
近百年来异龙湖有机碳和无机碳埋藏响应水体富营养化的协同变化特征
2023-10-26 08:09:30陈光杰孔令阳黄林培杨关绍韩桥花
中国环境科学 2023年10期
李 平,陈光杰,孔令阳,黄林培,王 旭,杨关绍,索 旗,李 静,王 露,韩桥花,周 起,郭 雯,李 蕊,张 涛
近百年来异龙湖有机碳和无机碳埋藏响应水体富营养化的协同变化特征
李 平1,2,陈光杰1*,孔令阳1,黄林培1,王 旭1,杨关绍1,索 旗1,李 静1,王 露1,韩桥花1,周 起1,郭 雯1,李 蕊1,张 涛1
(1.云南师范大学地理学部,高原地理过程与环境变化云南省重点实验室,云南 昆明 650500;2.西昌学院资源与环境学院,四川 西昌 615013)
湖泊碳埋藏模式及其动态变化对陆地碳汇通量与全球碳收支估算具有重要影响,然而现有湖泊碳埋藏评估主要基于有机碳的埋藏模式,缺乏对无机碳埋藏以及有机碳-无机碳协同变化过程的系统评估.本文通过对云南岩溶区异龙湖开展沉积物岩芯样品的多指标分析和湖盆3个点位的记录对比,结合210Pb、137Cs年代学序列,识别了1900年以来异龙湖有机碳和无机碳埋藏通量的变化历史和空间特征,并甄别了二者协同变化的主要特征及其转化路径.结果表明,近百年来流域开发导致的营养盐长期输入(如沉积物氮和磷的持续富集)显著提高了异龙湖的内源初级生产力,表现为1900~2020年间沉积物藻类色素含量持续增加,蓝藻叶黄素浓度由~7.02μg/g显著增加到~38.99μg/g.同期有机碳和无机碳的埋藏通量均出现持续的上升趋势,且两者的同步变化特征显著(=0.98~0.99,<0.001).回归分析表明,藻类生物量是碳埋藏通量上升的主要驱动因子(2=0.33~0.91,<0.001),指示了藻类光合作用以及“生物碳泵”效应对碳埋藏过程的驱动作用.在流域地表物质输入强度总体下降(如磁化率信号和沉积物C:N比值的减小)的背景下,藻类生物量和内源有机碳的持续上升导致了有机质矿化过程中自生碳酸盐的转化作用明显增加(如无机碳δ13C信号的持续下降),进而促进了有机碳与无机碳埋藏通量的协同增长.上述结果表明,水体富营养化过程促进形成了有机碳和无机碳埋藏通量同步增加的长期模式,加强了异龙湖有机碳和无机碳埋藏过程的耦合作用.因此在人类活动与气候变化持续的背景下,对岩溶地区湖泊碳埋藏模式的科学评估需要关注无机碳埋藏过程及其对陆地碳汇通量的动态影响.
富营养化;碳埋藏;无机碳;协同变化;异龙湖
湖泊碳埋藏作为全球陆地碳收支的重要组成,在区域乃至全球碳汇评估中具有重要作用[1-3].全球湖泊面积仅占海洋面积的0.8%,其碳埋藏通量却达到了海洋的45%[4-5],湖泊碳埋藏的变化已成为评估陆地剩余碳汇的重要研究内容[1,6-8].在区域增温和流域开发背景下,近百年来全球湖泊的碳埋藏速率明显上升[9-12],显示了其对环境压力呈现持续变化的响应特征.尤其是在水体富营养化的背景下,部分湖泊可以通过埋藏更多的有机碳和减少脱气可转变为明显的大气CO2汇[13-16].因此在未来气候变化和人类活动持续的场景下,对湖泊碳埋藏模式和变化过程的系统研究,将有助于精确评估湖泊生态系统在全球碳收支中的作用及其演化趋势.
湖泊碳埋藏包括有机碳和无机碳两大组成部分.在岩溶地区以及干旱、半干旱地区,流域输出影响下无机碳在沉积物中占有较高比重,且无机碳的占比可能是比有机碳更为重要的组分[2,17-20].而已有的湖泊碳埋藏估算主要基于有机碳埋藏,对无机碳埋藏通量和无机碳库储量缺乏系统评估和科学估算.世界范围内,碳酸盐岩分布约占陆地面积的15%,而在我国则超过了陆地面积的26%[21].岩溶区湖泊通过水生植物的光合作用和“生物碳泵”效应,以有机碳和无机碳埋藏的方式,实现了对流域碳酸盐风化产生的溶解性无机碳(DIC)的长期固定,因此被认为是重要的碳汇过程[8,22].在区域增温的背景下,增强的有机质分解和矿化作用可以促进硬水湖泊碳酸盐的形成与无机碳埋藏量的增加[23-27].在流域开发持续的影响下地表溶蚀作用减弱、外源无机碳输入降低,而水体的富营养化不仅导致内源有机质产量和有机碳埋藏量的持续增加,同时更多的有机质可以分解并形成无机碳埋藏,均可以导致有机碳和无机碳埋藏通量的协同模式出现显著变化[19,28].在湖泊的富营养化过程中水体的理化特征变化显著(如水体pH值、溶解氧水平),可以促进内源有机质的矿化作用与自生碳酸盐的溶蚀作用[29-31],也可能维持或减少内源有机碳向自生碳酸盐的转化[32],从而显著改变岩溶区湖泊碳埋藏通量和碳汇过程.因此对有机碳-无机碳协同变化过程的认识有助于科学解释湖泊碳埋藏通量的变化趋势并识别有机碳-无机碳转换的耦合作用.然而,现有研究对水体富营养化过程中有机碳-无机碳的协同变化机制及其对湖泊碳埋藏的影响还缺乏系统性理解.基于以上背景,开展岩溶地区湖泊无机碳埋藏通量以及有机碳-无机碳协同变化的研究,将有助于科学评估气候变化和人类活动影响下的湖泊碳埋藏变化及有机碳-无机碳转化过程的耦合作用.
1 材料与方法
1.1 研究区概况
异龙湖(23°39¢~23°42¢N, 102°30¢~102°38¢E)位于云南省红河州石屏县境内(图1),为云南省九大高原湖泊之一.异龙湖由晚新生代石屏断裂与大桥-石屏-新街主断层相互作用断陷和溶蚀形成.流域内呈北西向带状断陷溶蚀堆积地貌,碳酸盐岩分布占流域面积的约30%.异龙湖流域属亚热带高原季风气候,年平均气温约18℃,自1950年以来流域气温呈逐渐上升趋势(图2(a)).多年平均降水量约919.9mm,多年平均蒸发量1034.5mm[46],近几十年来降水呈明显的年际波动变化(图2(b)).流域干湿季分明,5~10月降水量约占全年降水量的80%(图1).
异龙湖平均水深约2.5m,最大水深约5.2m(2021年),换水周期约2.4a[44].湖盆长约13.1km,最大宽约3.2km,水域面积约30km2,蓄水量近1.05×108m3.湖底标高1406.20m,最大运行水位1414.17m(1985国家基准高程).流域集水面积约326km2[45],年入湖水量约0.84×108m3,其中59.83%来自地表径流,15.59%由地下水补给,湖面降水占比24.64%[46].主要入湖径流分布在湖西北,包括城河、城南河和城北河,3条河流约贡献了入湖径流量的85%.异龙湖出水口在湖东北面海河口,年出湖水量主要受水量调度控制.
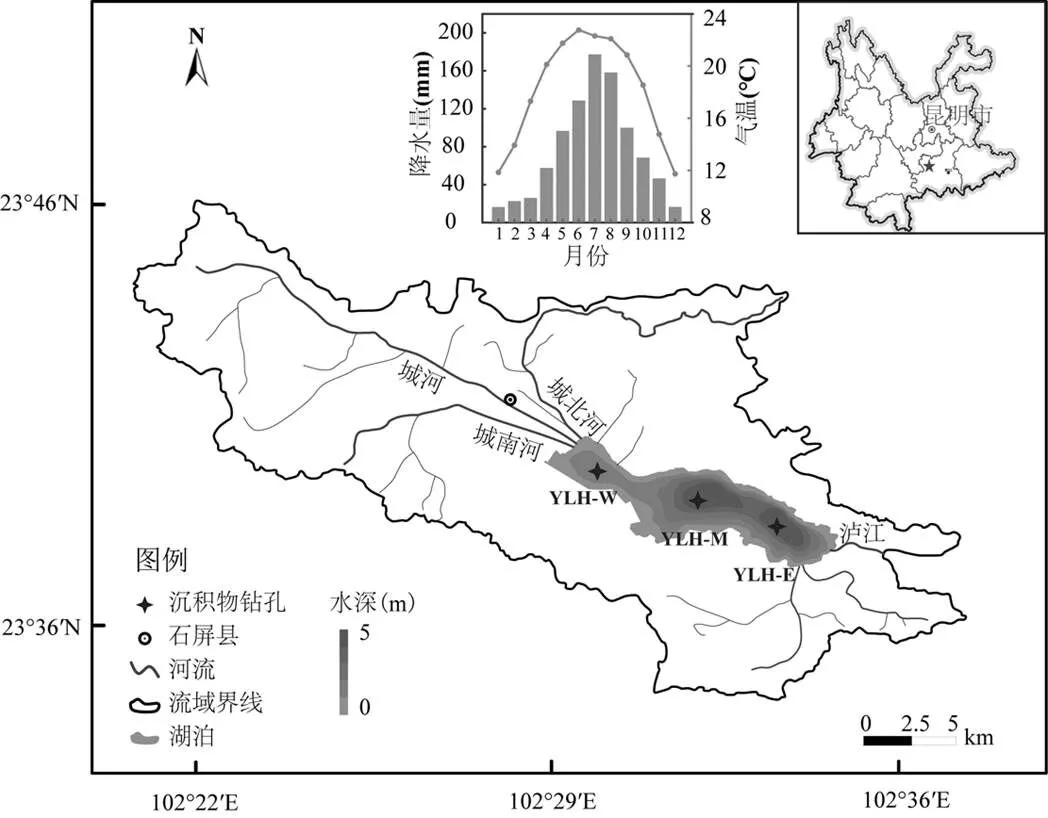
图1 研究区位置概况以及采样点分布
近几十年来受人类活动影响,异龙湖水质经历了逐步恶化的过程.20世纪50年代,异龙湖沉水植物等高等水生植物繁茂,海菜花((.))、轮藻()和蓖齿眼子菜()占优.50~70年代两次大规模放水的围湖造田活动和80~90年代大规模网箱养鱼的渔业开发对异龙湖水质产生了重要影响.至1983年左右指示清洁水体的海菜花在异龙湖绝迹,1990年代开始蓝藻逐步占优[47],并在2009年转变为藻型湖泊[48].近年来异龙湖水生高等植物以挺水植物(芦苇()、香蒲()、荷花())和漂浮植物为主[49],主要分布在西部湖盆沿岸和中部湖盆的北岸.
