
珠江口大型底栖动物群落结构特征及影响因素
2023-10-24 03:18李纯厚李亚芳舒黎明肖雅元唐广隆
南方水产科学 2023年5期
孙 叶,刘 永,李纯厚,吴 鹏,李亚芳,舒黎明,林 琳,肖雅元,唐广隆
1.浙江海洋大学 水产学院,浙江 舟山 316000
2.中国水产科学研究院南海水产研究所/农业农村部南海渔业资源开发利用重点实验室/广东珠江口生态系统野外科学观测研究站/广东省渔业生态环境重点实验室,广东 广州 510300
3.西沙岛礁渔业生态系统海南省野外科学观测研究站/海南省深远海渔业资源高效利用与加工重点实验室/三亚热带水产研究院,海南 三亚 572018
海洋大型底栖动物是指生活周期的全部或大部分时间栖息在水体底部的水生动物群。大多栖息固着于岩石等牢固基体上,或是深埋于泥质、沙质等松软的底质中,具有区域性强、迁移能力弱等特点,对栖息环境的变化回避性弱[1]。大型底栖动物群落类群主要由环节动物、软体动物和棘皮动物等大型无脊椎动物组成[2]。河口生态系统物质循环的稳定性离不开大型底栖动物的参与,相关研究表明大型底栖动物通过生物沉降等活动在底层耦合及化学循环过程等方面发挥了重要作用[3]。此外,大型底栖动物分布广、食性杂,能够摄取水域中的有机碎屑、无机颗粒物等,从而有效降低水体富营养化水平,净化了水体环境,在水域环境健康状况评价上发挥了重要作用[4]。
珠江口作为珠江入海口,被称为广东最大的出海口,与黄河口、长江口并誉为中国的三大河口,在我国河口生态系统中占据重要地位[5]。珠江口具有得天独厚的河口自然资源,水网分布密集,包含了河口、红树林等多种生态栖息环境,物种多样性丰富,生态环境容量大,是粤港澳大湾区规划发展的海上生态健康屏障,同时也是进行地球科学和生态学综合研究的重点区域[6]。近年来陆续对珠江口大型底栖动物群落开展了一系列研究[7-8],但有关珠江口大型底栖动物群落结构季节性变化特征及其与环境因子相关性的调查仍较少。本文探究了珠江口大型底栖动物群落的季节性差异,并揭示其季节演变规律,以为维护珠江口生态系统的稳定以及为其他栖息生物对河口自然资源的利用提供基础资料。
1 材料与方法
1.1 研究区域
分别于2019 年12 月 (冬季)、2020 年8 月 (夏季)、2021 年3 月 (春季) 和10 月 (秋季) 在珠江口海域进行了4 个航次的大型底栖动物生态学调查。共设置了20 个大面调查站点 (图1,表1),由于时间安排、天气条件和技术等问题,少数站位(A17、B2、B6、C1、C2、C5) 未能进行连续采样。
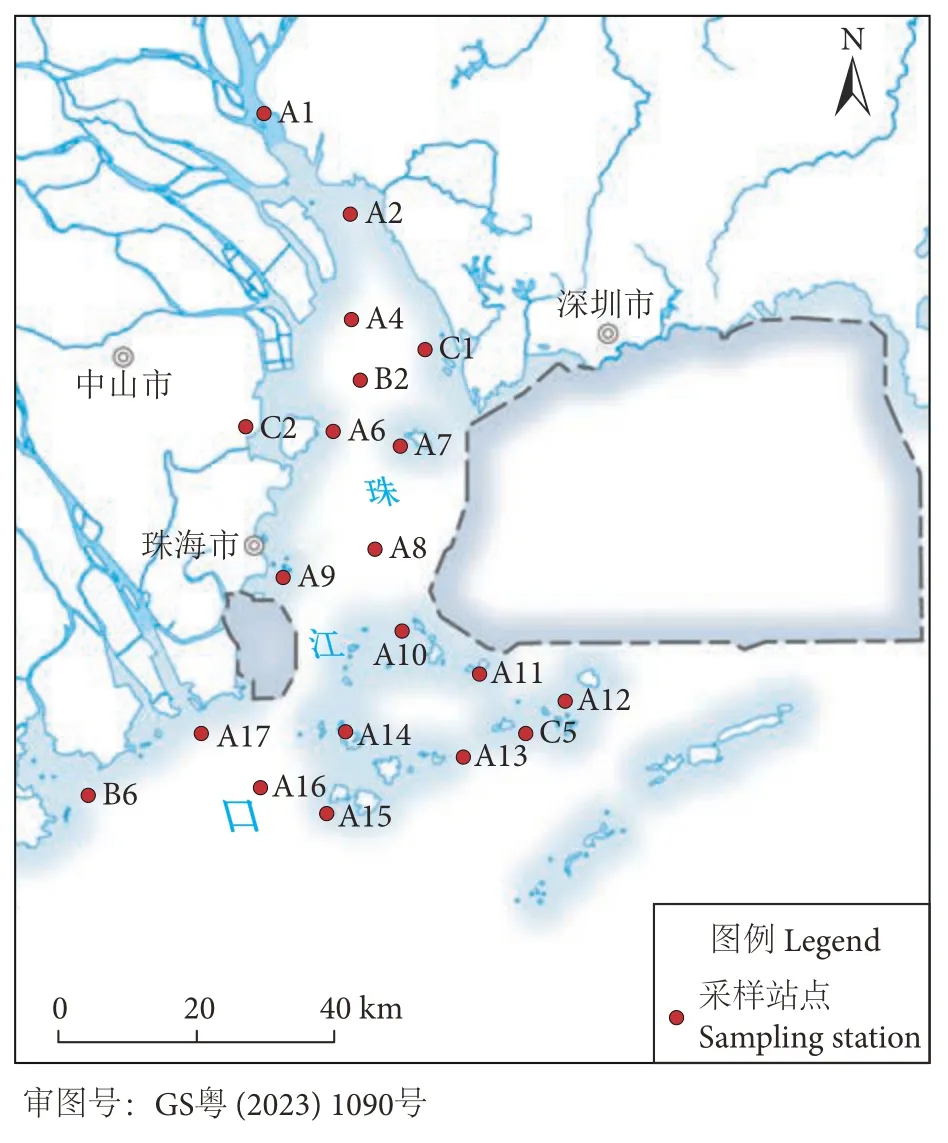
图1 珠江口大型底栖动物采样站位图注:因C2 站点靠近岸边,选择在涨潮期进行监测。Fig.1 Sampling stations of macrobenthos in Pearl River EstuaryNote: C2 station is close to the shore,so the monitoring was carried out during the high tide period.
