
紫草科2 种短命植物功能性状的差异化协变特征
2023-10-21 05:41金欣悦龚莉王梦亭陶冶周多奇
草业学报 2023年10期
金欣悦,龚莉,王梦亭,陶冶,周多奇*
(1. 安庆师范大学生命科学学院,皖西南生物多样性研究与生态保护安徽省重点实验室,安徽 安庆 246133;2. 荒漠与绿洲生态国家重点实验室,干旱区生态安全与可持续发展重点实验室,中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011;3. 新疆抗逆植物基因资源保育与利用重点实验室,中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011)
植物功能性状(functional trait)被定义为直接影响个体性能(即生长、繁殖和生存)且间接影响其适合度的各类植物属性[1-2],其中形态与生物量等反映个体大小的性状是植物功能性状的重要组成部分,是植物个体适合度的最直接体现。通常,植物株高与植物邻体间竞争光照资源及传播种子的能力相关,即较高的植物更能舒展叶片以获得更多光照面积,同时能促进种子传播以增强植物的繁殖能力[3]。植物生物量则与植物的竞争力和繁殖力密切相关[4]。植物形态特征和生物量分配的变化是植物与环境共同作用的结果,是植物对环境适应能力及生长发育规律的体现[5-6]。在环境影响下,植物进化出了不同的功能群,同一功能群的植物在某一方面(如物候、叶片大小、木质化程度等)表现出趋同适应特征[7],而不同功能群植物则具有明显的趋异适应特征。但即使在同一功能群内,由于物种不同,植物在某些功能性状及其相互关系方面也可能出现较大差异[8-9]。此外,植物各器官的生物量分配比例等性状指标还与植物个体大小有关,即呈现个体大小依赖(body-size dependence)[5,10]。因此,了解植物多性状之间的相互关系及其在不同物种间的差异性是深入揭示植物生态适应策略的重要手段。
异速生长(allometry)反映了植物个体水平上两种性状生长速率不同的现象[11],可用来表示两两性状间的内在关联。异速生长关系是某些生物学特征(如个体形态指标、生物量等)与个体大小(重量)的函数关系,可通过幂函数的形式表达[11-12]。异速生长关系广泛应用于生态学领域,能够揭示植物生物学特征间与尺度无关的内在规律[12-15]。依据代谢理论和分型网络模型,植物性状之间的异速生长指数为1/4 的倍数[14-15]。例如,大量研究证实,植物地上和地下生物量一般呈等速生长关系(即异速生长指数=1)[12,16]。异速生长关系也会随外界条件改变而发生漂变,其中异速生长指数呈现显著差异则认为相关的植物策略发生了本质改变。因此,异速生长关系是揭示植物性状两两关系的重要手段。
植物功能性状间存在着复杂的关系,多种性状相互作用、共同调节着植物功能,使得植物能够应用多种策略对不同环境进行适应并完成生长发育[17-18]。因此,多性状间的相互关系是植物功能性状研究的重要内容。如,Shipley 等[19]通过建立多元模型确定了植物功能性状间相互作用产生的贡献,Díaz 等[20]发现植物性状间是密切联系的,多种性状共同协调植物功能。在植物多种性状中,一些性状群是紧密协调的,且与另一些性状群发挥的功能有所差异,它们共同构成了不同维度的性状集合[21]。解析植物多种性状间的复杂关系多使用主成分分析(principal component analysis,PCA)[22],而当前植物性状网络(plant trait network,PTN)分析已成为前沿技术[23]。PTN 由性状及性状-性状间的关系所构成,用来表达植物性状之间的复杂关系[24]。PTN 不仅是多性状复杂关系网络可视化的工具,而且能够通过网络参数量化多性状间的复杂关系,解析网络结构、整体关联性,并确定核心性状。相关研究已证实PTN 分析的优越性[22-24],其对于揭示植物的性状关联十分必要。