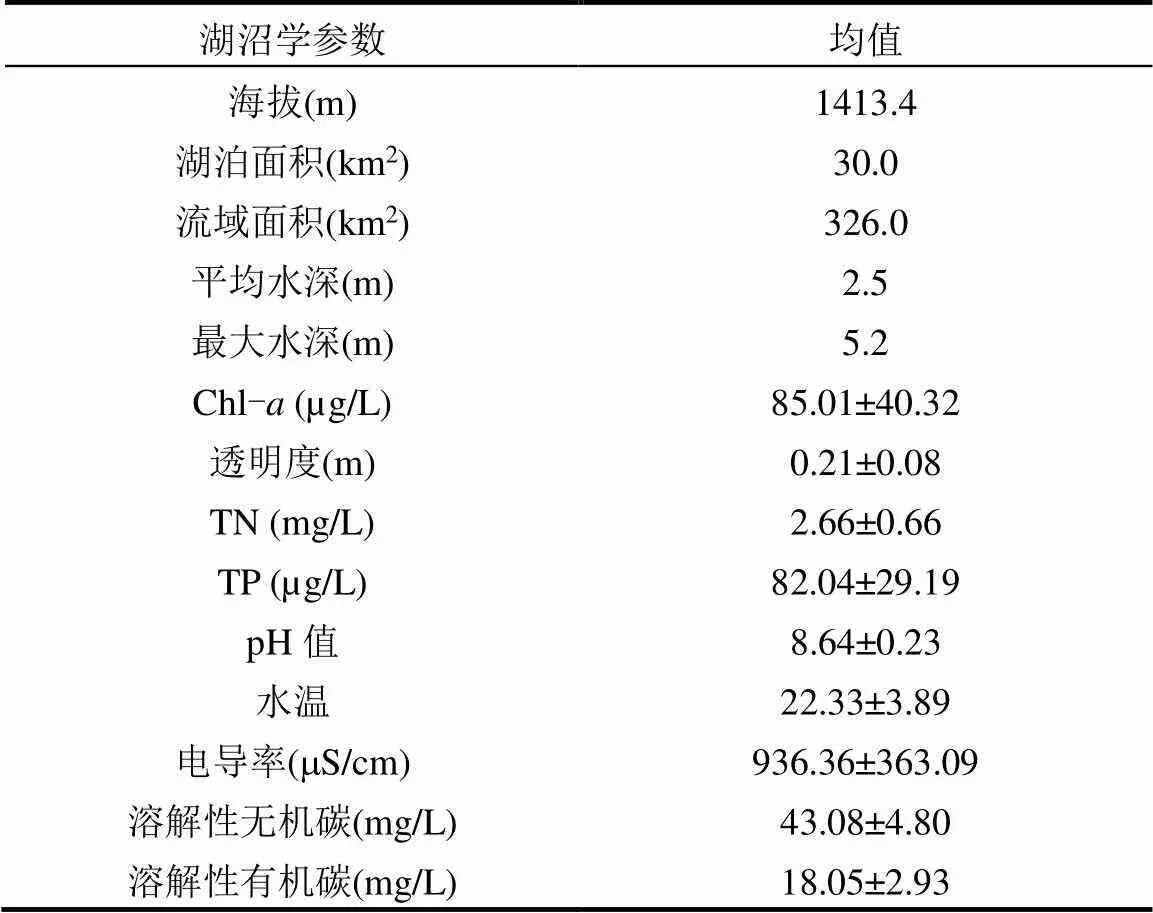
表1 异龙湖主要湖沼学特征
注:数据来源为2020~2022年的异龙湖野外监测.
监测数据显示,异龙湖目前为劣五类水质的重富营养水平水体(表1).2021年,湖水透明度约0.2m,pH值8.64,叶绿素(Chl-)约为85.01μg/L,水体TP和TN浓度分别为82.04μg/L和2.66mg/L(表1).长期调查数据显示,流域开发和农业扩展导致了近几十年来化肥施用量激增以及水体营养盐富集(图2(d),(e)).其中,农业面源污染和生活污水是入湖营养盐的主要来源,通过3条主要入湖河流贡献的入湖营养盐超过了总入湖量的50%,其中总氮(TN)和总磷(TP)分别达到每年360t和26.67t[50].
1.2 样品采集与年代测定
利用活塞重力取样器(HTH,瑞典)于2020年6月在异龙湖西部(23°41'10'', 102°31'29'')、中部(23°40'16", 102°34'16'')和东部(23°39'54'', 102°35'58'')湖盆分别提取沉积物岩芯(编号分别为YLH-W、YLH-M和YLH-E,长度分别为39、32和28cm).采用1cm间隔现场分样,4℃避光保存运回实验室.
利用沉积物210Pb、137Cs结合CRS(The constant rate of210Pb supply)模型建立异龙湖沉积物钻孔年代序列[51].测年样品的处理过程为:称取5g左右冻干沉积物样品,于标准规格试管中反复压实,测量样品高度,注入约1cm厚度环氧树脂AB结构胶密封静置1个月后将样品放于高纯锗谱仪(BEGe™ GCW3023, Canberra Inc)探测器中,提取沉积物样品210Pbex和137Cs比活度(解谱软件为Genie 2000V3.2.1和LabSOCS™),每个样品提取时间为24h.以上分析内容在高原地理过程与环境变化云南省重点实验室完成.
两件加氢环氧乙烷反应器上、下管板,材质为S A-508 GR.3 CL1,交货尺寸为φ6268mm×310mm(见图1),是制造厂首次生产如此大直径管板锻件。
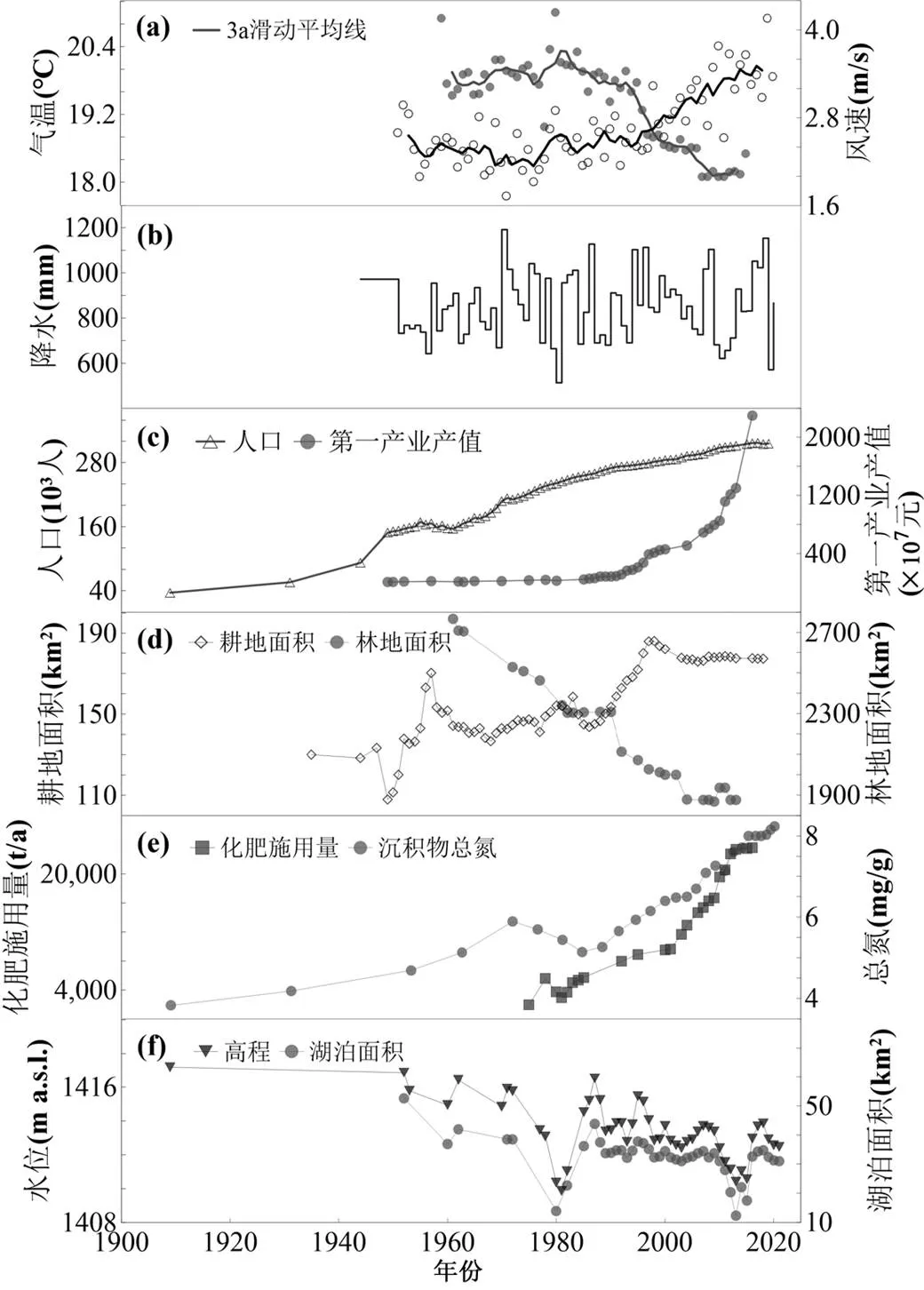
图2 近百年来异龙湖流域环境变化趋势
1.3 样品处理与分析
沉积物磁化率利用英国Bartington MS-2型磁化率仪进行测定.样品处理方法为:将冻干样品放于无磁聚苯乙烯测试盒内反复压实,确保样品填满测试盒后放入仪器中进行测试.利用仪器的不同频段分别测得高频(4650Hz)和低频(465Hz)磁化率,每个样品每个频段反复测量3次,取平均读数,单量程精度为±1%.沉积物粒度的测定方法为:称取约0.5g冻干沉积物样品于烧杯中,加入足量的10%过氧化氢(H2O2),50℃水浴加热至无气泡产生,加入2mol/L盐酸充分反应以去除碳酸盐,后加入足量蒸馏水静置24h并去除上清液.测样前再加入约10mL浓度为0.05mol/L的六偏磷酸钠(Na6O18P6)分散剂,超声振荡10min后用马尔文激光粒度仪(Mastersize 2000,英国)进行粒度测定.
沉积物TP的测定方法采用碱熔-钼锑抗分光光度法(HJ632-2011).样品处理方法如下:称取约0.25g冻干样品于镍坩埚内,加入100μL无水乙醇以及2g氢氧化钠覆盖.在400和640℃下加热15min,将熔融物全部转移至50mL离心管离心并提取上清液.加入2~3滴浓度为0.002g/mL的2,4-二硝基酚作为指示剂,和1mL浓度为0.1g/mL的抗坏血酸溶液并混匀, 30s后加入2mL浓度为0.13g/mL的钼酸盐溶液,再次混匀并静置15min后在700nm波长处测定其吸光值,并根据标准曲线计算TP含量.
采用分光光度法测定沉积物色素.称取约0.4g 样品于聚乙烯离心管并加入25mL 90%丙酮溶液静置萃取10h,用0.45μm聚醚砜针头滤膜过滤萃取液置于100mL容量瓶中.再次向原离心管加入丙酮溶液反复萃取4次,将所有萃取液移至容量瓶,用90%丙酮溶液定容至100mL待测.用紫外分光光度计在666、412、504和528nm波长测定其吸光值,参照Swain(1985)的方法计算叶绿素及其衍生物总量(CD)和蓝藻叶黄素(Myx)浓度[52].
沉积物有机碳含量(TOC)采用酸处理后样品测定,利用FLASH HT 2000元素分析仪联用Thermo Fisher MAT 253同位素质谱仪测定.样品前处理采用Meyers(2001)所描述方法[53],称取约0.5g经研磨过的冻干样品(研磨前去除植物残体、软体动物壳体),用2mol/L盐酸(HCl)在50℃下反应6~8h去除碳酸盐,用去离子水将样品反复冲洗至中性后烘干,称取约8mg样品用锡杯包裹用于测试δ13C和C、N元素含量,分析结果的精度优于±0.1‰,结果报告采用VPDB(Vienna Pee Dee Belemnite)国际标准.全岩样品经冻干磨样处理后直接测定.沉积物无机碳含量(TIC)由全岩样品总碳(TC)和有机碳含量(TOC)相减得出,并根据质量平衡原理计算无机碳同位素(δ13CTIC)[32].以上分析内容均在高原地理过程与环境变化云南省重点实验室完成.