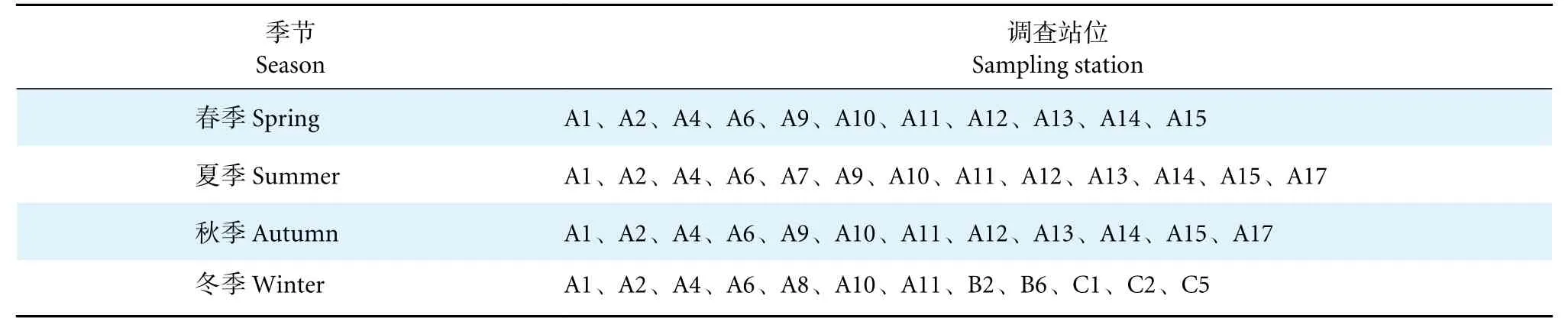
表1 珠江口大型底栖动物各季节的调查站位Table 1 Sampling stations of macrobenthos in Pearl River Estuary in different seasons
1.2 大型底栖动物采集
用0.1 m2的彼得森采泥器在同一站点进行2 次重复采集,过60 目的筛网[9]。两次采样所得样品作为一个数据,将采集的所有样品用75% (体积分数) 的乙醇固定后带回实验室,对样品进行种类鉴定、计数和称质量 (0.001 g 天平),将采集个体的数量与质量除以对应采样面积即可得到各个大型底栖动物的栖息密度与生物量[10-11]。
1.3 环境参数的测定
水温、溶解氧 (Dissolved oxygen,DO)、pH 和化学需氧量 (Chemical oxygen demand,COD) 等环境因子由便携式水质分析仪进行现场检测,用容积为5 L 的有机玻璃采水器采集海域水样,水深<5 m 仅采集表层 (表下0.5 m) 的水样;当水深介于5~25 m 时,分为表层和底层,底层距离海底2 m。活性磷酸盐 (PO43-) 等指标根据《海洋监测规范》(GB 17378—2007 和GB 17378.4—2007) 测定。
1.4 数据分析
大型底栖动物群落的Peilou 均匀度指数 (J')、Margalef 丰富度指数 (D) 和Shannon-Wiener 多样性指数 (H') 的计算公式为:
式中:S为所采大型底栖动物的总种类数;N为大型底栖动物总数量;Pi为第i种物种个体数量占获得大型底栖动物数量总和的比例。
将大型底栖动物的优势度指数作为优势种的判断依据,计算公式为:
式中:Y为优势度;fi表示第i种在调查点中出现的频率;pi为第i种在大型底栖动物总数量中的比例。若Y>0.02,即将该物种认定为优势种[12]。
运用SPSS 22.0 软件分别进行珠江口海域4 个季节大型底栖动物群落多样性的单因素方差分析(One-way ANOVA) 和大型底栖动物的群落多样性指数、丰富度指数等与环境因子的Pearson 相关性分析。运用PRIMER 6.0 软件对大型底栖动物群落结构进行聚类分析。
2 结果
2.1 大型底栖动物物种组成和季节性差异
