催化结构域(PF00891),包括SAM/SAH结合袋和底物结合位点[29],SAM/SAH结合袋高度保守,而底物结合位点对不同组中的蛋白质具有特异性[30](图1)。利用ExPASy在线工具进行蛋白理化性质分析(表2),84.6%的葡萄COMT氨基酸长度超过300 aa,且氨基酸数分布在189~395个之间,其中VvCOMT24(VIT_215s0048g02460)序列最长,有395个氨基酸,VvCOMT3(VIT_208s0032g01130)序列最短,氨基酸数只有189。蛋白质相对分子质量在21 179.45(VvCOMT3)~43 521.88 Ku(VvCOMT24);等电点分布在5.18(VvCOMT22)~6.23(VvCOMT3)之间,且等电点都小于7;蛋白质不稳定系数在27.21(VvCOMT23)~43.30(VvCOMT5),其中6条不稳定系数大于40属于不稳定蛋白;亲水指数在-0.158(VvCOMT16)~0.079(VvCOMT7)之间均为两性蛋白;亚细胞定位结果显示26个COMT蛋白定位于细胞质和细胞外。
2.2 系统发育树的构建和蛋白质二级结构分析
为更好地了解葡萄与其他植物COMT的相似性和差异性,利用26条葡萄COMT蛋白与8条拟南芥、14条玉米、28条水稻、16条大豆和12条番茄共104条蛋白序列构建了系统发育树(图2)。VvCOMT的系统发育分析显示,26个VvCOMT蛋白序列可分为两组:GroupⅠ包含17个VvCOMT蛋白,其余9个VvCOMT蛋白属于Group Ⅱ。葡萄COMT基因家族成员呈现集中分布在2个类群之中,在进化过程中较保守,具有高度的相似性,其中与拟南芥和水稻亲缘关系较近,与玉米的亲缘关系最远,表明葡萄与玉米之间的COMT基因差异显著。
通过ProtParam在线分析工具预测葡萄COMT基因家族成员的二级结构(表3),葡萄COMT基因家族均含有α-螺旋、β-转角、无规则卷曲和延伸链4种构型,其中α-螺旋和无规则卷曲两种构型的总占比为70%,而β-转角与延伸链两种构型则只占总比的30%。
2.3 染色体定位
利用TBtools软件进行染色体定位分析,结果显示,26条基因在7条染色体骨架上呈无规则分布,且不同染色体骨架上的基因分布密度不同(图3),其中第12号染色体上基因分布最多,含有13条VvCOMT基因,第2、18和19号染色体上成员最少,各含有1条VvCOMT基因。
2.4 基因结构与基序分析
利用MEME在线工具分析,发现葡萄COMT基因存在19个较为保守的motif(图4),每条VvCOMT基因分布4~14个motif,其中motif 6存在于所有的COMT基因,表明motif 6具有很强的保守性。同时发现同一类群的COMT基因包含的motif相同,如Group Ⅰ中亲缘关系较近的VvCOMT都含有motif 5、7、8、17和19,Group Ⅱ与之不同,大多数COMT基因都含有motif 13、14和16,VvCOMT4中只含有motif 2、3、6、10基序,不同的分支所包含基序的不同可能是VvCOMT进化过程中发生功能分化的原因之一。
根据VvCOMT系统发育关系,26个基因被分为3个亚组(图4),其中Ⅰ组与Ⅱ组与COMT基因外显子-内含子结构相似,都含有2个外显子和1个内含子,长度在302~775 bp之间,而Ⅲ组则包含3~4个外显子,且在同一进化枝中的外显子数量接近,说明系统发育树的可靠性。此外,26个COMT基因中形成9个旁系同源对,其中VvCOMT4/5/6,VvCOMT8/9及VvCOMT10/11步长值高达100。
2.5 启动子顺式作用元件分析
通过对葡萄COMT基因家族成员上游2000 bp启动子区的顺式作用元件进行分析,笔者发现葡萄26条COMT基因中共存在243个顺式作用元件,其中192个激素相关元件(其中乙烯相关元件109个,脱落酸相关元件39个,水杨酸相关元件21个,茉莉酸相关元件18个,赤霉素相关元件5个),真菌诱导相关元件20个,防御和应激反应相关元件31个(图5)。说明葡萄COMT基因可能参与激素响应和逆境胁迫响应过程。
2.6 葡萄COMT基因家族成员接种霜霉病的表达分析