钻孔样品的碳累积速率包括有机碳累积速率(OCAR, g C/m2/a)和无机碳累积速率(ICAR, g C/m2/a),均按标准方法计算获得[9].二者的计算方式为每个间隔分样(1cm)的碳含量(%)乘以每层样品的干物质质量累积速率[SAR,g/(m2×a)]:
SAR=/(×) (1)
OCAR=TOC×SAR (2)
ICAR=TIC×SAR (3)
式中:为每个分样的干物质质量,“g”;为钻管横切面积“m2”;为每层样品的时间间隔“a”,由年代模型确定.
2 结果与分析
2.1 年代模型
钻孔测年的分析结果显示,210Pbex比活度总体上随深度增加而减小(图3(a),(b),(c)),具备利用CRS模型重建年代序列的基础.部分钻孔0~10cm深度210Pbex比活度由下而上逐渐下降,可能与湖泊初级生产力的上升导致210Pbex信号被稀释有关[54].其中,YLH-E钻孔年代学结果也与异龙湖已有研究中相邻钻孔的年代学结果基本一致,28cm深度处对应1930年左右[54].计算结果显示YLH-W、YLH-M和YLH-E 3根钻孔的底部年代分别大致对应1907、1941和1928年.
基于CRS年代模型结果,计算得到了钻孔的沉积速率.结果显示,3根钻孔的干物质累积速率总体随深度的减小而明显增加,且沉积速率变化基本一致,均值在0.4~0.6cm/a范围内.其中,中、东部湖盆的沉积速率增长略快于西部湖盆(图3(d),(e),(f)).
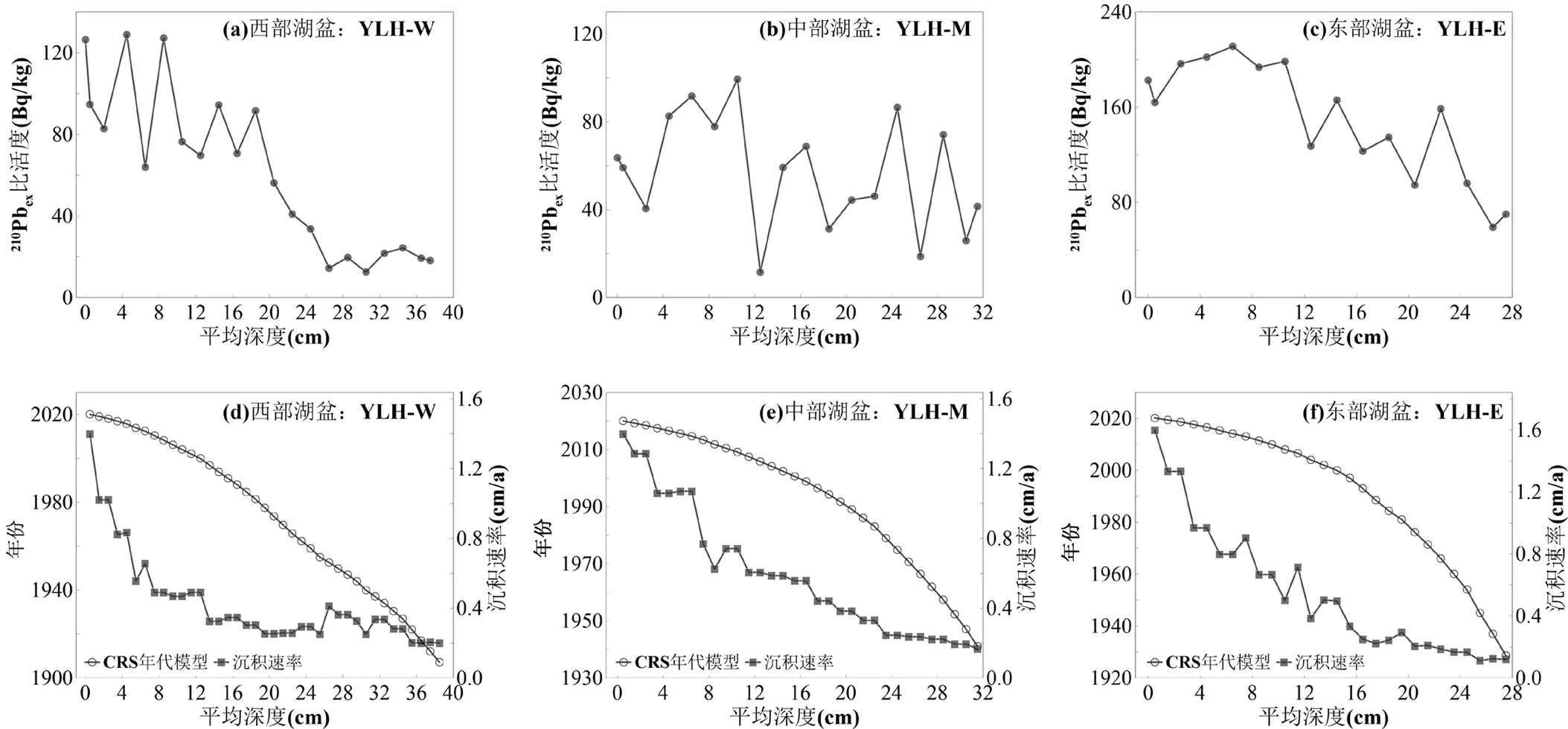
图3 异龙湖钻孔沉积物210Pbex、137Cs比活度曲线以及CRS年代模型结果与沉积速率变化
2.2 沉积物环境代用指标变化
沉积物代用指标的结果表明,近百年来流域地表物质输入总体下降但营养盐输入强度明显增加.异龙湖的粒度测定结果显示,3根钻孔的中值粒径具有快速的波动变化特征,反映了水位升降对浅水湖泊中颗粒物质的搬运过程具有明显的年际差异.但总体上中值粒径呈现减小的趋势(图4b),以1953年开始大规模放水围湖造田为参照,从1953前的(9.56±3.81)μm(=20)减小到1953年以后的(8.87± 3.68)μm(=78)(<0.05).反映了湖泊水位下降背景下水动力的减弱造成粗颗粒物质搬运能力的总体下降(图4(a)).与异龙湖流域耕地面积的扩张以及林地的减少(图2(d))趋势相反,近百年来湖泊沉积物中磁化率信号有所下降.除西部湖盆外,中、东部分湖盆的低频磁化率呈明显的下降趋势(图4(c)).两根钻孔的低频磁化率从1950年前的(78.20± 27.4)´10-8m3/kg(=6)下降至1950年后的(43.50± 10.4)´10-8m3/kg (=51)(<0.05).可能反映了1950年代以来水文调控(如上游筑坝和大量水库的修建)的加强使入湖径流减少从而降低了流域侵蚀物的入湖量.因此,中值粒径、磁化率强度的降低共同反映了近代以来水文调控等导致的流域地表物质输入强度的下降.但另一方面,与入湖径流下降同步,入湖营养盐负荷在明显提升,反映为沉积物中营养盐的浓度呈持续增加的趋势.其中沉积物总氮(TN)由1950年前的(5.99±1.53)mg/g(=17)显著增加到1950年后的(7.59±1.37)mg/g(=81)(< 0.001)(图4d).沉积物总磷(TP)也由1950年前的(0.31±1.03)mg/g(=17)增加到1950年后的(0.51± 1.09)mg/g(=81)(<0.001)(图4(e)).沉积物营养盐的富集与流域农业扩展和化肥施用量的增加具有明显的一致性(图2(e)),总体反映了营养盐输入强度的持续提高.
异龙湖水体逐渐富营养化导致内源初级生产力持续上升.与TN、TP的富集过程同步,沉积物中叶绿素含量(CD)也显示了明显的增长过程,由1900年左右的约0.68u/g(org)增加到2020年左右的约1.36u/g(org)(图4 (f)).蓝藻叶黄素也由1900年代的7.02μg/g(org)显著增加到2020年左右的约38.99μg/g(org)(图4(g)).并且蓝藻叶黄素解释了沉积物叶绿素主要的变化(2=0.91,<0.001),进一步表明了营养盐富集造成的湖泊富营养化导致了藻类初级生产力的明显上升,而藻类生物量上升则是异龙湖内源初级生产力增加的主要表现.同时,藻类生物量的持续增加逐渐主导了异龙湖的碳埋藏过程,表现为沉积物有机碳氮比(C:N)值的持续减小以及沉积物δ13CTIC和δ13CTOC的同步下降.C:N值从1950年前的(11.90±1.58)(=17)显著减小到1950年后的(8.88±1.92) (=81)(<0.001)(图4 (h)).沉积物δ13CTIC则从1950年前的(-6.11±5.87)‰(=17)下降到1950年后的(-8.52±6.20)‰(=81)(<0.05)(图4 (m)).沉积物δ13CTOC也从1950年之前的(-23.96±0.98)‰ (=17)下降至1950年以后的(-26.67±0.92)‰ (<0.001) (=81) (图4 (n)).反映了流域地表物质输入减少、湖泊内源初级生产力上升背景下藻源有机质和自生碳酸盐积累的增加.
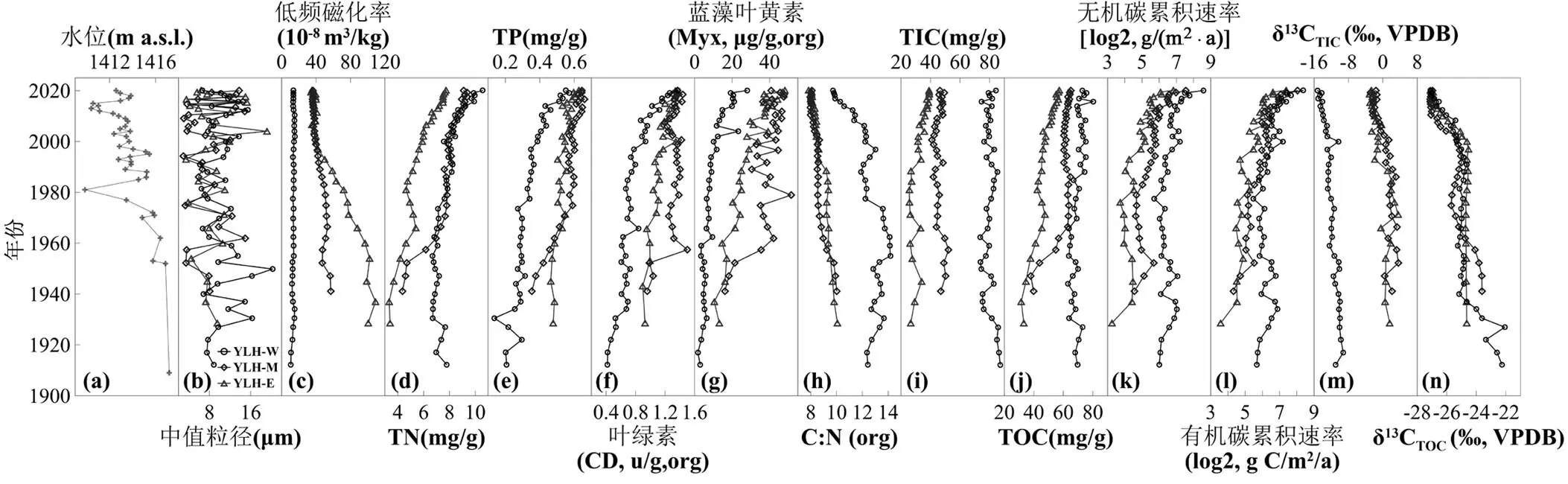
图4 异龙湖沉积物环境代用指标的变化趋势
(a)水位; (b)中值粒径; (c)低频磁化率; (d)总氮; (e)总磷; (f)叶绿素; (g)蓝藻叶黄素; (h)有机碳氮比; (i)总无机碳; (j)总有机碳; (k)无机碳累积速率; (l)有机碳累积速率; (m)无机碳同位素; (n)有机碳同位素
2.3 沉积物碳含量及碳埋藏通量变化
与藻类生物量的持续上升相对应,近百年来异龙湖沉积物碳含量出现明显上升趋势.沉积物总碳含量(TC)由1900年左右的约102.66mg/g增加到2020年左右的约118.31mg/g.其中沉积物有机碳含量(TOC)呈明显的增加趋势,由1950年前的(56.90±15.70)mg/g(=17)增加到1950年后的(61.07± 9.58)mg/g (=81)(<0.05),TOC含量的空间分布表现为西部湖盆>中部湖盆>东部湖盆(图4 (j)).另一方面,近百年来西、中部湖盆沉积物无机碳含量(TIC)呈现小幅下降的趋势,从由1950年前的(66.30± 21.70)mg/g(=17)下降至1950年后的(53.10± 19.90)mg/g(=81)(<0.05).但东部湖盆TIC含量的变化趋势与TOC的变化趋势总体一致(图4 (i)),显示了明显的增加趋势.