对珠江口水域大型底栖动物开展4 个航次调查,共采集到大型底栖动物138 种,隶属于9 门。各类群大型底栖动物物种组成以软体动物种数最多,环节动物次之,刺胞动物和螠虫动物在本次调查中的种数最少 (表2)。
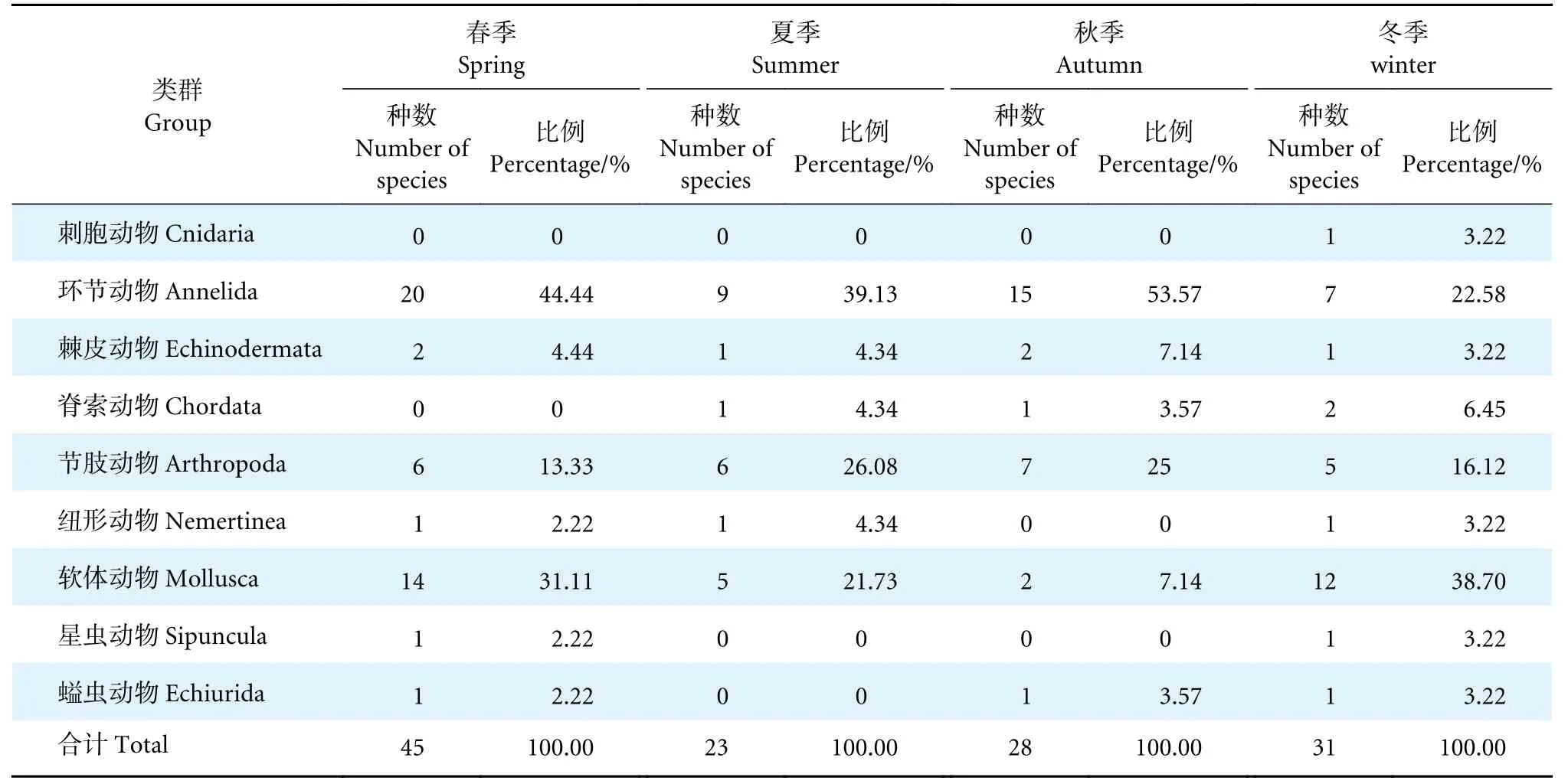
表2 珠江口大型底栖动物各类群组成的季节分布Table 2 Seasonal distribution of group composition of macrobenthos in Pearl River Estuary
大型底栖动物群落在各季节呈现的种类组成各不相同。春季种类最多,夏季最少,仅为春季的一半多。春、夏和秋季均以环节动物出现频率最高,其次为软体动物、节肢动物等,而刺胞动物和星虫动物仅在个别季节中出现。
2.2 优势种
由表3 可知,4 个航次采集获得的大型底栖动物中共发现6 种优势种。其中,春季珠江口海域有2 种优势种,为光滑河篮蛤 (Potamocorbula laevis) 和光滑倍棘蛇尾 (Amphiopholislaevis),分别属于软体动物和棘皮动物;夏季珠江口海域有3 种优势种,为裸盲蟹 (TyphlocarcinusnudusStimpson)、光滑倍棘蛇尾和角海蛹 (OphelinaacuminateOersted),分别属于节肢动物、棘皮动物和环节动物;秋季有2 种优势种,为背蚓虫 (Notomastus latericeus) 和光滑倍棘蛇尾,分别属于环节动物和棘皮动物;冬季优势种仅有短角双眼钩虾 (Ampeliscabrevicornis)。优势物种的季节性类群差异性较大,棘皮动物为珠江口海域调查的主要优势类群。光滑倍棘蛇尾为春、夏和秋三季共有优势种,其优势度在夏季达到最高值 (0.05),在各个季节优势地位明显 (表1)。
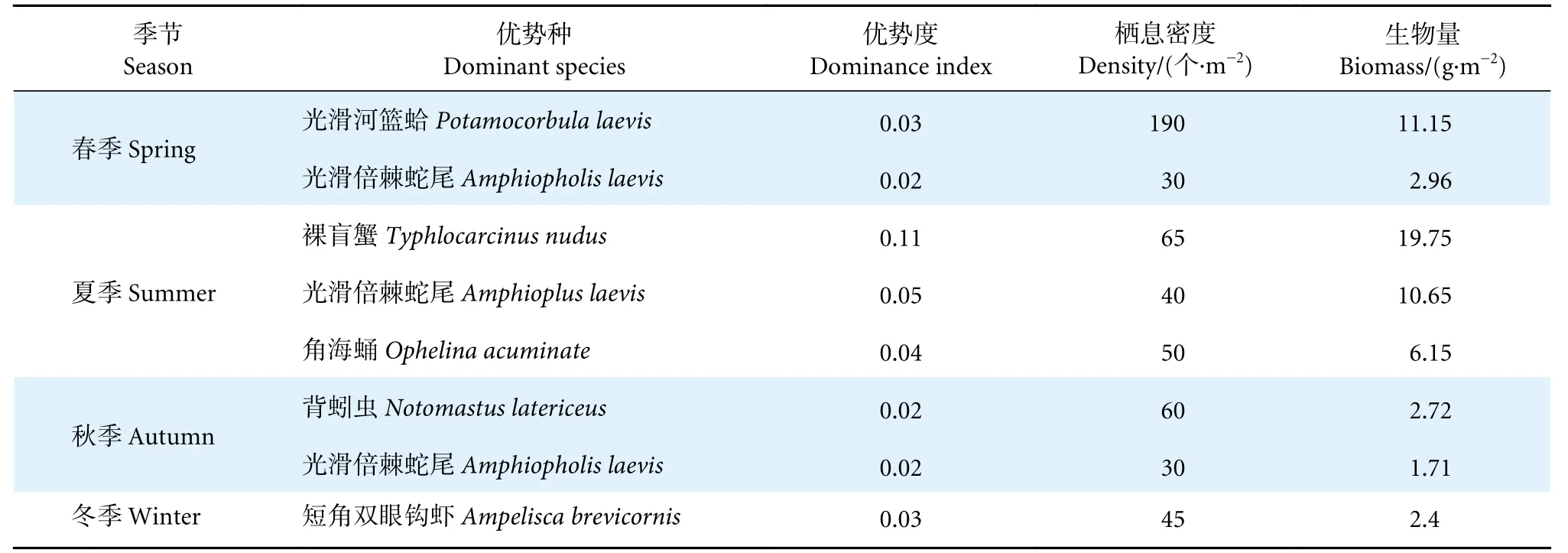
表3 珠江口大型底栖动物优势种及优势度分布Table 3 Distribution of dominant species and dominance of macrobenthos in Pearl River Estuary
2.3 生物量和栖息密度
珠江口海域大型底栖动物的栖息密度与生物量的季节变化如图2 所示,栖息密度平均值在春季最高 (590 个·m-2),冬季次之 (510 个·m-2),秋季为455 个·m-2,夏季最低 (375 个·m-2)。珠江口水域大型底栖动物生物量的平均值在冬季最高 (263.9 g·m-2),其次为春季 (139.88 g·m-2),夏季为72.55 g·m-2,秋季最低 (56.46 g·m-2)。
