
延河流域不同生活型植物功能性状特征及其对环境变化的响应
2023-10-23 03:14朱晓旭温仲明张格语王维维刁晓晨万锦枫
水土保持研究 2023年6期
朱晓旭, 温仲明,, 郑 诚, 高 原, 张格语, 王维维, 刁晓晨, 万锦枫
(1.西北农林科技大学 草业与草原学院, 陕西 杨凌 712100; 2.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100; 3.中国科学院大学, 北京 100049)
植物功能性状是表征植物生长发育、繁殖及一系列生命活动的标志性特征,是衡量植物环境适应性的重要指标[1]。同时,植物功能性状也是生态系统功能在个体水平上的映射,能够在个体水平上揭示群落乃至更大尺度的生态问题。植物功能性状由其物种生物遗传和环境因子共同作用形成,植物功能性状一定程度上也对环境变化有预测意义。开展植物功能性状特征及其对环境变化响应研究对探究植物生长策略和植被恢复等生态问题有着重要意义[2]。
生活型是植物长期适应环境而反映在外部形态、结构、性状上的分类类型,是生态学研究的重要分类单位[3]。不同的生活型植物功能性状不同,对环境变化的响应也存在差异。同一生活型物种采取相近的生存策略,亲缘性不高的物种在邻近的地理位置也可以表达出相似的外部形态和功能性状[4]。植物生活型影响着生境的物质循环和能量流动,一方面生活型对植物养分重吸收的影响不同,乔木植物具有更强的碳同化能力而草本植物叶片通过比乔木植物更强的N,P吸收能力推动着氮、磷循环,另一方面不同生活型植物叶片叶绿素含量均值存在差异,叶绿素含量则直接影响植物的光合作用速率[5]。植物生活型物种数量的动态变化也反映出生境的群落演替过程,譬如在保护或还林超过3 a的群落中,灌木和乔木的物种数量、种子数量占比会大幅增加占据绝对优势。在生态学研究中,植物生活型可以直观地表征群落外貌以及群落的垂直结构,是研究植物功能性状变异的重要工具。
不同生活型植物在功能性状表现上存在差异,如覃张芬等[6]在热带珊瑚岛进行的研究发现该区域乔木、草本植物的比叶面积、叶氮含量均显著高于灌木,又如黄小等[7]在湖北星斗山进行的研究发现该区域灌木植物比叶面积高于草本和乔木,草本植物叶氮含量高于乔木和灌木。叶经济谱理论和根经济谱理论的出现让植物生活型成为了植物生存策略选择的重要解释工具,如张姗姗等[8]在苏州留园所作的研究显示,不同生活型植物的叶性状间存在显著差异,草本生活型为获取型生存策略,灌木、乔木生活型为保守型生存策略。又如周汀等[9]在贵州茂兰保护区所作的研究显示,不同生活型植物对应在了叶经济谱的不同位置,乔木植物采取保守型生存策略而灌木植物采取获取型生存策略。