近百年来沉积物碳累积速率也在持续增加.3根钻孔的分析结果显示了近百年来异龙湖有机碳累积速率(OCAR)呈整体上升趋势.OCAR从1950年前的(38.40±29.50)g C/m2/a(=17)显著增加到1950年后的(84.90±50.00)g C/m2/a (=81)(<0.001)(图4 (l)).而无机碳累积速率(ICAR)也显示了与有机碳高度同步的增长趋势,从1950年前的(44.90±34.80)g C/(m2×a) (=17)显著增加到1950年后的(72.70± 49.90)g C/(m2×a) (=81)(<0.01)(图4 (k)).显示了在部分湖盆TIC含量下降的背景下,由于沉积速率的上升(图3),ICAR仍然保持了稳定的增长.空间上,西部湖盆ICAR和OCAR总体高于中、东部湖盆.总体而言,无机碳累积速率与有机碳累积速率保持了高度的同步性,二者的相关系数在YLH-W、YLH-M和YLH-E 3根钻孔中达到0.98~0.99(<0.001),显示了无机碳埋藏和有机碳埋藏在响应环境压力的过程中高度的协同变化特征.以上结果表明近百年来异龙湖的碳埋藏通量出现了显著的增长,并表现为有机碳累积速率与无机碳累积速率保持协同增加的趋势.
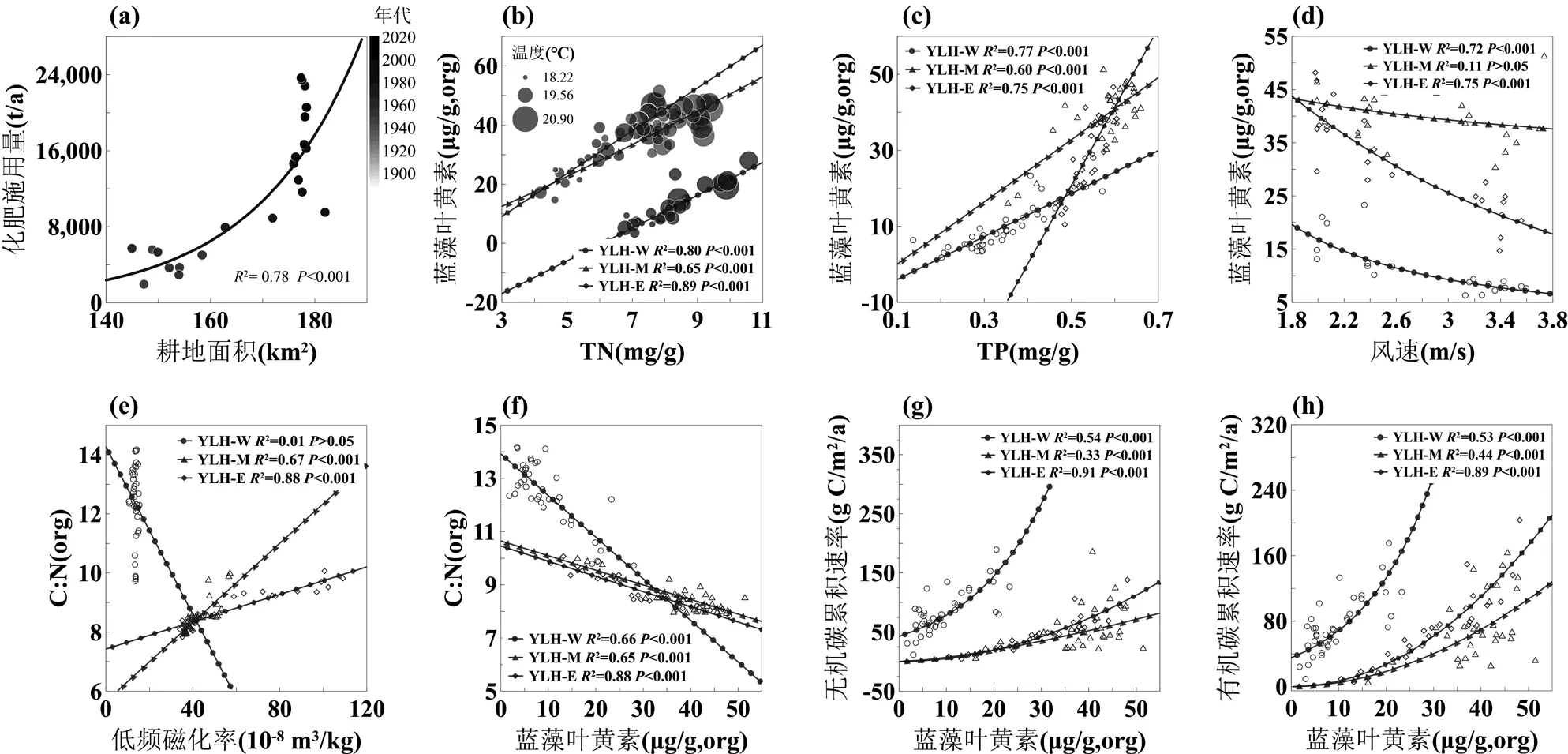
图5 异龙湖无机碳和有机碳埋藏过程中主要环境代用指标之间的关系
(a)化肥施用量与耕地面积的关系; (b)蓝藻叶黄素浓度与总氮的关系; (c)蓝藻叶黄素与总磷的关系; (d)蓝藻叶黄素与风速的关系; (e)有机碳氮比与低频磁化率的关系; (f)有机碳氮比与蓝藻叶黄素的关系; (g)无机碳累积速率与蓝藻叶黄素的关系; (h)有机碳累积速率与蓝藻叶黄素的关系
3 讨论
3.1 流域开发和气候变化背景下湖泊碳埋藏通量的变化
近百年来异龙湖流域开发和区域气候变化共同促进了湖泊富营养化和藻类生物量的增加.自1950年以来,流域人口的增长带动了耕地面积的不断扩张(图2(c),(d)).耕地扩张又引起化肥施用量的上升(图5(a)),进一步导致了水体和湖泊沉积物中总氮和总磷的持续积累(图4(d),(e)).营养盐的富集使水体逐渐富营养化,并促进了藻类生物的增加.早在1957年,铜绿微囊藻已经在异龙湖西部湖区形成了局部水华[33].这反映了蓝藻作为嗜营养藻类,其生物量的增加对流域营养盐输入具有敏感的响应特征[58-59].由于流域营养盐负荷的持续增加,至2009年异龙湖已转变为蓝藻占主导的藻型湖泊[47,49-50].水体的逐步富营养化和蓝藻的占优,与沉积物蓝藻叶黄素含量的持续上升是同步的(图4(g)).已有研究表明,氮、磷等营养盐供给是藻类爆发的主要驱动因素[60-63].而在异龙湖中,TN分别解释了西、中、东湖盆沉积物中80%、65%和89%的蓝藻叶黄素浓度变化(<0.001)(图5(b);表2),而总磷的解释量也分别达到了77%、60%和75%(<0.001)(图5(c)).这些结果显示了近百年来流域开发导致的营养盐输入是异龙湖水体富营养化和藻类爆发的重要驱动因素.
此外,区域增温和风速下降的持续客观上有利于藻类生长和初级生产力提升.温度分别解释了西、中、东湖盆蓝藻叶黄素变化的42%、14%和55%(图5(b)),风速则分别解释了72%、11%和77%(图5(d)).近百年来蓝藻生物量对区域变暖的正向响应,主要是由其喜温性决定的.蓝藻的生长曲线具有明显的温度响应特征[63-64].同时,蓝藻DNA和蛋白质合成系统的热稳定性帮助其在温度升高的环境中形成竞争优势[65].增温背景下,近百年来异龙湖蓝藻的增长和占优与区域及全球湖泊研究结果是一致的[66-68].而风速的下降则有利于减少水体垂直混合,从而促进蓝藻水华的发展、扩散和维持[69].综上所述,近百年来异龙湖流域开发和气候变化等因素共同导致了异龙湖水体的富营养化和藻类生物量的上升.
钻孔记录与对比分析表明,藻类生物量的增加显著提高了近百年来异龙湖的有机碳埋藏通量.已有研究表明,水体富营养化和藻类生物量增加是近代以来全球湖泊有机碳埋藏速率快速增长的主要原因[5,10,70-71].在异龙湖中,藻类生物量和营养盐是近百年来有机碳累积速率增加的主要驱动因子(表2),蓝藻叶黄素的增加解释了主要的有机碳埋藏通量的增长(图5(h)),显示了与已有研究相似的模式.借助C:N可以有效识别有机碳埋藏的主要来源.异龙湖流域陆生植被的C:N>20(均值(54.03±40.05),=20),水生维管束植物>15[72],而藻类约为6~8[72].因此,藻类生物量的增长将导致沉积物中内源有机碳的积累和C:N值下降(图5(f)),表现为C:N从1900年左右的~11.60下降到2020年左右的~8.55(图4(h)),表明了藻源有机质积累是异龙湖有机碳埋藏增加的主要来源.而有机C:N与低频磁化率的关系同样也反映了内源贡献的增长是异龙湖有机碳埋藏上升的主要原因(图5(e)).此外,异龙湖浮游藻类的碳稳定同位素值约为-27‰[73],明显低于大型水生植物(-10‰~-17‰)[73]和陆源有机质(约-25.1‰,=19),因此3个湖盆中沉积物δ13CTOC的显著下降(图4 (n))也同样反映了藻类生物量的上升对有机碳埋藏过程的主导作用.这些研究结果共同表明水体富营养化背景下,藻类生物量的增加导致了湖泊内源有机碳的积累并提高了异龙湖的有机碳埋藏速率.