短命植物(ephemerals)是一类特殊的草本植物类群,其利用冬季积雪融水和春季降水而迅速完成生活史(一般不超过2 个月),是我国荒漠植物区系中重要而独特的组成部分[25]。不同种类的短命植物是为逃避炎热夏季而产生趋同适应的结果,除生长期短外,短命植物多具有个体较小、叶片氮含量高、相对生长速率高的特点[26-27]。短命植物生长发育时间与北疆沙漠风沙活动强盛的时期相吻合,这一独特的时空分布规律对于沙面稳定意义重大[28]。目前对短命植物的研究涉及广泛,如短命植物生长对环境的响应[29-30]、短命植物的繁殖策略[31]、异时萌发[32]及耐旱机理[33]等,但不同种短命植物之间是否具有相同或相似的多性状耦合特征目前仍不清楚。
紫草科植物是新疆北部早春短命植物区系中的重要成分[25]。本研究选择广布于古尔班通古特沙漠的紫草科两种不同属的短命植物硬萼软紫草(Arnebia decumbens)和假狼紫草(Nonea caspica)作为试验对象,探究个体形态迥异的两种短命植物是否具有一致的环境适应策略。通过对其传统形态指标及器官生物量异速生长关系、性状协变特征及性状网络结构的对比分析,解析不同种短命植物功能性状及性状间关系的异同,揭示两种植物功能性状间的异速生长关系,阐释二者在相同环境下适应策略的差异性,为深入了解干旱区该类生活型植物的生存策略提供新的科学依据。
1 材料与方法
1.1 研究区概况
古尔班通古特沙漠(44o11′-46o50′ N,84o31′-91o20′ E,4.88×104km2)是我国第二大沙漠,也是我国最大的固定和半固定沙漠。该沙漠年均蒸发量>2000 mm,而年均降水量普遍不超过150 mm,沙漠腹地仅有70~100 mm,且主要集中于冬春季;年均温6~10 ℃,极端高温为40 ℃以上,≥10 ℃的年积温可达3000~3500 ℃,空气相对湿度50%~60%,5-8 月通常<45%[28]。梭梭(Haloxylon ammodendron)和白梭梭(Haloxylon persicum)是荒漠植物群落的主要灌木建群种;该地区具有丰富的短命和类短命植物,是早春荒漠生态系统的重要组成部分[25,34]。本研究中的2 种短命植物在古尔班通古特沙漠广泛分布,通常生于裸沙区或结构不甚致密的生物结皮区(主要为藻结皮和地衣结皮)。
1.2 野外样品采集
所有植物样品于2020 年5 月初短命植物成熟期采集。每个物种设置3 个调查样地,大小为20 m×20 m。在每个样地内按照对角线法选取10 株健康且形状规则的植株,首先利用直尺测定并记录植株高度(height,H,cm)、冠幅长(crown length,L,cm)和宽(crown width,W,cm),统计花的数量(flower number,FN,个);然后采用全株挖掘法(挖掘深度和广度以根系实际分布状况灵活确定)完整地获取待测植物,尤其是根系,随后测量主根长度(root length,RL,cm)。2 种植物最终样本量均为30 株。将每株植物冲洗干净后,分为地上和地下两部分,装在做好标记的信封或布袋内,带回实验室。
1.3 植物功能性状测定
本研究选择易测的形态和生物量指标作为目标功能性状。将野外测定的每株植物的L 和W 求平均值得到平均冠幅直径(crow diameter,D,cm);计算株高与根长(H/RL)和冠幅直径与株高(D/H)的比值,作为2 个重要形态性状指标。植物样品带回实验室后置于75 ℃烘箱内烘干至恒重。将每株植物地上和地下部分单独称重(天平精度为0.0001 g),得到每株地上生物量(aboveground biomass,AGB,g)、地下生物量(belowground biomass,BGB,g)和总生物量(total plant biomass,TB=AGB+BGB,g),并计算根冠比(BGB/AGB,记为R/S)。