黄土丘陵区是我国水土流失与生态问题最严重的区域之一。为控制水土流失,该区先后实施了一系列的生态恢复项目,尤其是1999年实施的退耕还林(草)工程,使该区植被覆盖度和物种多样性水平显著提高,水土流失得到明显控制,生态环境得到极大改善。但在植被恢复过程中,也存在物种选择不当、恢复植被超过了立地环境承载力、土壤干层发育广泛、植被可持续性较差等问题。导致退耕还林(草)工程边际效应递减、生态成效不持续等新问题的出现[10]。近些年来国内关于植物功能性状响应环境因子在内蒙古[11]、新疆[12]等地的研究被大量报道,涵盖了森林生态系统[13]、草地生态系统[14]、荒漠生态系统[15]在内的大部分陆地生态系统。但在延河流域从生活型角度分析植物功能性状特征及其对环境因子响应的研究还较少。为此,本研究以黄土丘陵区延河流域3种植物生活型为研究对象,调查6种植物功能性状,旨在回答如下两个问题:(1) 延河流域不同生活型植物功能性状特征是否存在差异,如何影响不同生活型植物的生存策略;(2) 不同环境因子影响植物适应生境的方式、强度有什么不同。
1 材料与方法
1.1 研究区概况
本研究选择黄土丘陵区延河流域(36°50′±0°27′N,109°36′30″±0°51′30″E)为研究区,该流域位于黄土高原中部,平均海拔为1 371.9 m,年平均温度为8.8℃,年降雨量为505 mm。区域内黄绵土分布广泛,土质疏松、质地单一、抗侵蚀能力差,呈现黄土塬梁丘陵沟壑地貌,生态环境脆弱。延河流域降雨量自东南向西北方向递减,植被呈地带性分布,主要植物生活型分为草本、灌木、乔木3种。常见草本有铁杆蒿(ArtemisiastechmannianaBess.)、达乌里胡枝子〔Lespedezadavurica(Laxmann) Schindler〕、长芒草(StipabungeanaTrin.)、白羊草〔Bothriochloaischaemum(Linnaeus) Keng〕等,常见灌木有杠柳(PeriplocasepiumBunge)、狼牙刺〔Sophoradavidii(Franch.) Skeels〕、柠条(CaraganakorshinskiiKom.)等,常见乔木有鸡爪槭(AcerpalmatumThunb.)、辽东栎(QuercusmongolicaFisch. ex Ledeb.)、杜梨(PyrusbetulifoliaBunge)等[16]。