为探究葡萄COMT基因对霜霉病的响应,利用荧光定量PCR技术,分析抗病品种摩尔多瓦和感病品种夏黑在接种霜霉病后COMT基因家族成员的表达量。由图6和图7可以看出,在抗病品种摩尔多瓦中,25个VvCOMT在霜霉病胁迫下均显著上调,其中42%的COMT基因在接种后的48 h即显著上调;而在感病品种夏黑中,VvCOMT1、2、、10、15、26和VvCOMT27在接种后病原菌后出现显著下调,63%的COMT基因在接种后的24 h出现显著上调,其中VvCOMT2在2个品种中均无显著上调,表明其不响应霜霉病菌的侵染。此外,COMT基因在抗病品种的表达量显著高于感病品种,GroupⅠ中VvCOMT5/6/8/9和19在抗病品种中分别比感病品种高出13、15、21、120和7倍;Group Ⅱ中VvCOMT1和VvCOMT7在抗病品种中分别比感病品种高出102和3580倍;Ⅲ家族成员VvCOMT25与VvCOMT26比较特殊,均显著下调。综合COMT基因在抗感品种中的表达,VvCOMT1、5、6、7、8、9和VvCOMT19上调最为显著,其最有可能在葡萄抵抗霜霉病菌胁迫过程中发挥着更重要的作用。
3 讨 论
植物O-甲基转移酶(OMTs)构成一大类酶,其中Ⅰ型OMT形成功能发散的基团,并使多种底物(如类黄酮,生物碱和二苯乙烯)甲基化,主要以COMT为代表[31]。在植物木质素生物合成、抵御病原菌侵染和抗逆胁迫中发挥重要作用。本研究基于phytozome 13数据库,利用拟南芥COMT基因对葡萄COMT基因家族进行了鉴定与分析,确定了26个VvCOMT基因,所有基因均包含咖啡酸辅酶A-O-甲基转移酶结构域,系统进化分析结果与基因结构及保守域分析结果一致,具有相同数量的内含子和保守基序的基因家族成员优先聚为一类,这与先前的研究结果一致[32],此外,各个亚类基因家族成员的蛋白质理化性质存在差异,本研究得到的26个COMT基因二级结构同拟南芥COMT基因家族的蛋白质组成和结果相似,其中α-螺旋和无规则卷曲比例最高,这与玉米COMT基因二级结构相同[33]。启动子是RNA聚合酶识别、结合和开始转录的一段DNA序列,它含有RNA聚合酶特异性结合和转录起始所需的保守序列,关系到基因的时空表达,是基因的开关,分析其启动子顺式作用元件有助于推测基因的潜在功能。VvCOMT基因的启动子中含有许多响应激素调控的作用元件(TGA-elemet生长素响应元件,TCA-elemet响应水杨酸响应元件,ABRE脱落酸响应元件,TGACG-motif茉莉酸甲酯响应元件),防御和应激反应作用元件提示基因在应对胁迫方面有重大作用,如VvCOMT1、5、13、14、19、20和VvCOMT26都含有防御和应激反应相关元件(TC-rich)。启动子分析结果表明,COMT基因家族作用于葡萄的抗病虫、抗逆等方面,这与其他物种中COMT基因家族的研究结果相一致[34-35]。
据报道,COMT与COMT-like基因在植物次生物质合成和抗逆胁迫反应中起着关键性作用[36-37]。单木质素生物合成是细胞壁贴合(cell wall apposition)过程中的关键,也是植物抵御病原菌的首要防线之一[38]。这一理论在其他物种中也得以验证,小麦中COMT-3D基因过表达可以提高其对纹枯病的抗性[25],增加木质素的积累;棉花中n-乙酰转移酶1(GhSNAT1)和咖啡酸O-甲基转移酶(GhCOMT)沉默导致褪黑素的生物合成减少,从而影响木质素和棉酚的合成,降低了对棉花黄萎病的抗性[39]; COMT的表达量下调降低了6个月龄的杨树中木质素的含量水平[40];水稻中咖啡酸O-甲基转移酶的过表达通过5-甲氧基色胺途径同样也增加了褪黑素的产生,以此提高植物抗性[41]。同样的,在VvCOMT基因家族在受到葡萄霜霉病侵染时,抗病品种COMT基因家族除VvCOMT2外均在接种霜霉病后48 h出现显著上调,而在感病品种中只有63%的COMT基因在接种后的24 h内出现短时上调,且抗病力不同的品種,其表达量和表达模式也不同,摩尔多瓦多在6~48 h之间显著表达,而夏黑则是24 h后表达量提高,且抗病品种中96%的基因表达量都高于感病品种,说明在抗病品种抵御霜霉病时COMT基因发挥重要作用,这与笔者先前预期的结果一致。植物生长发育过程中不会像动物拥有体细胞适应性免疫系统能够主动避开病原微生物和病虫害,只能依靠植物合成的化学成分和本身的一些结构作为屏障[42-43]。而在葡萄中,不同品种的抗霜霉病程度不一样,其COMT基因家族的表达量也不一样,结果中显示VvCOMT1、5、6、7、8、9和VvCOMT19基因在抗病品种摩尔多瓦中表达量显著高于感病品种夏黑,同时发现VvCOMT1、5、19启动子区域中包含抗防御和应激反应响应元件,可以证明这些基因在葡萄抗霜霉病侵染中发挥重要作用,后期可对这几个基因进行过表达处理,获得阳性转基因苗,以期挖掘出VvCOMT基因家族更多的潜在功能,为葡萄抗病品种的培育做出贡献。