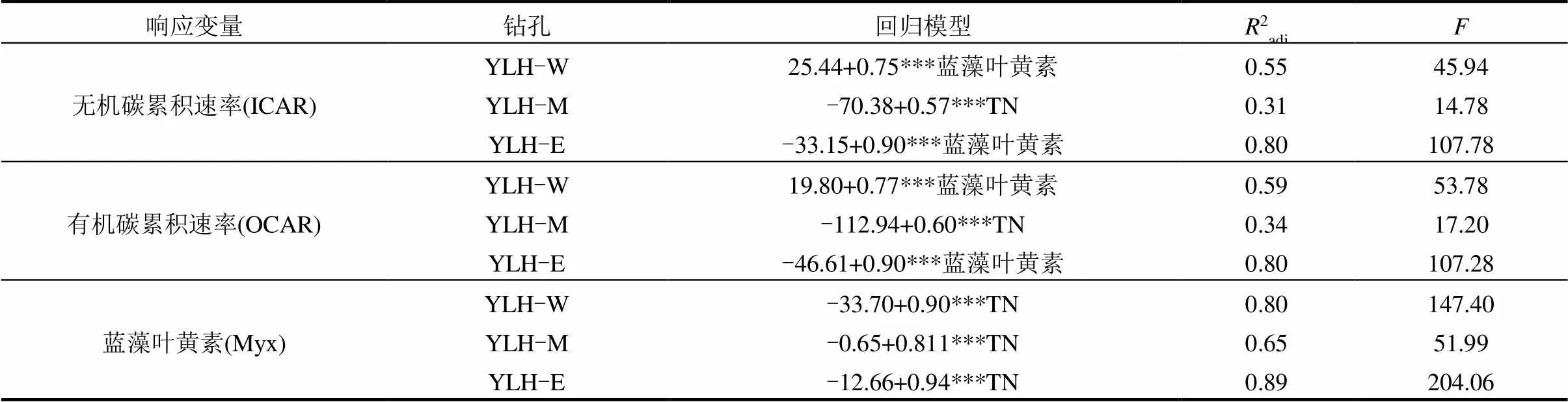
表2 异龙湖碳埋藏速率和蓝藻叶黄素变化的多元回归模型
注:***代表<0.001的显著性水平.
藻类生物的上升同样也促进了异龙湖无机碳埋藏通量的持续增加.已有研究表明,蓝藻等初级生产者的光合作用是湖泊水体中碳酸盐沉淀发生的主要诱因之一[74-78].藻类光合作用通过消耗水体CO2和释放OH-提升水体pH值,来提高矿物饱和度.蓝藻等为应对CO2不足而进化的碳浓缩机制(CCMs),也为其诱导碳酸盐沉淀提供了有利条件[77,79].同时,蓝藻等微型藻类的生长还为碳酸盐的结晶提供了凝结核,进一步促进了碳酸盐的沉淀[78],并使部分水体在欠饱和的情形下也能形成碳酸盐沉积.而藻类爆发可引起大量碳酸盐结晶沉淀并导致了部分湖泊出现“白化”现象[80].异龙湖中,水体的富营养化造成藻类的大量繁殖和原生碳酸盐矿物的大量沉积(图7 (a), (b)),无机碳埋藏速率的变化受到受蓝藻生物量的显著驱动(图5 (g);表2),反映了水体富营养化过程中藻类生物量的上升明显提高了自生碳酸盐的沉积和埋藏速率.另一方面,异龙湖中光叶眼子菜、轮藻、狐尾藻等沉水植物对CO2的光合吸收也可引起碳酸盐沉淀[81].在转变为藻型湖泊之前,水生植物的光合作用可能是异龙湖无机碳沉积的重要来源.然而,异龙湖近百年来沉水植物不断减少并趋于消亡,浮游藻类逐步增加并占优[47,49,54].在此过程中,沉水植物的减少与无机碳累积速率的增加趋势是相反的,证明大型水生植物对近百年来异龙湖无机碳埋藏的影响相对次要.除此之外,温度是控制湖泊无机碳沉积的重要因素之一.在增温背景下,湖水蒸发浓缩以及水体CO2溶解度下降等过程有利于碳酸盐析出[24,75,82].然而,温度对异龙湖西、中、东3根钻孔无机碳累积速率的解释程度分别只有20%、27%和52% (2=0.20、0.27和0.52;<0.05、0.01和0.001).在多元回归模型中,温度也不是无机碳累积速率变化的主要驱动因子(表2),表明了近百年来生物因素对异龙湖无机碳埋藏的驱动强度大于物理化学因素.进一步地,相关分析结果显示西、中、东湖盆无机碳累积速率与低频磁化率的相关系数分别为0.05、-0.83和-0.92(>0.05、<0.001和0.001),表明流域输入对异龙湖无机碳埋藏影响也较小,无机碳埋藏过程主要受湖泊内循环过程控制.这与磁化率、粒度结果显示的近百年来异龙湖流域物质输入强度总体下降的趋势是一致的.流域植被破坏、大量蓄水设施的修建以及人为取水的快速增加使1950年以来入湖径流量和地表颗粒物质入湖量的逐渐下降,这可能是异龙湖中值粒径减小和主要湖盆磁化率下降的重要原因,总体反映了流域碳酸盐输入相对次要.这些结果共同反映了在水体逐步富营养化过程中,藻类生物量的增加是异龙湖无机碳埋藏加速的主要原因之一.
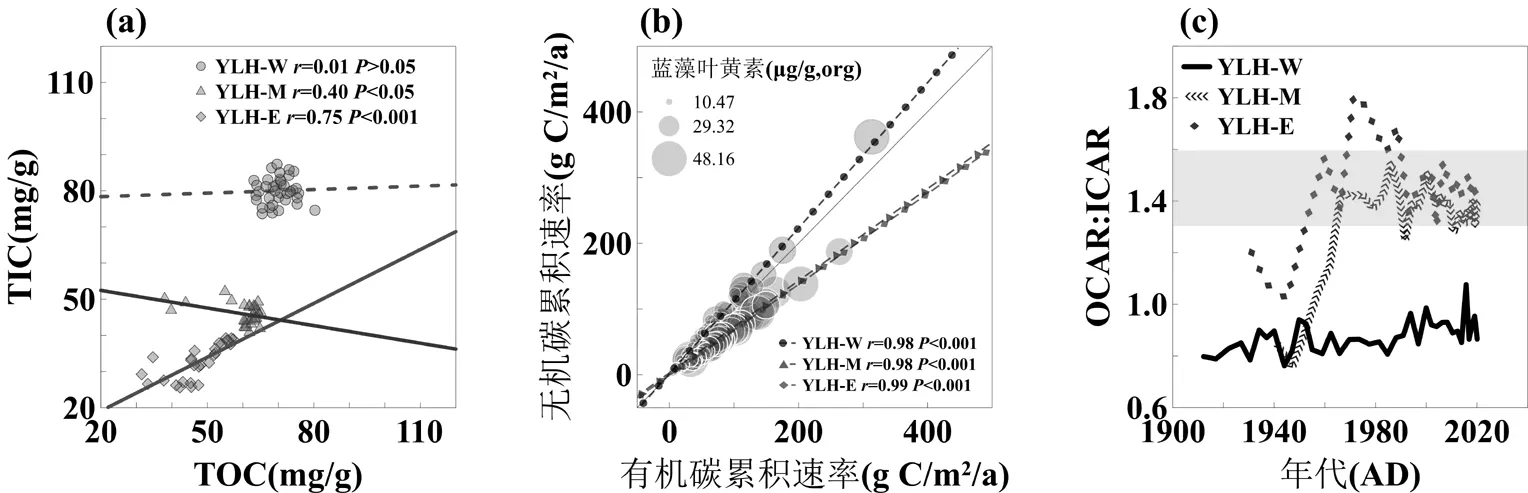
图6 异龙湖沉积物总无机碳(TIC)与总有机碳(TOC)之间的协同变化关系
灰色区域代表光合作用固定有机碳和自生碳酸盐的理论比值区间(a)沉积物TIC与TOC的关系; (b)无机碳累积速率与有机碳累积速率的关系; (c)OCAR:ICAR的时间变化
3.2 水体富营养化过程中有机碳和无机碳埋藏的协同变化特征
近百年来异龙湖有机碳和无机碳埋藏速率呈现明显的共变特征,并主要受湖泊初级生产力驱动.已有研究表明,岩溶地区湖泊等水体的水生光合作用驱动了DIC向内源有机质和自生碳酸盐的同步转化,并通过“生物碳泵”效应实现内源有机碳和无机碳的同步沉积[8,15,19,22,83-87].而在水体富营养化过程中,内源初级生产力的增长则可以同步提高有机碳和无机碳的埋藏通量[88].异龙湖近百年来有机碳累积速率和无机碳积累速率呈现极其显著的共变关系,并且同时与藻类生物量呈明显的共变特征(=0.98~0.99,<0.001,图6(b);图5(g), (h)),同时与沉积物有机C:N值的下降一致(图4(h)),表明了藻类初级生产力是驱动有机碳和无机碳埋藏同步变化的主要因素.异龙湖沉积物δ13CTIC和δ13CTOC的高度共变特征(西、中、东3根钻孔2分别为0.67、0.72和0.58,<0.001)也反映了光合作用和“生物碳泵”效应对碳埋藏过程的驱动作用[20].藻类光合作用将水体DIC转化为内源有机质的同时,促进了水体碳酸盐矿物饱和度的上升以及自生碳酸盐的形成,从而使有机碳与无机碳沉积同步发生.通常,水生光合固碳及其碳酸盐沉淀固碳的物质的量比在1.3~1.6之间[89-91].在不考虑有机碳矿化以及碳酸盐溶蚀的情况下,由生物因素主导的碳埋藏过程将导致有机碳和无机碳埋藏通量随湖泊初级生产力发生同步变化[28,88].这些结果共同反映了湖泊初级生产力是控制近百年来异龙湖有机碳和无机碳埋藏通量同步变化的主要因素,而藻类光合作用以及“生物碳泵”效应是异龙湖碳埋藏的主要驱动过程.
受埋藏过程以及有机碳-无机碳的相互作用影响,沉积物有机碳和无机碳含量的共变关系与埋藏通量明显不同.异龙湖东部湖盆中,碳埋藏过程主要受“生物碳泵”主导,沉积物TIC和TOC含量呈显著的共变特征(图6 (a),YLH-E钻孔),与云南其他湖泊的研究结果较为类似[23-24].而另一方面,浅水湖区水体富营养化形成的大量有机质在氧化过程中产生的H+将导致碳酸盐的溶蚀[29,31](图7 (d)),加上内源贡献或外源输入造成的有机碳大量积累也可能对沉积物TIC的含量形成稀释效应[24],因此可能导致了异龙湖中部湖盆TIC和TOC的反向变化趋势(图6 (a),YLH-M钻孔).此外,微生物的参与使部分有机碳在矿化过程中被转化为湖泊自生碳酸盐[92],从而造成TIC和TOC含量此消彼长或共同增加的趋势.在以上多种因素共同作用下,可能使沉积物TIC与TOC含量变化没有明显相关性(图6 (a),西部湖盆YLH-W钻孔).这也与滇中地区富营养化浅水湖泊的研究结果是一致的[27].由此可见,在同一湖泊中,碳埋藏过程的差异以及有机碳-无机碳的相互作用可以使二者含量之间呈现不同的共变属性,这与水体富营养化的背景下,“生物碳泵”效应造成的有机碳-无机碳的埋藏通量同步上升的趋势有明显区别.因此对于气候变化以及水体富营养化压力下湖泊碳埋藏过程而言,有机碳-无机碳含量和通量的变化可能具有不同的环境指示意义.