1.2 样地设置、野外调查与样本采集
为分析延河流域不同生活型植物功能性状特征及其对环境变化的响应,本研究的采样点尽量能覆盖较大的环境梯度(图1)。在每个采样点设置4~6个样方,乔木样方面积为10 m×10 m,灌木样方面积为5 m×5 m,草本样方面积为1 m×1 m。将所采样方的物种分为草本、灌木、乔木3种生活型(表1)。

表1 延河流域采样物种生活型划分Table1 Divison of plant life form
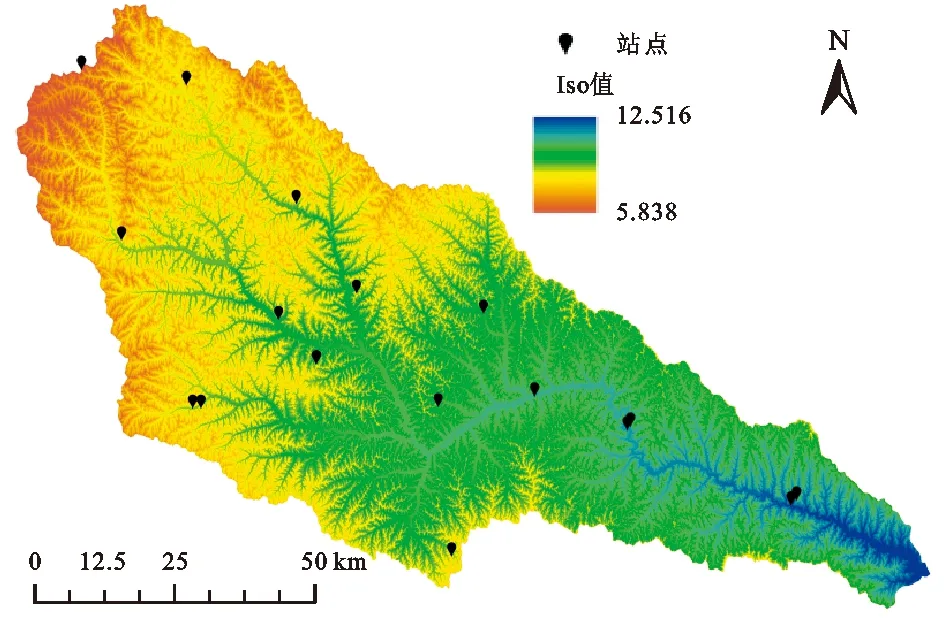
图1 研究区采样点分布Fig. 1 Sampling sites
1.3 性状测定
本研究主要选择的植物功能性状有比叶面积(SLA)、叶组织密度(LTD)、比根长(SRL)、根组织密度(RTD)、叶氮含量(LNC)、根氮含量(RNC)。比叶面积(SLA)主要表征植物对光能的捕获能力和潜力[17]。比根长(SRL)主要表征植物根系对土壤中高肥斑块的觅养能力[18]。叶组织密度(LTD)、根组织密度(RTD)主要表征植物叶片、细根的抗拉力以及防御能力。叶氮含量(LNC)、根氮含量(RNC)主要表征植物氮固定生理过程的强度,量化植物吸收氮的能力,反映植物资源获取与营养代谢的动态平衡[19]。
样本测量方法:在所选样方内按照五点取样法选取自然生长且长势良好的铁杆蒿、达乌里胡枝子、阿尔泰狗娃花、长芒草、白羊草、黄刺玫、杠柳、丁香、连翘、荆条、狼牙刺等植株,对所选植株按照东西南北的顺序采取10~15片感光良好、大小基本一致、表面完全展开且未感染病虫害的叶片,并用铁铲挖取所选植株根系长度对应深度的土块,清掉土和杂质后,采集直径<2 mm的细根,所采叶片和细根样品均按照物种分类并装入塑封袋编号,而后放入车载冰箱存放。(1) 叶面积。先用扫描仪扫描平展的叶片获得图像,再用Image-J软件根据扫描图像的像元数得到叶面积;(2) 叶干质量。在高温条件(105℃)下将叶片杀青15 min,烘干(60±12) h(85℃),用万分之一精度天平称重得到叶干重;(3) 叶片厚度。