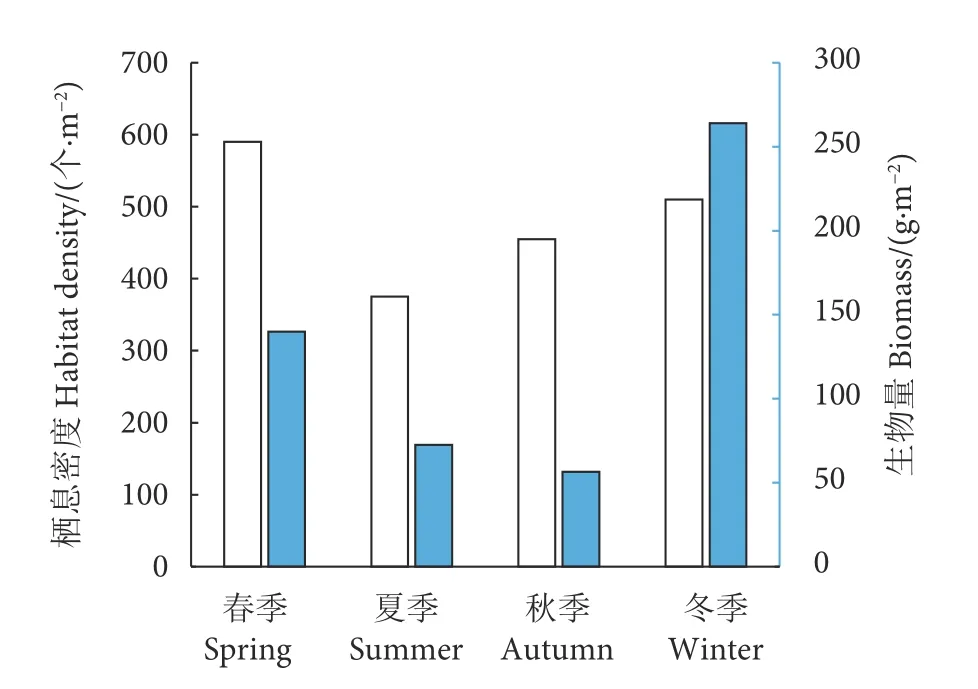
图2 珠江口大型底栖动物栖息密度和生物量的季节变化Fig.2 Seasonal variation of total habitat density and total biomass of macrobenthos in Pearl River Estuary
对四季不同类群大型底栖动物的生物量与栖息密度进行统计分析,结果如表4 所示。环节动物的栖息密度在秋季达到最高值 (300 个·m-2),生物量在夏季达到最高值 (14.2 g·m-2);棘皮动物的栖息密度在春季达到最高值 (70 个·m-2),生物量在夏季达到最高值 (31.26 g·m-2);软体动物的栖息密度和生物量的最高值均出现在春季,分别为190 个·m-2和83.56 g·m-2。其他大型底栖动物类群的栖息密度和生物量最高值均出现在冬季,分别为195个·m-2和158.25 g·m-2。各季节环节动物的栖息密度始终处于较高水平,生物量处于较高水平的物种则为软体动物。

表4 珠江口大型底栖动物主要类群栖息密度和生物量季节变化Table 4 Seasonal variation of habitat density and biomass of main groups of macrozoobenthos in Pearl River Estuary
对珠江口大型底栖动物四季的栖息密度进行聚类分析,结果如图3 所示。春季,A6 与A12 站位,A1 与A13 站位,A2 与A4 站位间的相似度较高;A6、A12 站位同其他站位间的相似度较低。夏季,A7 与A12 站位,A17、A10 与A14 站位,A2 与A6 站位间的相似度较高,而A1 与A4 站位、A2 与A6 站位,同其他调查站位之间的相似度较低。秋季,A15、A10 与A12 站位,A1 与A2 站位,A11 与A14 站位,A4 与A6 站位间的相似度较高;A13 站位除了与A4、A6 站位间的相似度较高外,与其他站位的相似度均较低。冬季,C1、A1 与A4 站位,B6 与C2 站位,A10 与A11 站位,C5、A2 与B2 站位间的相似度较高。