有机碳-无机碳的转化过程可能促进其埋藏通量的协同变化.受湖泊初级生产力上升的驱动,近百年来沉积物OCAR: ICAR整体增长并趋于接近“生物碳泵”效应1.3~1.6的理论比值范围(图6(d)).表明水体富营养化过程中,沉积物有机碳-无机碳埋藏的协同增长呈现有机碳快于无机碳的总体特征.然而在初级生产力较高的西部湖盆(图4(j)),沉积物OCAR: ICAR远低于1.3~1.6比值范围(图6(d)灰色区域).可能的原因主要包括:一是浅水湖区(西部湖盆平均水深只有~2m,图1)有机质的充分曝氧使有机碳不易积累[93-94];二是内源有机碳大量转化为自生碳酸盐[32,95],从而导致了OCAR:ICAR的值较低.已有研究显示,富营养化过程中水体δ13CDIC有逐渐正偏移的趋势[15,75,88,96],因此表层水体沉积的原生碳酸盐的δ13C值应当相应地增大.而在异龙湖,随着藻类生物量的增加(C:N比值的下降),3个湖盆沉积物δ13CTIC值都出现了明显的减小过程(图4(m)),表明有机碳转化形成的自生碳酸盐的占比可能在增加(即成岩碳酸盐[82,97],其δ13C与有机碳接近).尤其是西部湖盆的δ13CTIC值远低于中、东部湖盆(图4(m)),可能反映了其更高的成岩碳酸盐占比.已有研究表明,在有机质矿化过程中,包括氨化、反硝化、硫酸盐/铁/锰还原、产甲烷等过程均可因微生物活动而导致微环境中pH值和碱度的上升[92,98-100],进而诱发成岩碳酸盐的形成和有机碳向无机碳的转化.因此上述的第二种原因即内源有机碳向自生碳酸盐的转化可能是西部湖盆OCAR:ICAR低于中、东部湖盆的重要原因.这也表明在初级生产力较高的湖区,有机碳通过早期成岩向无机碳的转化可能更为明显.另一方面,较高的有机质供给和氧化(色素的保存度较低:图4(f),(g))可能造成了西部湖盆较为明显的碳酸盐溶蚀(图7(c)),理论上将阻碍有机碳-无机碳埋藏通量的同步增加.然而,有机碳向无机碳埋藏的转化可以较好地补偿碳酸盐溶蚀造成的无机碳损失,从而维持了较高的无机碳累积速率并使有机碳和无机碳保持同步累积(图4(k), (l);图6 (b)).因此,在西部湖盆中,通过有机碳-无机碳转化以及有机质氧化等作用,可以使无机碳累积速率总体高于有机碳累积速率并保持了二者的协同增加(图6(b)).由此可见,在异龙湖富营养化过程中,有机碳-无机碳的转化可能是形成二者埋藏通量高度协同变化的重要原因之一.
3.3 有机碳和无机碳埋藏的协同变化对岩溶区湖泊碳埋藏评估的指示意义
异龙湖研究结果表明的有机碳-无机碳的协同变化过程为湖泊碳埋藏评估提供了新的思路,尤其是有机碳与无机碳之间的转化路径表明了基于有机碳埋藏通量的评估模式可能已经低估了湖泊碳埋藏的动态变化过程.在非岩溶区湖泊中,有机碳是湖泊碳埋藏的主要形式,但是在岩溶区湖泊等常见的硬水水体中,无机碳是同等重要的碳埋藏组分[3].硬水水体占全球内陆水体体积的50%左右[101],硬水水体接收了流域岩石风化和大气沉降带来的大量无机碳,其埋藏通量可以达到Pg C/a级别[18],是不可忽视的陆地碳汇过程.然而在已有的研究中,碳埋藏的评估模式主要基于有机碳,可能已经低估了内陆水体的碳埋藏通量[3].尤其是在岩溶地区,湖泊等水体因为碳酸盐风化耦合水生光合作用而形成了持续的有机碳和无机碳埋藏,因此被认为是重要的碳汇过程[8,19,22,83-85].异龙湖研究的结果表明,内源有机碳通过矿化作用部分转化为自生碳酸盐,既可以补偿富营养化过程中有机质积累导致的碳酸盐溶蚀和损失,又促进了有机碳和无机碳埋藏通量的协同增长.更为重要的是,通过内源有机碳向自生碳酸盐的转化,实现了无机形式的有机碳埋藏并促进了湖泊碳埋速率的上升.在水体富营养化过程中,随着藻类生物量的增加,内源有机质向自生碳酸盐的转化明显加快,对湖泊碳埋藏通量的促进作用进一步提升.这些结果显示了有机碳-无机碳之间的转化过程及其协同作用可以显著改变岩溶地区湖泊的碳埋藏速率,而这些过程在已有的研究中很少被关注.因此仅基于有机碳的碳埋藏评估模式,可能低估了湖泊碳埋藏的动态变化过程进而影响了湖泊碳汇评估的准确性.异龙湖研究结果所显示的有机碳-无机碳的转化路径及其协同变化过程为岩溶区湖泊碳埋藏和碳汇的评估提供了新的视角.在人类活动与气候变化持续的背景下,对岩溶地区湖泊碳埋藏模式的科学评估需要关注无机碳埋藏过程以及有机碳-无机碳的协同作用.

图7 异龙湖沉积物典型碳酸盐矿物的扫描电镜图像
(a)原生碳酸盐; (b)藻类分解后留下的规则孔洞; (c)西部湖盆碳酸盐溶蚀痕迹; (d)中部湖盆碳酸盐溶蚀痕迹
4 结论
4.1 近百年来流域开发和气候变化共同导致了湖泊初级生产力的明显提高,表现为沉积物色素含量的持续增加(如蓝藻叶黄素由~7.02μg/g上升至~ 38.99μg/g).
4.2 受藻类生物量上升的驱动,有机碳和无机碳埋藏速率呈明显的同步增长趋势(=0.98~0.99,<0.001),并表现为ICAR、OCAR和蓝藻叶黄素的高度共变性.
4.3 近百年来异龙湖有机碳-无机碳埋藏显示了高度协同变化的长期模式,反映了水体富营养化背景下,藻类光合作用以及“生物碳泵”效应对碳埋藏过程的显著驱动作用.
4.4 不同湖区碳埋藏过程的差异以及有机碳-无机碳的相互作用,使沉积物有机碳和无机碳含量出现了不同的协同变化特征.并且同位素证据显示了藻类生物量的上升明显增加了内源有机碳向自生碳酸盐的转化,进而促进了有机碳-无机碳埋藏通量的协同增长以及湖泊碳埋藏速率的显著提高.
这些结果表明对岩溶地区湖泊碳埋藏模式和湖泊碳汇的科学评估,需要重视无机碳埋藏过程以及有机碳-无机碳协同作用的影响.
[1] Cole J J, Prairie Y T, Caraco N F, et al. Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget [J]. Ecosystems, 2007,10(1):172-185.
[2] Tranvik L J, Downing J A, Cotner J B, et al. Lakes and reservoirs as regulators of carbon cycling and climate [J]. Limnology and Oceanography, 2009,54(6part2):2298-2314.
[3] Drake T W, Raymond P A, Spencer R G M. Terrestrial carbon inputs to inland waters: A current synthesis of estimates and uncertainty [J]. Limnology and Oceanography Letters, 2018,3(3):132-142.
[4] Battin T J, Luyssaert S, Kaplan L A, et al. The boundless carbon cycle [J]. Nature Geoscience, 2009,2(9):598-600.
[5] Mendonça R, Müller R A, Clow D, et al. Organic carbon burial in global lakes and reservoirs [J]. Nature communications, 2017,8(1):1694.
[6] Dean W E, Gorham E. Magnitude and significance of carbon burial in lakes, reservoirs, and peatlands [J]. Geology, 1998,26(6):535-538.
[7] Einsele G, Yan J, Hinderer M. Atmospheric carbon burial in modern lake basins and its significance for the global carbon budget [J]. Global and Planetary Change, 2001,30(3/4):167-195.
[8] Liu Z, Macpherson G L, Groves C, et al. Large and active CO2uptake by coupled carbonate weathering [J]. Earth-Science Reviews, 2018, 182:42-49.
[9] Heathcote A J, Anderson N J, Prairie Y T, et al. Large increases in carbon burial in northern lakes during the Anthropocene [J]. Nature Communications, 2015,6(1):1-6.
[10] Anderson N J, Heathcote A J, Engstrom D R, et al. Anthropogenic alteration of nutrient supply increases the global freshwater carbon sink [J]. Science Advances, 2020,6(16):eaaw2145.
[11] Gui Z, Xue B, Yao S, et al. Organic carbon burial in lake sediments in the middle and lower reaches of the Yangtze River Basin, China [J]. Hydrobiologia, 2013,710(1):143-156.
[12] Zhang F, Yao S, Xue B, et al. Organic carbon burial in Chinese lakes over the past 150years [J]. Quaternary International, 2017,438:94-103.
[13] Balmer M B, Downing J A. Carbon dioxide concentrations in eutrophic lakes: undersaturation implies atmospheric uptake [J]. Inland Waters, 2011,1(2):125-132.
[14] Pacheco F S, Roland F, Downing J A. Eutrophication reverses whole-lake carbon budgets [J]. Inland waters, 2014,4(1):41-48.
[15] Nõges P, Cremona F, Laas A, et al. Role of a productive lake in carbon sequestration within a calcareous catchment [J]. Science of the Total Environment, 2016,550:225-230.
[16] Grasset C, Sobek S, Scharnweber K, et al. The CO2-equivalent balance of freshwater ecosystems is non‐linearly related to productivity [J]. Global Change Biology, 2020,26(10):5705-5715.
[17] Yu Z, Wang X, Zhao C, et al. Carbon burial in Bosten Lake over the past century: Impacts of climate change and human activity [J]. Chemical Geology, 2015,419:132-141.
[18] Li Y, Zhang C, Wang N, et al. Substantial inorganic carbon sink in closed drainage basins globally [J]. Nature Geoscience, 2017,10(7): 501-506.
[19] He H, Liu Z, Chen C, et al. The sensitivity of the carbon sink by coupled carbonate weathering to climate and land-use changes: Sediment records of the biological carbon pump effect in Fuxian Lake, Yunnan, China, during the past century [J]. Science of the Total Environment, 2020,720:137539.
[20] 张风菊,薛 滨,姚书春.1850年以来呼伦湖沉积物无机碳埋藏时空变化[J]. 湖泊科学, 2019,31(6):1770-1782. Zhang F J, Xue B, Yao S C. Spatiotemporal pattern of inorganic carbon sequestration in Lake Hulun since 1850 [J]. J. Lake Sci., 2019, 31(6):1770-1782.
[21] Goldscheider N, Chen Z, Auler A S, et al. Global distribution of carbonate rocks and karst water resources [J]. Hydrogeology Journal, 2020,28(5):1661-1677.
[22] Liu Z, Dreybrodt W, Wang H. A new direction in effective accounting for the atmospheric CO2budget: considering the combined action of carbonate dissolution, the global water cycle and photosynthetic uptake of DIC by aquatic organisms [J]. Earth-Science Reviews, 2010, 99(3/4):162-172.
[23] Hodell D A, Brenner M, Kanfoush S L, et al. Paleoclimate of southwestern China for the past 50,000yr inferred from lake sediment records [J]. Quaternary Research, 1999,52(3):369-380.