使用电子游标卡尺(精度0.01 mm),三点式取点法沿叶片的主脉方向均匀选取3个点,且3个点距离叶片主脉两侧距离约0.25 cm,用游标卡尺测其厚度d1,d2,d3,则叶片的厚度为LT=(d1+d2+d3)/3;(4) 根长。根洗净并分离出细根(直径<2 mm),用精度为0.01 mm的电子游标卡尺测量细根的长度直径;(5) 根体积。排水法将洗净擦干的细根完全浸入盛水量筒5 s,读取根系体积;(6) 根干重。细根105℃杀青15 min后85℃烘干(60±12) h称重,读取根干重;(7) 叶氮(LNC)、根氮(RNC)。将烘干的叶片和细根粉碎,测定叶氮(LNC)、根氮(RNC)含量(单位:mg/g)[20]。
性状计算方法:比叶面积(SLA)=叶面积/叶干质量(cm2/g);叶组织密度(LTD)=叶干质量/叶体积(g/cm3);比根长(SRL)=根长度/根干质量(m/g);根组织密度(RTD)=根干质量/根体积(g/cm3)。
1.4 环境数据提取
对照采样点的经纬度信息,使用ArcGIS,从延河流域环境因子专题图中提取年降雨量(MAP)、年均温(MAT)、雨季降雨量(Rain789)等气候数据,其他地形因子数据从野外调查中获取。
1.5 数据处理
采用单因素方差分析(ANOVA)对延河流域不同生活型植物功能性状进行差异分析,并对性状关系做线性回归分析,再采用冗余分析(RDA)研究植物对环境变化的响应。数据分析中为符合统计学要求,对原始数据做标准对数化处理(lgdata/lndata)[21]。
2 结果与分析
2.1 延河流域不同生活型植物功能性状特征
延河流域不同生活型植物功能性状特征分析结果显示(图2—3)植物功能性状的值在统计分析中的变化范围如下,SLA:2.77~5.32;LTD:-4.17~-2.23;LNC:-0.06~1.82;SRL:2.19~4.68;RTD:-2.52~0.77;RNC:-0.84~1.29。草本SLA,LNC,SRL,RNC显著高于灌木和乔木,LTD和RTD显著低于灌木和乔木。灌木SLA,SRL,LTD,LNC,RTD,RNC与乔木无显著性差异。
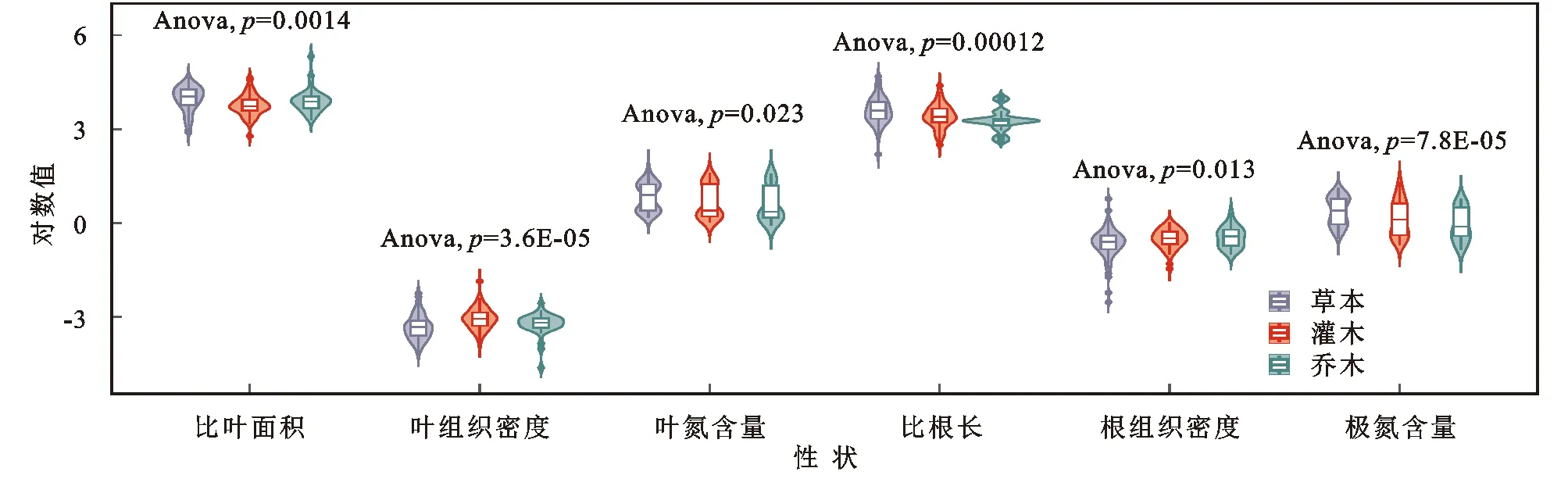
注:比叶面积单位为cm2/g;叶组织密度单位为g/cm3;比根长单位为m/g;根组织密度单位为g/cm3;叶氮含量、根氮含量单位为mg/g。图2 延河流域不同生活型植物功能性状组间差异Fig. 2 Characteristics differences between groups of functional traits of plants in Loess Hilly Region