4 结 论
笔者在本研究中鉴定了葡萄26个COMT基因家族成员,在接种霜霉病后均有表达,尤其在抗病品种中表达较显著,推测COMT基因在葡萄抗霜霉病侵染过程中发挥重要的作用,可为下一步研究其在抗病过程中的分子功能和在基因中的调控奠定基础。
参考文献References:
[1] 王璐璐,孙云开,徐旋,周洁,王清明,祝学珍. 葡萄霜霉病的发病规律和防治研究进展[J]. 园艺与种苗,2022,42(9):31-33.
WANG Lulu,SUN Yunkai,XU Xuan,ZHOU Jie,WANG Qingming,ZHU Xuezhen. Research progress on pathogenesis and control of grape downy mildew [J]. Horticulture & Seed,2022,42(9):31-33.
[2] 李卓,周婷婷,楊超,李映程,孙琦,任毓忠,赵宝龙,张莉,李国英. 葡萄霜霉病菌侵染抗病和感病品种过程的组织学观察[J]. 园艺学报,2017,44(5):861-870.
LI Zhuo,ZHOU Tingting,YANG Chao,LI Yingcheng,SUN Qi,REN Yuzhong,ZHAO Baolong,ZHANG Li,LI Guoying. Histological studies on the infection processs of the grape downy mildew between susceptible cultivar and resistant cultivar[J]. Acta Horticulturae Sinica,2017,44(5):861-870.
[3] 杜兴兰. 葡萄霜霉病和白粉病生物防治的研究[D]. 保定:河北农业大学,2008.
DU Xinglan. Biological control of grape downy mildew and powdery mildew[D]. Baoding:Hebei Agricultural University,2008.
[4] GUO W F,JIN L,MIAO Y H,HE X,HU Q,GUO K,ZHU L F,ZHANG X L. An ethylene response-related factor,GbERF1-like,from Gossypium barbadense improves resistance to Verticillium dahliae via activating lignin synthesis[J]. Plant Molecular Biology,2016,91(3):305-318.
[5] BOERJAN W,RALPH J,BAUCHER M. Lignin biosynthesis[J]. Annual Review of Plant Biology,2003,54(1):519-546.
[6] VANHOLME R,DE MEESTER B,RALPH J,BOERJAN W. Lignin biosynthesis and its integration into metabolism[J]. Current Opinion in Biotechnology,2019,56:230-239.
[7] LEWIS N G,YAMAMOTO E. Lignin:Occurrence,biogenesis and biodegradation[J]. Annual Review of Plant Physiology and Plant Molecular Biology,1990,41:455-496.