[24] 陈敬安,万国江,汪福顺,等.湖泊现代沉积物碳环境记录研究[J]. 中国科学(D辑:地球科学), 2002,32(1):73-80. Chen J A, Wan G J, Wang F S, et al. A study of the environmental record of carbon in modern lacustrine sediments [J]. Science China Earth Sciences, 2002,32(1):73-80.
[25] Hillman A L, Abbott M B, Yu J Q. Climate and anthropogenic controls on the carbon cycle of Xingyun Lake, China [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018,501:70-81.
[26] Liu Y, Chen G, Meyer-Jacob C, et al. Land-use and climate controls on aquatic carbon cycling and phototrophs in karst lakes of southwest China [J]. Science of The Total Environment, 2021,751:141738.
[27] 王 露,陈光杰,黄林培,等.近百年来湖泊有机碳与无机碳埋藏响应流域开发的协同变化——以石林喀斯特地区为例[J]. 湖泊科学, 2022,34(5):1751-1764. Wang L, Chen G J, Huang L P, et al. Synchronic changes of lake organic and inorganic carbon burial in response to catchment development over the past century: A case study of Shilin Karst area [J]. J. Lake Sci, 2022,34(5):1751-1764.
[28] Huang C, Zhang L, Li Y, et al. Carbon and nitrogen burial in a plateau lake during eutrophication and phytoplankton blooms [J]. Science of the Total Environment, 2018,616:296-304.
[29] Dean W E. The carbon cycle and biogeochemical dynamics in lake sediments [J]. Journal of paleolimnology, 1999,21(4):375-393.
[30] Wu J L, Gagan M K, Jiang X H, et al. Sedimentary geochemical evidence for recent eutrophication of Lake Chenghai, Yunnan, China [J]. Journal of Paleolimnology, 2004,32(1):85-94.
[31] Müller B, Wang Y, Dittrich M, et al. Influence of organic carbon decomposition on calcite dissolution in surficial sediments of a freshwater lake [J]. Water Research, 2003,37(18):4524-4532.
[32] Sun D, He Y, Wu J, et al. Hydrological and ecological controls on autochthonous carbonate deposition in lake systems: A case study from Lake Wuliangsu and the global perspective [J]. Geophysical Research Letters, 2019,46(12):6583-6593.
[33] 黎尚豪,俞敏娟,李光正,等.云南高原湖泊调查 [J]. 海洋与湖沼, 1963,5(2):87-113.Li S H, Yu M K, Li K C, et al. Limnological survey of the lake of Yunnan plateau [J]. Oceanologia et Limnologia Sinica, 1963,5(2):87-113.
[34] 王云飞,胡文英,张秀珠.云南湖泊的碳酸盐沉积[J]. 海洋与湖沼, 1989,20(2):122-130,197-198. Wang Y F, Hu W Y, Zhang X Z. Carbonate sediments in lakes of Yunnan, China [J]. Oceanologia et Limnologia Sinica, 1989,20(2): 122-130,197-198.
[35] 王丽芳,熊永强,吴丰昌,等.滇池的富营养化过程:来自结合态脂肪酸C(16:0)δ13C的证据[J]. 湖泊科学, 2009,21(4):456-46. Wang L F, Xiong Y Q, Wu F C, et al. The eutrophication process of Lake Dianchi: Evidences from the δ13C value of the bound C(16:0)fatty acid [J]. J. Lake Sci, 2009,21(4):456-464.
[36] Zhou Q, Wang W, Huang L, et al. Spatial and temporal variability in water transparency in Yunnan Plateau lakes, China [J]. Aquatic Sciences, 2019,81(2):1-14.
[37] Liu W, Li S, Bu H, et al. Eutrophication in the Yunnan Plateau lakes: The influence of lake morphology, watershed land use, and socioeconomic factors [J]. Environmental Science and Pollution Research, 2012,19(3):858-870.
[38] Huang C, Yao L, Zhang Y, et al. Spatial and temporal variation in autochthonous and allochthonous contributors to increased organic carbon and nitrogen burial in a plateau lake [J]. Science of the Total Environment, 2017,603:390-400.
[39] 刘会基,刘恩峰,于真真,等.近百年来洱海沉积物有机碳埋藏时空变化[J]. 湖泊科学, 2019,31(1):282-292. Liu H J, Liu E F, Yu Z Z, et al. Spatio-temporal patterns of organic carbon burial in the sediment of Lake Erhai in China during the past 100years [J]. J. Lake Sci, 2019,31(1):282-292.
[40] Liu W, Wu J, Zeng H, et al. Geochemical evidence of human impacts on deep Lake Fuxian, southwest China [J]. Limnologica, 2014,45:1-6.
[41] Wu D, Zhou A, Chen X, et al. Hydrological and ecosystem response to abrupt changes in the Indian monsoon during the last glacial, as recorded by sediments from Xingyun Lake, Yunnan, China [J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2015,421:15-23.
[42] Zan F, Huo S, Xi B, et al. A 60-year sedimentary record of natural and anthropogenic impacts on Lake Chenghai, China [J]. Journal of Environmental Sciences, 2012,24(4):602-609.
[43] 吴 汉,常凤琴,张虎才,等.异龙湖近百年来湖泊沉积物有机碳、氮稳定同位素变化及其环境指示意义[J]. 生态学杂志, 2020,39(8):2478-2487. Wu H, Chang F Q, Zhang H C, et al. Changes of organic C and N stable isotope and their environmental implication during the past 100 years of Lake Yilong [J]. Chinese Journal of Ecology, 2020,39(8): 2478-2487.
[44] Li Y, Liu Y, Zhao L, et al. Exploring change of internal nutrients cycling in a shallow lake: a dynamic nutrient driven phytoplankton model [J]. Ecological Modelling, 2015,313:137-148.
[45] 杨 岚,李 恒.云南湿地[M]. 北京:中国林业出版社, 2010:94-101. Yang L, Li H. Wetlands of Yunnan [M]. Beijing: China Forestry Publishing House, 2010:94-101.
[46] 王苏民,窦鸿身.中国湖泊志[M]. 北京:科学出版社, 1998:368-388. Wang S M, Dou H S. Lakes annals of China [M]. Beijing: Science Press, 1998:368-388.
[47] 崔保山,翟红娟.高原湿地功能退化的表征及其恢复中的生态需水量[J]. 科学通报, 2006,51(S2):106-113. Cui B S, Zhai H J. Characterization of functional degradation of plateau wetlands and their ecological water demand in restoration [J]. Chinese Science Bulletin, 2006,51(S2):106-113.
[48] 魏 翔,唐光明.异龙湖近20年来营养盐与水生生态系统变化[J]. 环境科学导刊, 2014,33(2):9-14. Wei X, Tang G M. Changes of nutrients and aquatic ecosystem of yilong lake in recent twenty decades [J]. Environmental Science Survey, 2014,33(2):9-14.
[49] 周虹霞,刘卫云,孔德平,等.异龙湖湿地大型水生植物群落特征分析[J]. 环境科学与技术, 2016,39(9):199-203. Zhou H X, Liu W Y, Kong D P, et al. Community features of aquatic macrophytes in wetland of Yilong Lake [J]. Environmental Science & Technology. 2016,39(9):199-203.
[50] 赵 磊.云南异龙湖生态系统稳态转换过程与关键灾变因子反演研究[D]. 北京:中国科学院大学.
[51] Appleby P G, Oldfield F. The calculation of lead-210 dates assuming a constant rate of supply of unsupported 210 Pb to the sediment [J]. Catena, 1978,5(1):1-8.
[52] Swain E B. Measurement and interpretation of sedimentary pigments [J]. Freshwater Biology, 1985,15(1):53-75.
[53] Meyers P A, Jane L T. Sediment organic matter. In: Tracking environmental change using lake sediments [M]. Dordrecht: Kluwer Academic Publisher, 2001:239-270.
[54] Hu K, Chen G J, Gregory-Eaves I, et al. Hydrological fluctuations modulate phototrophic responses to nutrient fertilization in a large and shallow lake of Southwest China [J]. Aquatic Sciences, 2019,81:1-17.
[55] Wieland E, Santschi P H, Höhener P, et al. Scavenging of Chernobyl137Cs and natural210Pb in Lake Sempach, Switzerland [J]. Geochimica et Cosmochimica Acta, 1993,57(13):2959-2979.
[56] Sommerfield C K, Nittrouer C A. Modern accumulation rates and a sediment budget for the Eel shelf: a flood-dominated depositional environment [J]. Marine Geology, 1999,154(1-4):227-241.
[57] Blais J M, Kalff J. The influence of lake morphometry on sediment focusing [J]. Limnology and Oceanography, 1995,40(3):582-588.
[58] Qin B Q, Liu Z W, Karl H. Eutrophication of shallow lakes with special reference to Lake Taihu, China [M]. Dordrecht: Springer Science & Business Media, 2007:176-178.
[59] 王教元,陈光杰,黄林培,等.洱海蓝藻爆发的时空特征及影响因子[J]. 生态学报, 2017,37(14):4831-4842. Wang J Y, Chen G J, Huang L P, et al. Spatio-temporal patterns and drivers of cyanobacteria blooms in Erhai Lake, Southwest China [J]. Acta Ecologica Sinica, 2017,37(14):4831-4842.
[60] Conley D J, Paerl H W, Howarth R W, et al. Controlling eutrophication: nitrogen and phosphorus [J]. Science, 2009,323(5917):1014-1015.
[61] Lewis Jr W M, Wurtsbaugh W A, Paerl H W. Rationale for control of anthropogenic nitrogen and phosphorus to reduce eutrophication of inland waters [J]. Environmental Science & Technology, 2011,45(24): 10300-10305.
[62] Paerl H W, Scott J T, McCarthy M J, et al. It takes two to tango: When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems [J]. Environmental Science & Technology, 2016,50(20):10805-10813.
[63] Huisman J, Codd G A, Paerl H W, et al. Cyanobacterial blooms [J]. Nature Reviews Microbiology, 2018,16(8):471-483.
[64] Paerl H W, Huisman J. Blooms like it hot [J]. Science, 2008,320 (5872):57-58.
[65] 郑维发,曾昭琪.淡水蓝藻的高温适应[J]. 湖泊科学, 1994,6(4): 356-363. Zheng W F, Zeng S Q. High temperature adaptation of fresh water cyanobacterium [J]. J. Lake Sci., 1994,6(4):356-363.
[66] 商兆堂,任 健,秦铭荣,等.气候变化与太湖蓝藻暴发的关系[J]. 生态学杂志, 2010,29(1):55-61. Shang Z T, Ren J, Qin M R, et al. Relationships between climatic change and cyanobacterial bloom in Taihu Lake [J]. Chinese Journal of Ecology, 2010,29(1):55-61.
[67] Kosten S, Huszar V L M, Bécares E, et al. Warmer climates boost cyanobacterial dominance in shallow lakes [J]. Global Change Biology, 2012,18(1):118-126.
[68] Taranu Z E, Zurawell R W, Pick F, et al. Predicting cyanobacterial dynamics in the face of global change: the importance of scale and environmental context [J]. Global Change Biology, 2012,18(12): 3477-3490.
[69] Whitton B A. Ecology of Cyanobacteria II: Their Diversity in Space and Time [M]. New York: Springer Science & Business Media, 2012: 216-222.
[70] Anderson N J, Bennion H, Lotter A F. Lake eutrophication and its implications for organic carbon sequestration in Europe [J]. Global Change Biology, 2014,20(9):2741-2751.