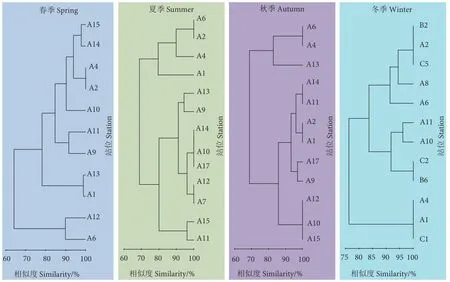
图3 珠江口大型底栖动物四季的栖息密度聚类分析Fig.3 Cluster analysis of habitat density of macrobenthos in Pearl River Estuary in four seasons
2.4 大型底栖动物多样性季节变化
珠江口海域大型底栖动物各季节H'、J'和D结果如表5 所示。H'和J'的最大值均出现在冬季,分别为1.66±0.15 和0.97±0.03;D最大值出现在春季(2.36±0.83)。春季J'最低(0.18±0.07),而秋季H'最低 (0.29±0.16)。
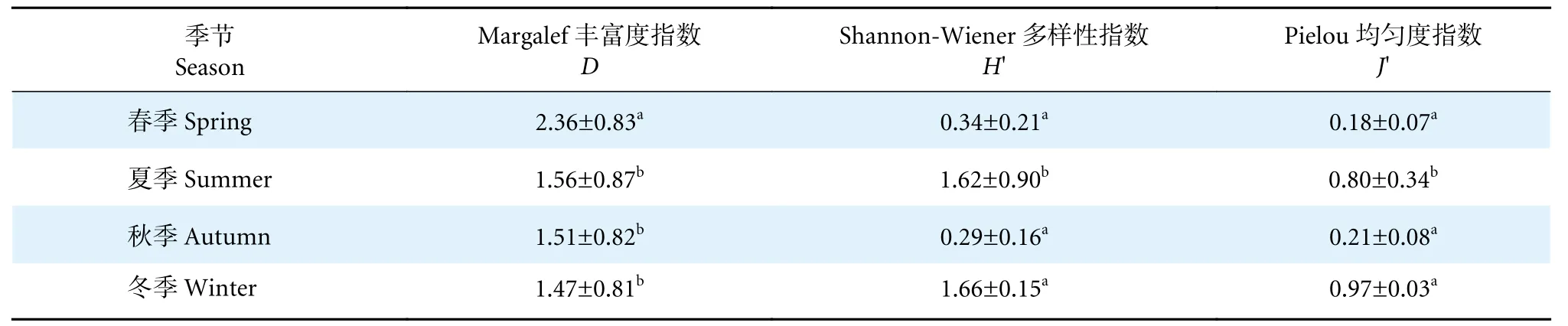
表5 珠江口大型底栖动物多样性指数季节分布Table 5 Seasonal distribution of macrobenthos diversity index in Pearl River Estuary
2.5 大型底栖动物群落与环境因子的相关性
各季节全部调查采样点的多样性指数 (H'、D和J') 与环境因子的Pearson 相关性分析结果显示,大型底栖动物H'与pH 呈极显著正相关性(P<0.01),与盐度则呈显著正相关性,同COD 呈极显著负相关性 (P<0.01),与PO43-呈显著正相关性。J'与pH、PO43-呈极显著正相关性,和COD 呈极显著负相关性,与悬浮物呈显著正相关性 (P<0.05);D与温度和PO43-呈显著负相关性,与盐度呈极显著正相关性 (表6)。
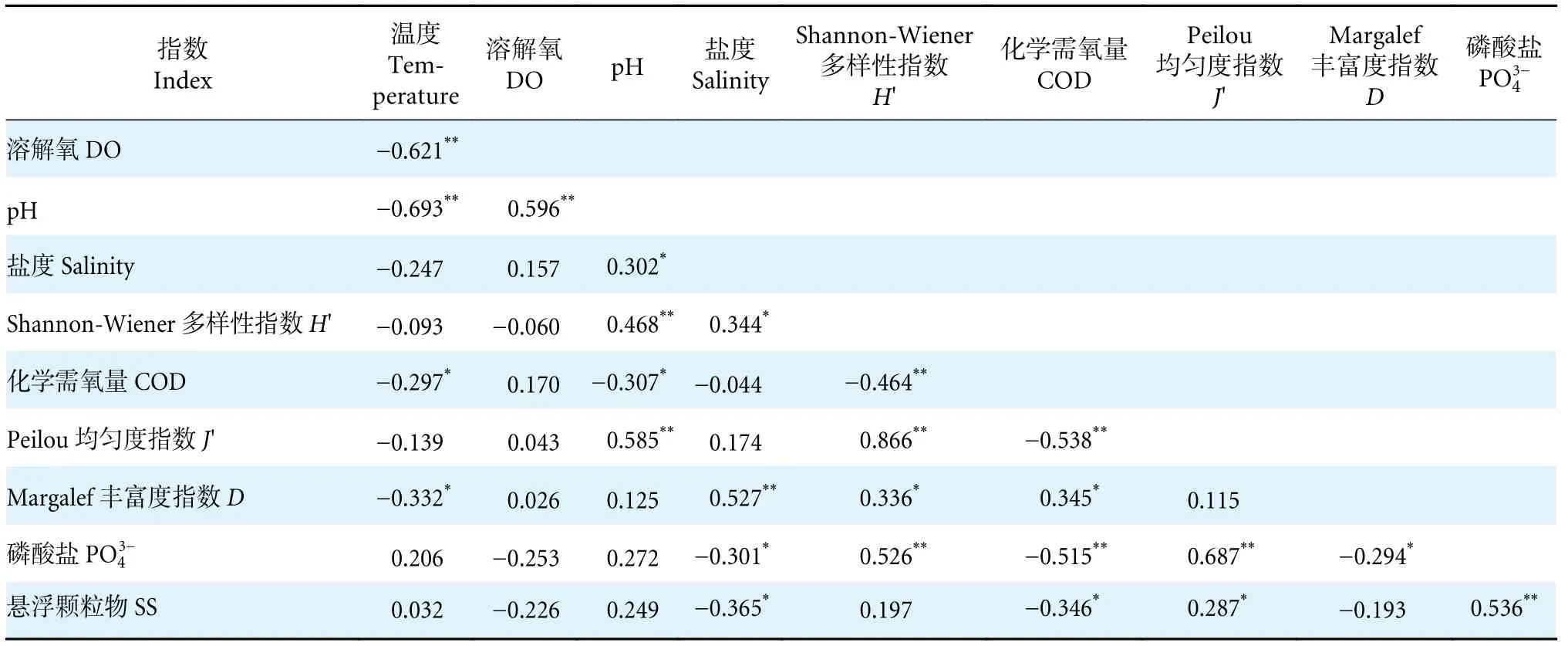
表6 珠江口大型底栖动物物种多样性指数与环境因子间的相关性Table 6 Correlation analysis of macrozoobenthos diversity index and environmental factors in Pearl River Estuary
3 讨论
3.1 珠江口大型底栖动物群落结构的变化特征