[8] ROJE S. S-Adenosyl-L-methionine:Beyond the universal methyl group donor[J]. Phytochemistry,2006,67(15):1686-1698.
[9] LAM K C,IBRAHIM R K,BEHDAD B,DAYANANDAN S. Structure,function,and evolution of plant O-methyltransferases[J]. Genome,2007,50(11):1001-1013.
[10] LU S W,ZHUGE Y X,HAO T Y,LIU Z J,ZHANG M W,FANG J G. Systematic analysis reveals O-methyltransferase gene family members involved in flavonoid biosynthesis in grape[J]. Plant Physiology and Biochemistry,2022,173:33-45.
[11] JOSHI C P,CHIANG V L. Conserved sequence motifs in plant S-adenosyl-L-methionine-dependent methyltransferases[J]. Plant Molecular Biology,1998,37(4):663-674.
[12] BERIM A,GANG D R. Methoxylated flavones:Occurrence,importance,biosynthesis[J]. Phytochemistry Reviews,2016,15(3):363-390.
[13] BENNETT M R,SHEPHERD S A,CRONIN V A,MICKLEFIELD J. Recent advances in methyltransferase biocatalysis[J]. Current Opinion in Chemical Biology,2017,37:97-106.
[14] SHI R,SUN Y H,LI Q Z,HEBER S,SEDEROFF R,CHIANG V L. Towards a systems approach for lignin biosynthesis in Populus trichocarpa:Transcript abundance and specificity of the monolignol biosynthetic genes[J]. Plant and Cell Physiology,2010,51(1):144-163.
[15] MOINUDDIN S G A,JOURDES M,LASKAR D D,KI C,CARDENAS C L,KIM K W,ZHANG D Z,DAVIN L B,LEWIS N G. Insights into lignin primary structure and deconstruction from Arabidopsis thaliana COMT (caffeic acid O-methyl transferase) mutant Atomt1[J]. Organic & Biomolecular Chemistry,2010,8(17):3928-3946.
[16] LI W,LU J X,LU K,YUAN J L,HUANG J H,DU H,LI J N. Cloning and phylogenetic analysis of Brassica napus L. caffeic acid O-methyltransferase 1 gene family and its expression pattern under drought stress[J]. PLoS One,2016,11(11):e0165975.
[17] PELLEGRINI L,GEOFFROY P,FRITIG B,LEGRAND M. Molecular cloning and expression of a new class of ortho-diphenol-O-methyltransferases induced in tobacco (Nicotiana tabacum L.) leaves by infection or elicitor treatment[J]. Plant Physiology,1993,103(2):509-517.
[18] HEATH R,HUXLEY H,STONE B,SPANGENBERG G. cDNA cloning and differential expression of three caffeic acid O-methyltransferase homologues from perennial ryegrass (Lolium perenne)[J]. Journal of Plant Physiology,1998,153(5/6):649-657.
[19] CHUNG N,ZHANG X D,KREAMER A,LOCCO L,KUAN P F,BARTZ S,LINSLEY P S,FERRER M,STRULOVICI B. Median absolute deviation to improve hit selection for genome-scale RNAi screens[J]. Journal of Biomolecular Screening,2008,13(2):149-158.
[20] CHOI G H,LEE H Y,BACK K. Chloroplast overexpression of rice caffeic acid O-methyltransferase increases melatonin production in chloroplasts via the 5-methoxytryptamine pathway in transgenic rice plants[J]. Journal of Pineal Research,2017,63(1):e12412.
[21] DALY P,MCCLELLAN C,MALUK M,OAKEY H,LAPIERRE C,WAUGH R,STEPHENS J,MARSHALL D,BARAKATE A,TSUJI Y,GOEMINNE G,VANHOLME R,BOERJAN W,RALPH J,HALPIN C. RNAi-suppression of barley caffeic acid O-methyltransferase modifies lignin despite redundancy in the gene family[J]. Plant Biotechnology Journal,2019,17(3):594-607.
[22] LIU Y S,WANG Y Z,PEI J B,LI Y D,SUN H Y. Genome-wide identification and characterization of COMT gene family during the development of blueberry fruit[J]. BMC Plant Biology,2021,21(1):5.