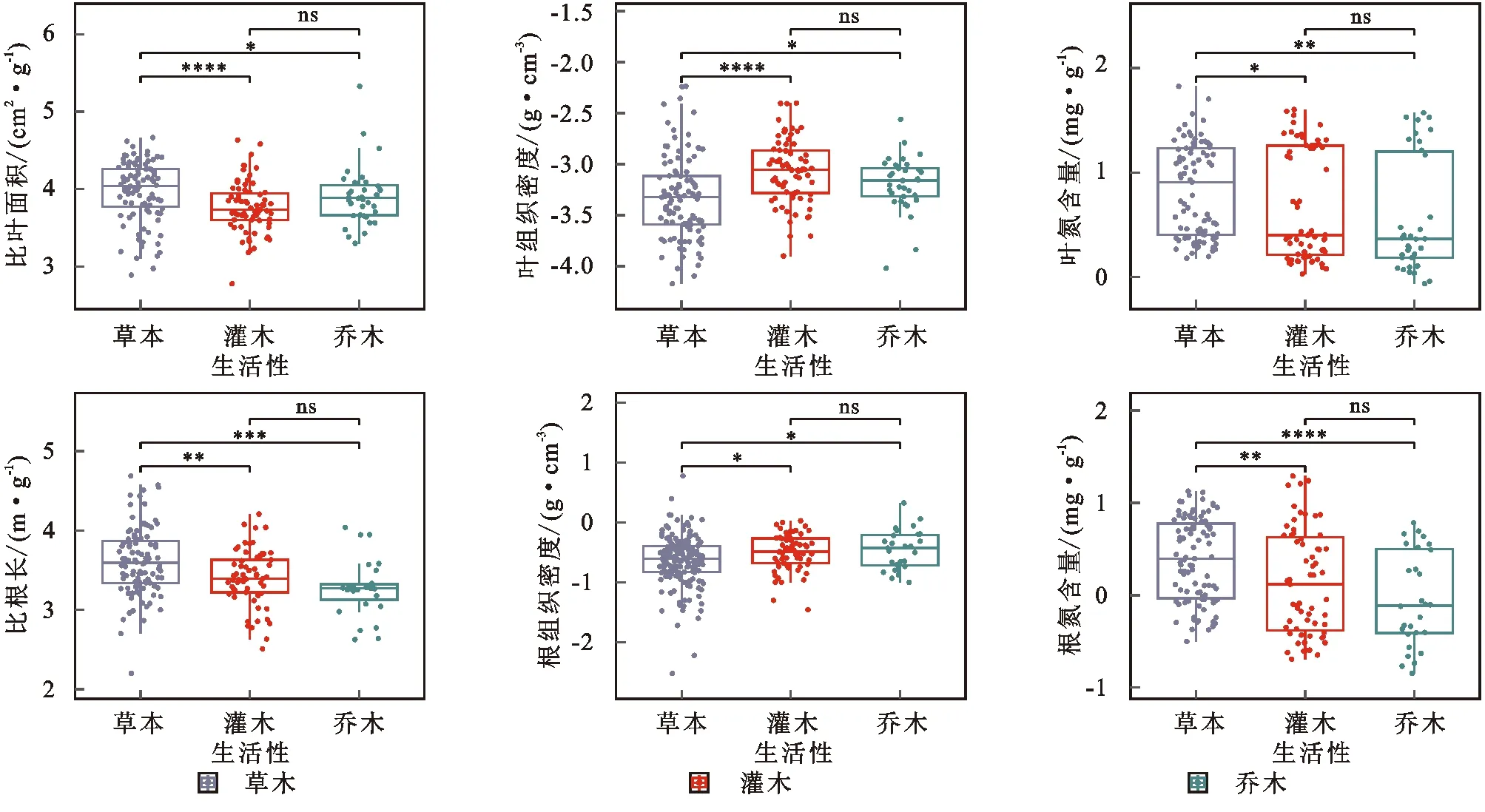
注:*代表0.05显著性水平;**代表0.01显著性水平;***代表0.001显著性水平;****代表0.0001显著性水平。图3 延河流域不同生活型植物功能性状特征Fig. 3 Characteristics functional traits of plants in Loess Hilly Region
ANOVA方差分析结果表明延河流域不同生活型植物的SLA,LTD,SRL,RTD,LNC,RNC均存在显著性差异,其中LTD和RNC在不同生活型间差异最大,LNC在不同生活型间差异最小。SLA,SRL,LNC,RNC方面草本显著高于灌木和乔木且灌木和乔木间差异不显著。LTD,RTD方面草本显著低于灌木和乔木且灌木和乔木间差异不显著。
2.2 植物功能性状关系在不同生活型上的表现
植物对环境变化的适应不仅体现在特定性状的变化上,也可通过不同性状间的协变演化,形成对环境的整体适应策略。因此,分析不同生活型植物功能性状关系,对于理解草本、灌木、乔木环境适应策略具有重要价值。分析结果表明(图4),延河流域草本、灌木、乔木植物的SLA与LTD,SRL与RTD,SRL与RNC表现出显著负相关关系,LTD随SLA变异速率顺序为灌木>草本>乔木,RTD随SRL变异速率顺序为灌木>乔木>草本,RNC随SRL变异速率顺序为乔木>灌木>草本。草本、灌木、乔木的LNC与RNC表现出显著正相关关系,RNC随LNC变异速率顺序为灌木>草本>乔木。草本植物的SLA与LNC,RTD与LNC,RTD与RNC,RTD与LNC表现出显著正相关关系,LTD与LNC,LTD与RNC表现出显著负相关关系。灌木植物的SRL与LNC表现出显著负相关关系。乔木植物的RTD与LNC表现出显著正相关关系,SRL与LNC表现出显著负相关关系。

图4 植物功能性状关系的线性回归分析Fig. 4 Line Regression for Relationships between traits
2.3 不同生活型植物功能性状对环境变化的响应