将本研究4 个航次大型底栖动物的调查结果同珠江口及周边海域的历史数据进行比较,结果见表7。可以看出,本研究中大型底栖动物的物种数为2008 年以来的最高值,可能是由于本次调查频次相较之前增加,故所获大型底栖动物的物种数增多;但与2004—2007 年相比,物种数却处于较低值,这可能是由于近年来珠江口海域出现了季节性的底层海水大范围缺氧现象[26],使得该海域大型底栖动物的生境被破坏,导致其种类和数量直线下降。而2010 年以后,随着环境生态保护措施投入的增加和海水污染源的监控,珠江口海域DO 处于相对稳定水平,因此大幅提高了大型底栖动物的生境稳定性[27]。
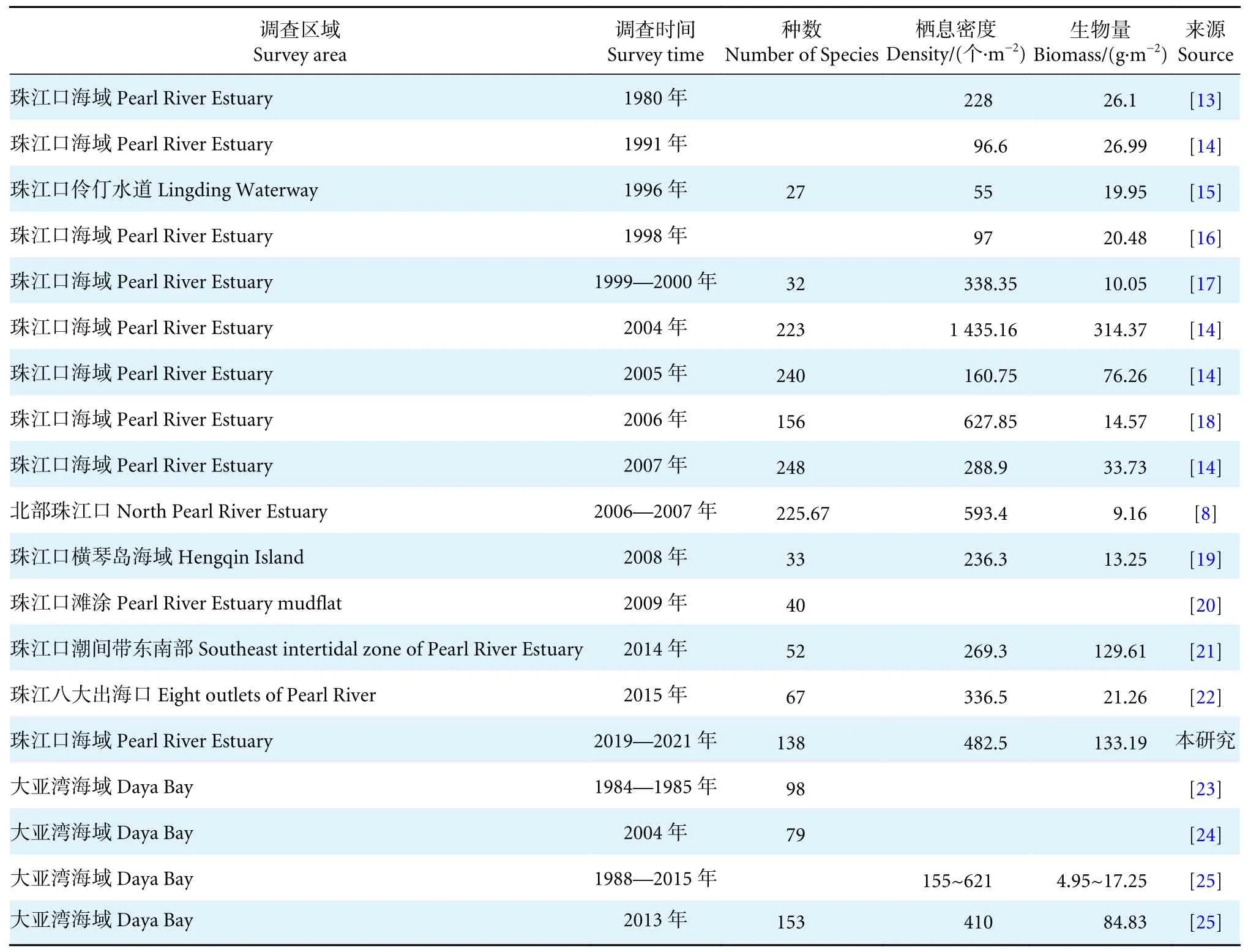
表7 本次调查与周边海域的比较Table 7 Comparison of results from this investigation and surrounding waters
在丰度和生物量方面,本研究的平均栖息密度为2008 年调查以来的最高值,大型底栖动物生物量也处于上升趋势。这可能是由于各航次采样调查点位于珠江口不同海域,大型底栖动物的栖息环境不同,本次调查点位于珠江口的外部海域,而2015 年的调查点主要位于珠江口八大入海口区域内。此外,两次大型底栖动物调查使用不同孔径底层筛[28],也可能导致密度与生物量存在一定差异。而同时间段的大亚湾海域调查结果显示大型底栖动物的栖息密度和生物量呈下降趋势[25]。这可能是由于2015 年后调查主要位于大亚湾西北部区域,该片区紧邻大亚湾工业集中区,相对湾口受人类干扰的影响更显著。近年来陆续开展的航道疏浚工程和填海工程产生的大颗粒悬浮物对大亚湾西北片区海域造成了严重污染[23-24],对大型底栖动物的生存环境造成较大影响,破坏了其生理过程的稳定性。同时随着环境污染加重,水体富营养化使得该海域赤潮现象频发,致使DO 浓度降低,大型底栖动物的饵料来源减少,从而导致大型底栖动物栖息密度和生物量降低[29]。
本次调查结果显示,软体动物为各类群中对生物量贡献最大的种类,主要优势种为光滑河篮蛤、波纹巴非蛤 (Paratapesundulatus) 和菲律宾蛤仔(Ruditapesphilippinarum) 等。这些双壳贝类数量庞大[30],以群居为主,主要栖居在近岸泥沙混合底质[31]。有研究表明光滑河篮蛤是珠江口春、秋季的优势种且高值区在春季[32-33],而本研究结果显示光滑河篮蛤在夏季的优势度远高于春、秋两季,这可能由不同调查区域降雨量差异所致。本次调查中夏季降雨量较以往增加,导致海水盐度降低,海水盐度稳定在5‰~25‰,促进了光滑河篮蛤的繁殖与生长,从而使其的夏季出现频率高于春季[34]。