[23] PETITOT A S,KYNDT T,HAIDAR R,DEREEPER A,COLLIN M,DE ALMEIDA ENGLER J,GHEYSEN G,FERNANDEZ D. Transcriptomic and histological responses of African rice (Oryza glaberrima) to Meloidogyne graminicola provide new insights into root-knot nematode resistance in monocots[J]. Annals of Botany,2017,119(5):885-899.
[24] FORNAL? S,SONBOL F M,MAES T,CAPELLADES M,PUIGDOM?NECH P,RIGAU J,CAPARR?S-RUIZ D. Down-regulation of the maize and Arabidopsis thaliana caffeic acid O-methyl-transferase genes by two new maize R2R3-MYB transcription factors[J]. Plant Molecular Biology,2006,62(6):809-823.
[25] WANG M X,ZHU X L,WANG K,LU C G,LUO M Y,SHAN T L,ZHANG Z Y. A wheat caffeic acid 3-O-methyltransferase TaCOMT-3D positively contributes to both resistance to sharp eyespot disease and stem mechanical strength[J]. Scientific Reports,2018,8:6543.
[26] 乃国洁,卢世雄,马维峰,李艳梅,陈佰鸿,毛娟. 葡萄EIN3/EIL转录因子家族成员鉴定及表达特征分析[J]. 果树学报,2021,38(6):856-870.
NAI Guojie,LU Shixiong,MA Weifeng,LI Yanmei,CHEN Baihong,MAO Juan. Genome-wide identification and expression characteristic analysis of EIN3/EIL transcription factor family in grape[J]. Journal of Fruit Science,2021,38(6):856-870.
[27] CHEN C J,CHEN H,ZHANG Y,THOMAS H R,FRANK M H,HE Y H,XIA R. TBtools:An integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant,2020,13(8):1194-1202.
[28] LIVAK K J,SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2?ΔΔCT method[J]. Methods,2001,25(4):402-408.
[29] RAMMESMAYER G,PICHORNER H,ADAMS P,JENSEN R G,BOHNERT H J. Characterization of IMT1,myo-inositol O-methyltransferase,from Mesembryanthemum crystallinum[J]. Archives of Biochemistry and Biophysics,1995,322(1):183-188.
[30] DO C T,POLLET B,TH?VENIN J,SIBOUT R,DENOUE D,BARRI?RE Y,LAPIERRE C,JOUANIN L. Both caffeoyl Coenzyme A 3-O-methyltransferase 1 and caffeic acid O-methyltransferase 1 are involved in redundant functions for lignin,flavonoids and sinapoyl malate biosynthesis in Arabidopsis[J]. Planta,2007,226(5):1117-1129.
[31] IBRAHIM R K,BRUNEAU A,BANTIGNIES B. Plant O-methyltransferases:Molecular analysis,common signature and classification[J]. Plant Molecular Biology,1998,36(1):1-10.
[32] 包穎亮. 构树咖啡酸-O-甲基转移酶基因的克隆及生物信息学分析[D]. 呼和浩特:内蒙古农业大学,2018.
BAO Yingliang. Cloning and bioinformatics analysis of caffeic acid O-methyl-transferase gene from Broussonetia papyrifera[D]. Hohhot:Inner Mongolia Agricultural University,2018.
[33] 徐碧莹,张敏艳,黄伟鹏,沙哈尼,吴委林,郑大浩. 玉米O-甲基转移酶(ZmOMT)基因家族的生物信息学分析[J/OL]. 分子植物育种,2021:1-12. [2021-11-05]. https://kns.cnki.net/kcms/detail/46.1068.s.20211104.1914.021.html.
XU Biying,ZHANG Minyan,HUANG Weipeng,SHA Hani,WU Weilin,ZHENG Dahao. Bioinformatics analysis of maize O-methyltransferase (ZmOMT) gene family[J/OL]. Molecular Plant Breeding, 2021:1-12. [2021-11-05]. https://kns.cnki.net/kcms/detail/46.1068.s.20211104.1914.021.html.