冗余分析RDA结果显示(图5)第Ⅰ轴主要体现了年降雨量(MAP)、雨季降雨量(Rain789),温度变化(Temsea)、坡度(Slope)、生长季均温(Tem410)对植物功能性状的影响,第Ⅱ轴主要体现了降雨变化(Rainsea)、坡位(Posi)、坡向(Aspe)对植物功能性状的影响。沿着贡献率大的RDA1轴,环境因子年降雨量(MAP)、雨季降雨量(Rain789),坡向(Aspe)与功能性状指标比叶面积(SLA)和比根长(SRL)夹角小于90°,与功能性状指标根氮(RNC)和叶氮(LNC)夹角大于90°,表明功能性状指标比叶面积(SLA)和比根长(SRL)随着环境因子年降雨量(MAP)、雨季降雨量(Rain789)的增加而增加,功能性状指标根氮(RNC)和叶氮(LNC)随着环境因子年降雨量(MAP)、雨季降雨量(Rain789)的增加而减小,而环境因子温度变化(Temsea)、坡度(Slope)、生长季均温(Tem410),海拔(Elevation)与功能性状指标比叶面积(SLA)和比根长(SRL)夹角大于90°,与功能性状指标根氮(RNC)和叶氮(LNC)夹角小于90°,表明功能性状指标根氮(RNC)和叶氮(LNC)随着环境因子温度变化(Temsea)、坡度(Slope)、生长季均温(Tem410),海拔(Elevation)的增加而增加,功能性状指标比叶面积(SLA)和比根长(SRL)随着环境因子温度变化(Temsea)、坡度(Slope)、生长季均温(Tem410),海拔(Elevation)的增加而减小。
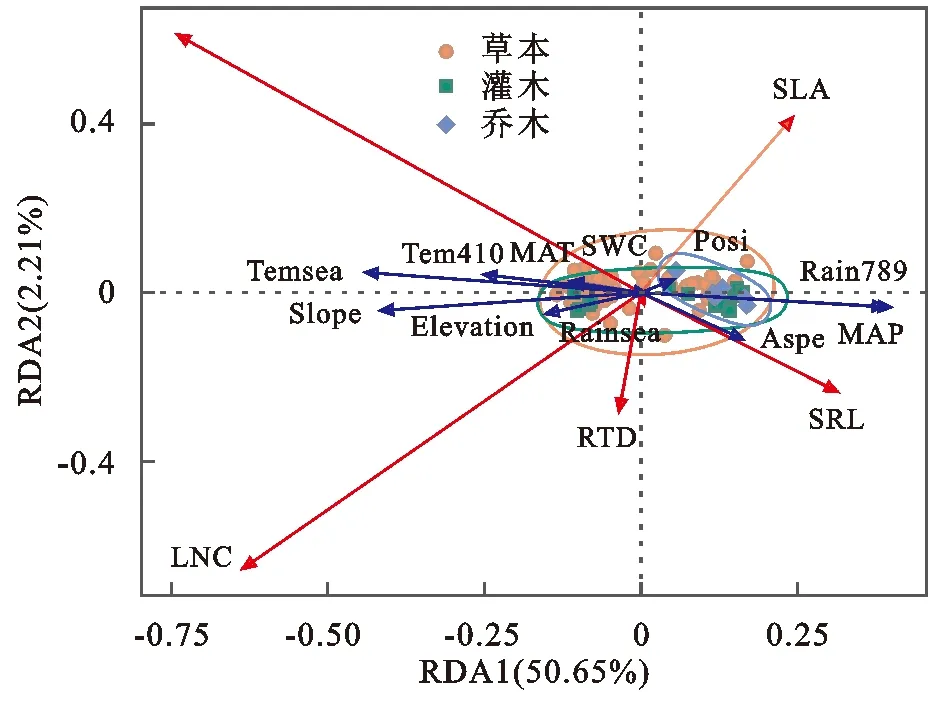
注:groups为生活型组别;Herbs为草本;Shrubs为灌木;Trees为乔木;MAT为年均温;MAP为年降雨量;Temsea为温度变化;Rainsea为降雨量变化;Tem410为生长季均温;Rain789为雨季降雨;SWC为土壤含水量;Elevation为海拔;Slope为坡度;Posi为坡位;Aspe为坡向;SLA为比叶面积;LTD为叶组织密度;SLR为比根长;RTD为根组织密度;LNC为叶氮;RNC为根氮。图5 植物功能性状对环境变化响应的冗余分析(RDA)Fig.5 RDA between plant functional traits and environmental factors
3 讨 论
3.1 延河流域不同生活型植物功能性状特征