本研究中珠江口大型底栖动物春季的均匀度指数和多样性指数均低于夏、秋季,与田伟等[34]对椒江口的调查结果一致。春季珠江口大型底栖动物调查期间处于枯水期,富营养化水平较高[35]。在沉积物有机质含量较高、沉积环境不稳定的条件下,调查海域的生态环境处于亚健康状态。随之而来的水体富营养化造成季节性缺氧现象发生,影响了大型底栖动物的生理过程,从而使大型底栖动物种数剧减、多样性降低[36]。因此,珠江口沉积物中高含量的有机质可能是造成该海域大型底栖动物多样性较低的重要原因之一[37]。
3.2 珠江口环境状况对大型底栖动物分布的影响
根据本研究中大型底栖动物群落多样性与环境因子的相关性结果,盐度是影响其群落多样性变化的关键因素。已有研究发现,盐度梯度性变化是河口海域的重要特征之一[38]。河口生态系统处于动态变化过程中,盐度是影响河口生物分布的主要限制因素之一,且盐度介于30‰~34‰的大型底栖动物群落的物种多样性较高;若盐度介于5‰~15‰,则会出现大型底栖动物群落种类较少、丰富度较低的现象。本文研究区域的盐度主要介于25‰~35‰,相较于以往调查区域处于较高水平,因而大型底栖动物物种数相对较多,多样性较为丰富。
大型底栖动物群落结构与环境因子的分析结果显示,悬浮物对大型底栖动物群落的丰富度指数也产生了一定影响。秦蕴珊和李凡[39]的调查表明,海水中悬浮物含量与颗粒大小会影响海水透明度,当悬浮物含量处于较低水平时,海水透明度呈现较高值[40]。海水中的悬浮物大多要沉降到海底,当悬浮物含量较高、颗粒较大时,下沉速度处于较慢水平,则会影响光在海水中的传播。这对海域中浮游植物群落的光合作用会产生干扰,阻碍其繁殖与生长,导致海水生态系统的初级生产效率降低、浮游植物群落丰富度下降,无法为大型底栖动物提供充足饵料,进而使其群落物种数、生物量和群落多样性降低。但本研究中Pearson 相关性分析显示,大型底栖动物多样性指数与悬浮物含量之间未表现出相关性 (P>0.05),可能是近年来珠江口海域大型底栖动物群落的栖息环境发生了一些变化,多种复杂的生态环境和底质变化,掩盖了悬浮物含量对大型底栖动物多样性的影响。