[34] WU C C,ZUO D Y,XIAO S P,WANG Q L,CHENG H L,LV L M,ZHANG Y P,LI P B,SONG G L. Genome-wide identification and characterization of GhCOMT gene family during fiber development and Verticillium wilt resistance in cotton[J]. Plants,2021,10(12):2756.
[35] 陈安琪. 川芎咖啡酸-O-甲基转移酶(LcCOMT)的功能鉴定与应用研究[D]. 成都:西南交通大学,2021.
CHEN Anqi. Functional identification and application of Ligusticum chuanxiong caffcie acid-O-methyltransferase (LcCOMT)[D].
Chengdu:Southwest Jiaotong University,2021.
[36] LEE J E,VOGT T,HAUSE B,L?BLER M. Methyl jasmonate induces an O-methyltransferase in Barley[J]. Plant and Cell Physiology,1997,38(7):851-862.
[37] TOQUIN V,GRAUSEM B,GEOFFROY P,LEGRAND M. Structure of the tobacco caffeic acid O-methyltransferase (COMT) II gene:Identification of promoter sequences involved in gene inducibility by various stimuli[J]. Plant Molecular Biology,2003,52(3):495-509.
[38] COLLINGE D B. Cell wall appositions:The first line of defence[J]. Journal of Experimental Botany,2009,60(2):351-352.
[39] LI C,HE Q L,ZHANG F,YU J W,LI C,ZHAO T L,ZHANG Y,XIE Q W,SU B R,MEI L,ZHU S J,CHEN J H. Melatonin enhances cotton immunity to Verticillium wilt via manipulating lignin and gossypol biosynthesis[J]. The Plant Journal,2019,100(4):784-800.
[40] BARAKAT A,CHOI A,YASSIN N B M,PARK J S,SUN Z C,CARLSON J E. Comparative genomics and evolutionary analyses of the O-methyltransferase gene family in Populus[J]. Gene,2011,479(1/2):37-46.
[41] BYEON Y,CHOI G H,LEE H Y,BACK K. Melatonin biosynthesis requires N-acetylserotonin methyltransferase activity of caffeic acid O-methyltransferase in rice[J]. Journal of Experimental Botany,2015,66(21):6917-6925.
[42] HO-YUE-KUANG S,ALVARADO C,ANTELME S,BOUCHET B,C?ZARD L,LE BRIS P,LEG?E F,MAIA-GRONDARD A,YOSHINAGA A,SAULNIER L,GUILLON F,SIBOUT R,LAPIERRE C,CHATEIGNER-BOUTIN A L. Mutation in Brachypodium caffeic acid O-methyltransferase 6 alters stem and grain lignins and improves straw saccharification without deteriorating grain quality[J]. Journal of Experimental Botany,2016,67(1):227-237.
[43] 李偉,熊谨,陈晓阳. 木质素代谢的生理意义及其遗传控制研究进展[J]. 西北植物学报,2003,23(4):675-681.
LI Wei,XIONG Jin,CHEN Xiaoyang. Advances in the research of physiological significances and genetic regulation of lignin metabolism[J]. Acta Botanica Boreali-Occidentalia Sinica,2003,23(4):675-681.
收稿日期:2023-04-04 接受日期:2023-05-24
基金项目:河南省科技攻关项目(222102110199、232102111090)
作者简介:许雯雯,女,硕士,研究方向为果树种质资源与遗传育种。Tel:15136767398,E-mail:xuwen199810@163.com
通信作者 Author for correspondence. Tel:13569430110,E-mail:liguirong10@163.com
猜你喜欢
现代交际(2016年20期)2017-02-15 18:33:15
科技资讯(2016年19期)2016-11-15 10:39:12
电子技术与软件工程(2016年18期)2016-11-14 00:49:04
戏剧之家(2016年20期)2016-11-09 23:08:07
老区建设(2016年4期)2016-09-28 05:09:17
电脑知识与技术(2016年16期)2016-07-22 19:10:51
科技视界(2016年13期)2016-06-13 21:00:36
戏剧之家(2016年6期)2016-04-16 11:52:45
科技视界(2016年4期)2016-02-22 20:53:20
科技资讯(2015年8期)2015-07-02 18:49:25