延河流域植物SLA表现为草本>乔木>灌木,且草本与乔木、灌木差异显著,这与湖北星斗山[22]的研究结果一致。比叶面积是植物最重要的功能性状指标之一,其定义为叶面积与干重之比,表征植物对光能的捕获能力和潜力。一方面延河流域气候条件与土壤理化性质较一致,降雨少,水分养分不充足,另一方面该区域温度高温差大蒸腾较强,矮小的草本需要更高的资源获取效率。
该区域灌木与乔木植物功能性状差异不显著,这与柴达木盆地[23]的研究结果一致。延河流域乔木植被主要为小乔木,该区域水土流失严重导致深层土壤含水量不高,地表温度较高植物蒸腾作用较强,小乔木相较于高大根深的大乔木更易存活。延河流域草本RTD与灌木、乔木存在显著性差异,这与瑞士高山雪原科尔瓦奇峰[24]的研究结果相同。科尔瓦奇峰的高海拔植物物种间RTD存在显著性差异,甚至一些物种的根强度能达到其他物种的两倍以上。延河流域植物氮含量表现为草本>灌木>乔木,草本LNC,RNC与灌木、乔木存在显著性差异,这与凉水红松林[25]的研究结果一致。氮是植物器官的组成部分且影响植物的生长速度,寿命短、生长快的植物需要更多的氮用于快速生长,草本植物的氮含量高于乔木和灌木。延河流域草本SLA,LNC,SRL,RNC较高,且与灌木、乔木均存在显著性差异,这说明草本的资源获取能力高于灌木和乔木,这是由矮小植株对较廉价组织投资和较快速投资回报的需求决定的。灌木和乔木的LTD,RTD较高,且与草本存在显著性差异,而灌木和乔木两者间各性状均不存在显著性差异,因为高大植株需要相对昂贵的组织投资和相对较慢的投资回报以保持外部形态并维持生理功能。
3.2 延河流域植物生活型植物功能性状关系差异
植物功能性状有力地解释了在漫长历史演化中植物生活型的形成。延河流域乔木植物RTD与LNC表现出显著正相关关系这与昆士兰北部热带雨林[26]的研究结果一致,植株茎密度和根组织密度可以表征植物的抗破坏能力,尤其是乔木植物,并且较高的组织密度与地上器官较高的氮含量特征也与植物从破坏中恢复的能力有关。延河流域草本的灌木的SLA与LNC呈正相关关系,这与厄瓜多尔安第斯山脉南部东坡[27]的研究结果一致。该区域物种SLA与LNC均呈正相关关系,且均与物种每质量光合速率呈正相关关系。延河流域草本植物的SLA与SRL呈正相关关系,SLA与RTD呈负相关关系,这与青藏高原高寒草甸[28]的研究结果一致。在过度放牧的贫瘠草地中,资源获取需求使得草本植物有较高的SRL水平,更高的SLA水平,以及较低的RTD水平,以确保充足的养分吸收满足植物生长需求。
3.3 植物功能性状对环境变化的响应与植物生存策略选择