本研究还发现,温度与大型底栖动物丰富度指数之间存在显著相关性,河口区的温度在一定程度上影响着底栖生物的定着、栖息和活动。张景平等[24]研究表明,温度与大型底栖动物群落多样性具有显著相关性,本研究调查的结果与其一致。在一定范围内,适当提升温度能够促进大型底栖动物的繁殖和发育,提高其群落生产力。在盐度、磷酸盐、pH 等环境要素保持适宜的前提下,摇蚊幼虫(Chironomusriparius) 在春夏季温暖的栖息环境下表现为较高的丰富度,而在冬季发育缓慢,生物量较低。若温度临界值超过其适应性范围,则会破坏其生长环境的平衡性,导致其大量死亡[41]。
珠江口大型底栖动物群落的栖息环境在近20 年发生了一定变化,多种影响因子间相互作用使得本次调查与以往研究调查的大型底栖动物在物种数、生物量等方面存在一定差异性,这说明了大型底栖动物采样调查中影响因子的波动性和偶然性。
4 小结
1) 4 个季节采样调查所获的大型底栖动物共计138 种,其中软体动物的种类最多 (50 种)。春季出现的大型底栖动物种数丰富度高于夏、秋、冬三季,夏季的物种数最少,仅为春季的一半多。各季节的优势种组成差异较大,然而,光滑倍棘蛇尾是珠江口海域的主要优势种,为春、夏、秋三季的共有优势种。
2) 4 个航次调查的大型底栖动物的栖息密度平均值在春季最高 (590 个·m-2)。就类群组成而言,秋季环节动物的栖息密度平均值为300 个·m-2。大型底栖动物4 个季节生物量的平均值为532.79 g·m-2,季节性差异大,最高值出现在冬季,秋季最低。
3) 从大型底栖动物多样性与环境因子的相关性分析来看,多样性指数、丰富度指数和均匀度指数受多种因子的共同作用,本研究海域的盐度处于适宜生物群落栖息的范围内 (25‰~35‰),故而大型底栖动物的群落丰富度较高、种类数较多,同时海水中悬浮物颗粒含量与大小也会影响大型底栖动物栖息环境的平衡。需进一步加强水域环境的健康监测,提高大型底栖动物群落生态系统的稳定性。
猜你喜欢
水文地质工程地质(2022年2期)2022-04-13
今日农业(2021年8期)2021-11-28
中国宝玉石(2021年5期)2021-11-18
水上消防(2021年3期)2021-08-21
石油化工建设(2020年1期)2020-08-24
人大建设(2019年5期)2019-10-08
新城乡(2018年11期)2018-11-22
海洋信息技术与应用(2017年2期)2017-06-21
海洋渔业(2016年6期)2016-04-16
中国海洋大学学报(自然科学版)(2014年8期)2014-02-28