1.4 数据处理
利用Levene’s test 对2 种短命植物D、H、RL、FN、H/RL、D/H、AGB、BGB、TB、R/S 共10 个性状进行方差齐性检验,随后进行t-检验以比较种间差异性。R/S、H/RL 及D/H 随个体大小(以总生物量表征)的变化趋势通过幂函数来判断,以揭示植物的比值性状是否随个体大小变化而发生改变,即是否存在个体大小依赖[8,35]。常规数据分析和作图在Excel 2016 中完成,t-检验在SPSS 19.0 中实现。
异速生长关系可以表示为Y=β·Xα,其中,Y是某种生物学特征或者功能,β是标准化常数,X指个体大小,α是异速生长指数[14]。α= 1 为等速关系,即因变量和自变量呈均匀或等比例变化;α≠ 1 为异速关系,即因变量和自变量间为不均匀或不等比例变化。确定异速生长参数时,须将幂函数转换为logY= logβ+α·logX的形式,利用降主轴回归(reduced major axis regression,RMA)方法计算回归模型的指数、95%置信区间(95%CI)和决定系数(R2)[12,36-37]。对数化之后的α即为幂函数对数化之后线性回归的斜率,logβ为线性回归的截距。异速生长指数(α)和截距(logβ)的计算、等速生长检验及α的方差分析均使用SMATR 软件完成。
对所有形态性状进行主成分分析(PCA),以判别性状的协变方向及相互关系。同一轴上性状特征的向量值>0.5 且具有相同符号(正或负),表明这些性状具有较强的协变趋势,而不同符号的性状表示协变趋势相反。如果具有协变的性状较少,则表明性状的分化程度较大;相反,协变的性状数目较多,则可以体现出整合格局的相似性[22,38]。
采用植物性状网络(PTN)拓扑参数解析植物多功能性状之间的复杂关系。为了避免性状之间的虚假相关性,需设定一个阈值(例如P<0.05)来确定性状之间是否真正存在相关性[39]。借助R 语言软件中的“igraph”软件包对 PTN 进行可视化并计算 PTN 的参数,选择节点参数度(degree)、紧密度(closeness)、介数(betweenness)和聚类系数(clustering coefficient)来识别不同性状在PTN 中的拓扑角色,选择边密度(edge density)、直径(diameter)、平均聚类系数(average clustering coefficient)及模块度(modularity)等参数来表征网络总体特征[23-24,40]。
2 结果与分析
2.1 2 种短命植物功能性状基本特征
研究表明,两种植物形态特征的变异系数均表现出中等偏强的变异程度,两者均为地上生物量的变异程度最大,且硬萼软紫草在个体大小的性状方面表现出更强的变异程度(表1)。硬萼软紫草FN 为1~37,平均为8.967;H 为3.4~17.6 cm,平均为10.327 cm;RL 为18.3~62.1 cm,平均为39.01 cm;D 为3.95~22.85 cm,平均为10.945 cm;D/H 为0.842~1.596,平均为1.138;H/RL 为0.148~0.446,平均为0.268。假狼紫草FN 为3~21,平均为10.367;H 为6.4~14.4 cm,平均为9.377 cm;RL 为6.5~58.4 cm,平均为31.497 cm,D 为6.0~24.1 cm,平均为14.260 cm;D/H 为1.088~2.104,平均为1.550;H/RL 为0.188~0.617,平均为0.314。其中,硬萼软紫草的RL 显著大于假狼紫草(P<0.05),而D小于假狼紫草(P<0.01)。