[71] Dietz R D, Engstrom D R, Anderson N J. Patterns and drivers of change in organic carbon burial across a diverse landscape: Insights from 116Minnesota lakes [J]. Global Biogeochemical Cycles, 2015,29 (5):708-727.
[72] 梁 红,黄林培,陈光杰,等.滇东湖泊水生植物和浮游生物碳、氮稳定同位素与元素组成特征[J]. 湖泊科学, 2018,30(5):1400-1412.Liang H, Huang LP, Chen GJ, et al.Patterns of carbon and nitrogen stable isotopes and elemental composition of lake primary producers and zooplankton in Eastern Yunnan [J]. J. Lake Sci, 2018,30(5):1400-1412.
[73] Meyers P A. Preservation of elemental and isotopic source identification of sedimentary organic matter [J]. Chemical geology, 1994,114(3/4):289-302.
[74] Stabel H H. Calcite precipitation in Lake Constance: Chemical equilibrium, sedimentation, and nucleation by algae 1 [J]. Limnology and Oceanography, 1986,31(5):1081-1094.
[75] Hodell D A, Schelske C L, Fahnenstiel G L, et al. Biologically induced calcite and its isotopic composition in Lake Ontario [J]. Limnology and Oceanography, 1998,43(2):187-199.
[76] Dittrich M, Obst M. Are picoplankton responsible for calcite precipitation in lakes? [J]. AMBIO: A Journal of the Human Environment, 2004,33(8):559-564.
[77] Riding R. Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic–Cambrian changes in atmospheric composition [J]. Geobiology, 2006,4(4):299-316.
[78] Obst M, Wehrli B, Dittrich M. CaCO3nucleation by cyanobacteria: Laboratory evidence for a passive, surface-induced mechanism [J]. Geobiology, 2009,7(3):324-347.
[79] Kamennaya N A, Ajo-Franklin C M, Northen T, et al. Cyanobacteria as biocatalysts for carbonate mineralization [J]. Minerals, 2012,2(4): 338-364.
[80] Strong A E, Eadie B J. Satellite observations of calcium carbonate precipitations in the Great Lakes [J]. Limnology and Oceanography, 1978,23(5):877-887.
[81] Apolinarska K, Pleskot K, Pełechata A, et al. Seasonal deposition of authigenic calcite out of isotopic equilibrium with DIC and water, and implications for paleolimnological studies [J]. Journal of Paleolimnology, 2021,66(1):41-53.
[82] Kelts K, Hsü K J. Freshwater carbonate sedimentation [M]//Lakes: Chemistry, Geology, Physics. Berlin: Springer-Verlag, 1978:295–323.
[83] He H, Wang Y, Liu Z, et al. Lake metabolic processes and their effects on the carbonate weathering CO2sink: Insights from diel variations in the hydrochemistry of a typical karst lake in SW China [J]. Water Research, 2022,222:118907.
[84] Zeng Q, Liu Z, Chen B, et al. Carbonate weathering-related carbon sink fluxes under different land uses: A case study from the Shawan Simulation Test Site, Puding, Southwest China [J]. Chemical Geology, 2017,474:58-71.
[85] Zeng S, Liu H, Liu Z, et al. Seasonal and diurnal variations in DIC, NO3-and TOC concentrations in spring-pond ecosystems under different land-uses at the Shawan Karst test site, SW China: carbon limitation of aquatic photosynthesis [J]. Journal of Hydrology, 2019,574:811-821.
[86] Yang R, Chen B, Liu H, et al. Carbon sequestration and decreased CO2emission caused by terrestrial aquatic photosynthesis: Insights from diel hydrochemical variations in an epikarst spring and two spring-fed ponds in different seasons [J]. Applied Geochemistry, 2015,63:248-260.
[87] Müller B, Meyer J S, Gächter R. Alkalinity regulation in calcium carbonate-buffered lakes [J]. Limnology and Oceanography, 2016, 61(1):341-352.
[88] Schelske C L, Hodeli D A. Recent changes in productivity and climate of Lake Ontario detected by isotopic analysis of sediments [J]. Limnology and Oceanography, 1991,36(5):961-975.
[89] Megard R O. Planktonic photosynthesis and the environment of calcium carbonate deposition in lakes [R]. Minneapolis, University of Minnesota, 1968.
[90] Wang S, Yeager K M, Wan G, et al. Dynamics of CO2in a karst catchment in the southwestern plateau, China [J]. Environmental Earth Sciences, 2015,73(5):2415-2427.
[91] Delille B, Harlay J, Zondervan I, et al. Response of primary production and calcification to changes ofCO2during experimental blooms of the coccolithophorid Emiliania huxleyi [J]. Global Biogeochemical Cycles, 2005,19(2):1-14.
[92] Zhu T, Dittrich M. Carbonate precipitation through microbial activities in natural environment, and their potential in biotechnology: A review [J]. Frontiers in Bioengineering and Biotechnology, 2016,DOI: 10.3389/fbioe.2016.0004.
[93] Hartnett H E, Keil R G, Hedges J I, et al. Influence of oxygen exposure time on organic carbon preservation in continental margin sediments [J]. Nature, 1998,391(6667):572-575.
[94] Ferland M E, Prairie Y T, Teodoru C, et al. Linking organic carbon sedimentation, burial efficiency, and long-term accumulation in boreal lakes [J]. Journal of Geophysical Research: Biogeosciences, 2014, 119(5):836-847.
[95] Wang W, Li S L, Zhong J, et al. Carbonate mineral dissolution and photosynthesis-induced precipitation regulate inorganic carbon cycling along the karst river-reservoir continuum, SW China [J]. Journal of Hydrology, 2022,615:128621.
[96] Teranes J L, McKenzie J A, Lotter A F, et al. Stable isotope response to lake eutrophication: Calibration of a high-resolution lacustrine sequence from Baldeggersee, Switzerland [J]. Limnology and Oceanography, 1999,44(2):320-333.
[97] Talbot M R, Kelts K. Primary and diagenetic carbonates in the anoxic sediments of Lake Bosumtwi, Ghana [J]. Geology, 1986,14(11): 912-916.
[98] Krause S, Liebetrau V, Löscher C R, et al. Marine ammonification and carbonic anhydrase activity induce rapid calcium carbonate precipitation [J]. Geochimica et Cosmochimica Acta, 2018,243:116-132.
[99] 蒋启财,刘 波,郭荣涛,等.湖相原生白云石的微生物成因机理探讨[J]. 古地理学报, 2017,19(2):257-269.Jiang Q C, Liu B, Guo R T, et al. Microbial mechanism of lacustrine primary dolomite [J]. Journal of Palaeogeography, 2017,19(2):257-269.
[100]Dupraz C, Reid R P, Braissant O, et al. Processes of carbonate precipitation in modern microbial mats [J]. Earth-Science Reviews, 2008,96(3):141-162.
[101]Finlay K, Vogt R J, Bogard M J, et al. Decrease in CO2efflux from northern hardwater lakes with increasing atmospheric warming [J]. Nature, 2015,519(7542):215.
Synergistic changes of organic and inorganic carbon burial in response to eutrophication in Yilong Lake over the past 100 years.
LI Ping1,2, CHEN Guang-jie1*, KONG Ling-yang1, HUANG Lin-pei1, WANG Xu1, YANG Guan-shao1, SUO Qi1, LI Jing1, WANG Lu1, HAN Qiao-hua1, ZHOU Qi1, GUO Wen1, LI Rui1, ZHANG Tao1
(1.Yunnan Key Laboratory of Plateau Geographical Processes and Environmental Changes, Faculty of Geography, Yunnan Normal University, Kunming 650500, China;2.School of Resource and Environment, Xichang University, Xichang 615013, China)., 2023,43(10):5389~5402
Carbon burial and its dynamic change in lakes play an indispensable role in the global carbon budget and residual land sink. Carbon sequestration in hard water lakes and other inland waters is deemed to be an important sink process owing to carbonate weathering coupled with aquatic photosynthesis, which stimulates continuous organic carbon and inorganic carbon burial. However, existing assessment paradigms are mainly based on organic carbon, lacking a comprehensive understanding of inorganic carbon burial and its synergistic change with organic carbon. Here, the history of carbon burial fluxes over the past century through multi-proxy and spatial sediment surveys of Yilong Lake in the karst area of Yunnan province was analyzed, in combination with210Pb and137Cs chronological profiles, for uncovering the patterns and mechanisms of synergistic changes in organic and inorganic carbon burial. The results showed that the allochthonous primary productivity in Yilong Lake has been significantly elevated due to nutrient input caused by watershed cultivation (e.g., enrichment of sediment nitrogen and phosphorus contents), with sediment pigment content increasing from 7.02μg/g in ~1900 to 38.99μg/g in ~2020. The accumulation rates of both organic and inorganic carbon showed a continuous trend of increasing and a high degree of synergistic changes (=0.99,<0.001). Regression analysis further showed that algal biomass was the main factor in driving the accumulation rates of both organic and inorganic carbon (2=0.33~0.91,<0.001). This indicated that algal photosynthesis and the "biological carbon pump" effect have largely controlled the processes of carbon burial. With the general decline of allochthonous watershed export (as evidenced by a decline of sediment magnetic susceptibility signal and C:N ratio) and a continuous increase of algal biomass, more allochthonous organic carbon can be transformed to authigenic carbonates (as evidenced in a sustained depletion of13CTIC) during the decaying and mineralization of organic matters, enhancing the synergistic increase in both organic and inorganic carbon burial. These results suggested a highly synchronous increase in the long-term fluxes of organic and inorganic carbon burial in response to lake eutrophication, highlighting the increasingly coupled processes in the burial of organic and inorganic carbon, and an increased rate in sediment carbon cycling and recycling. Therefore, under the continuing impact of human activities and climate change, refined assessments of carbon sink in karst lakes need to highlight the pattern of inorganic carbon cycling and its dynamic role in the flux and stock of terrestrial carbon.
eutrophication;carbon burial;inorganic carbon;synergistic changes;Yilong Lake
X524
A
1000-6923(2023)10-5389-14
2023-02-24
国家自然科学基金资助项目(42171072);云南省重点研发计划项目(202203AC100002-02);异龙湖高原浅水湖泊云南省野外科学观测研究站资助项目(202305AM070002);云南省John P. Smol院士工作站资助项目(202005AF150005)
* 责任作者, 教授, guangjiechen@ynnu.edu.cn
李 平(1987-),男,四川营山人,云南师范大学在读博士研究生,研究方向为湖泊环境与全球变化.发表论文7篇.717189582@qq.com.
李 平,陈光杰,孔令阳,等.近百年来异龙湖有机碳和无机碳埋藏响应水体富营养化的协同变化特征 [J]. 中国环境科学, 2023,43(10):5389-5402.
Li P, Chen G J, Kong L Y, et al. Synergistic changes of organic and inorganic carbon burial in response to eutrophication in Yilong Lake over the past 100years [J]. China Environmental Science, 2023,43(10):5389-5402.
猜你喜欢
海洋通报(2022年4期)2022-10-10 07:41:48
云南画报(2020年11期)2020-12-14 07:17:40
皮革制作与环保科技(2020年14期)2020-03-17 07:16:04
上海公路(2017年1期)2017-07-21 13:38:33
环境保护与循环经济(2017年10期)2017-03-16 03:16:20
创新作文(小学版)(2016年10期)2016-11-11 05:45:20
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:07
中国房地产业(2016年8期)2016-03-01 01:25:55
西南石油大学学报(自然科学版)(2015年4期)2015-08-20 09:05:12
食品工业科技(2014年11期)2014-03-11 18:16:00