RDA分析结果表明影响延河流域植物功能性状的主要环境因子依次是年降雨量、雨季降雨量、温度变化、生长季均温、坡度。总的来说,降雨是延河流域植物生存的限制性因子。年降雨量和雨季降雨量较高的区域,生长发育需水多且蒸腾作用耗水多的乔木分布较多,灌木分布也有一定程度的增多。延河流域植物对降雨和温度的响应存在一定程度的负相关关系,因为温度的升高和温差的增大会加剧植物的蒸腾失水和立地环境的干旱胁迫。
根据叶经济谱LES理论、根经济谱RES理论的分类思想和研究标准,由植物功能性状特征定义出一条连续的功能性状组合谱,功能性状谱的一端代表植物的“快速投资-收益型”策略(即获取型),拥有该特征的物种具有相对廉价的组织投资与相对快速的投资回报;而谱的另一端则代表植物的“缓慢投资-收益型”策略(即保守型),拥有该特征的物种具有相对昂贵的组织投资和相对较慢的投资回报[29]。
延河流域草本植物的叶、根组织密度较小,比叶面积、比根长、叶氮含量、根氮含量较大,有着较高的资源获取效率和较低的组织投资,在干旱缺水的延河流域生境下采取了“快速投资-收益”型(即获取型)策略以竞争稀缺的水分。乔木和灌木大多为多年生植物,其叶、根组织密度较大,比叶面积、比根长较小,有着较高的组织投资和较低的资源获取效率,利于资源保护和能量储存,适应环境的策略为“缓慢投资-收益”型(即保守型)。
4 结 论
通过分析延河流域不同生活型植物功能性状特征及其对环境变化的响应发现延河流域不同生活型植物功能性状特征存在显著性差异。水分是延河流域植物生存的限制性因子,高温和温差加剧了蒸腾失水和干旱胁迫。延河流域不同生活型植物采取了不同的资源获取策略和生存策略,乔木和灌木植物采取“缓慢投资-收益”型(即保守型)策略,草本植物采取“快速投资-收益”型(即获取型)策略,这是自然选择和植物进化共同作用的结果。
猜你喜欢
中华建设(2023年10期)2023-10-07
轻音乐(2022年1期)2022-02-11
少儿美术(2020年8期)2020-11-06
艺术品鉴(2019年12期)2020-01-18
音乐天地(音乐创作版)(2019年10期)2020-01-06
现代园艺(2017年13期)2018-01-19
成才之路(2017年21期)2017-07-27
安徽农学通报(2017年9期)2017-05-19
现代农业科技(2016年22期)2017-03-24
学周刊·下旬刊(2015年10期)2015-07-22