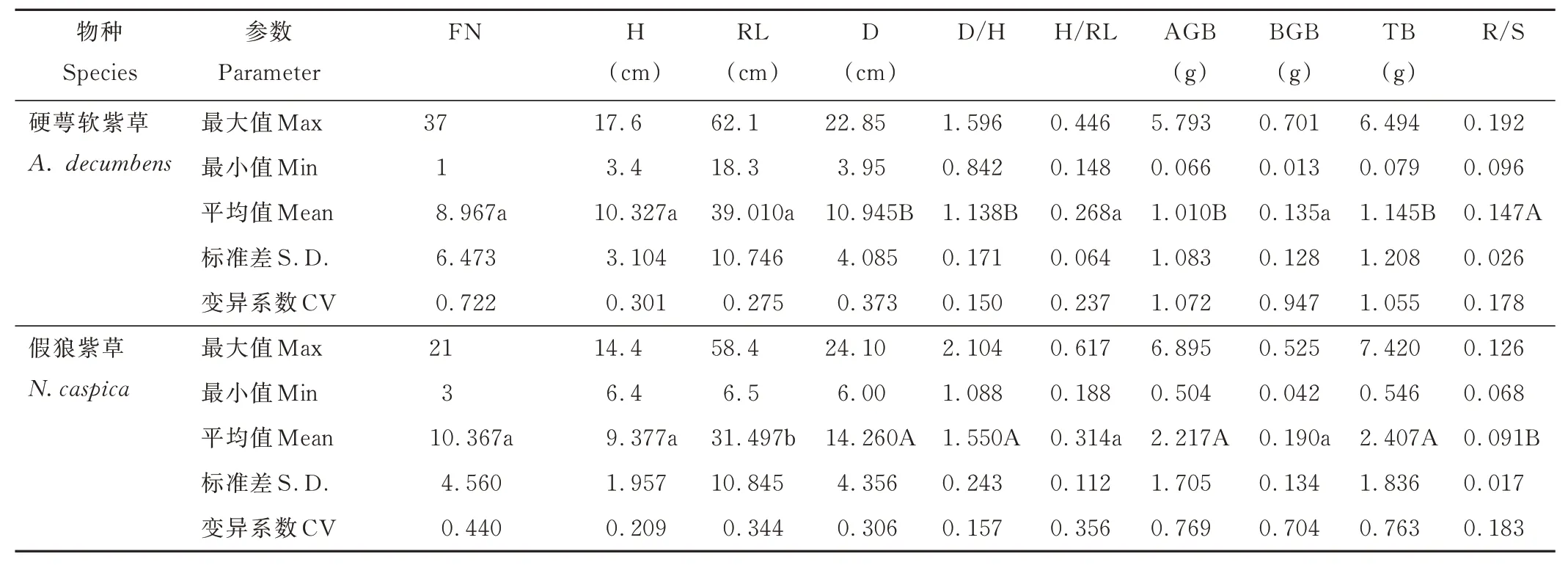
表1 硬萼软紫草和假狼紫草的功能性状对比Table 1 Comparison of functional traits of A. decumbens and N. caspica
硬萼软紫草器官生物量的变异系数为0.947~1.072,属强变异。单株AGB 为0.066~5.793 g,平均为1.010 g;单株BGB 为0.013~0.701 g,平均为0.135 g;TB 为0.079~6.494 g,平均为1.145 g,平均R/S 为0.147。假狼紫草构件生物量的变异系数为0.704~0.769,属中强变异。单株AGB 为0.504~6.895 g,平均为2.217 g;单株BGB 为0.042~0.525 g,平均为0.190 g;TB 为0.546~7.420 g,平均为2.407 g,平均R/S 为0.091。硬萼软紫草的TB 和AGB 显著小于假狼紫草(P<0.01),而R/S 显著大于假狼紫草(P<0.01)。
2.2 2 种短命植物比值性状与个体大小的关系
随着植株个体大小的增加,2 种植物的R/S 均呈显著下降趋势(P<0.01)(图1)。R/S 的下降趋势与植株大小有关,2 种紫草科植物均表现为个体较小时下降快,个体较大时下降速度趋缓,且硬萼软紫草表现更明显。可见,同一种植物在生长的不同阶段,生物量分配比例不同。硬萼软紫草的根冠比总体较假狼紫草高,即硬萼软紫草地下生物量投入比较高。2种植物的H/RL 相似,硬萼软紫草的株高与根长之比略低于假狼紫草,但二者H/RL 随着个体大小的变化未达到显著趋势。2 种植物的D/H 随个体大小的增加均有增加趋势,其中硬萼软紫草达到显著水平(P<0.05),说明植株越大时D 与H 间的差异越明显。硬萼软紫草的D/H 接近1,假狼紫草的D/H 远大于1,表明随个体变大假狼紫草的D 和H 变化不同步,表现为硬萼软紫草的地上形态较瘦高而假狼紫草较宽矮。

图1 硬萼软紫草和假狼紫草比值性状随个体大小的变化趋势Fig. 1 Change trends of ratio traits along with plant size ofA. decumbens and N. caspica
2.3 2 种短命植物功能性状间的异速生长关系
分析表明,硬萼软紫草的形态指标多呈显著的异速生长关系(表2)。等速生长检验表明,仅有RL 与H(α=0.842)、D 与H(α=1.095)、BGB 与FN(α=1.169)及各生物量特征间表现为等速关系,RL 与D(α=0.769)呈α<1(α介于0.326~0.831)的异速关系,其余均为α>1(α介于1.296~3.049)的异速关系,表明硬萼软紫草不同形态性状之间呈现了差异化的内在关联。除FN 与RL(α=1.191)及各生物量特征间表现为等速生长关系,假狼紫草其余形态指标间均为α>1(α介于1.346~3.499)的异速生长关系。两种植物AGB 与BGB 间均为显著的α<1 的异速生长关系,且二者无显著差异,表明二者AGB 分配速率高于BGB。此外,硬萼软紫草的D 与H 呈等速生长关系,而假狼紫草呈α>1 的异速生长关系,表明二者株型随个体大小的变化呈现明显差异;硬萼软紫草的FN 与RL 之间表现为α>1 的异速生长关系,而假狼紫草表现为等比例生长关系。硬萼软紫草生物量与H 的异速生长指数(α=2.566、2.279、2.526)显著低于假狼紫草(α=3.499、3.181、3.464)。硬萼软紫草生物量与RL 的异速生长指数(α=3.049、2.707、3.001)显著高于假狼紫草(α=1.763、1.603、1.745)。可见,2 种短命植物不同功能性状间的异速生长关系也有较大差异。
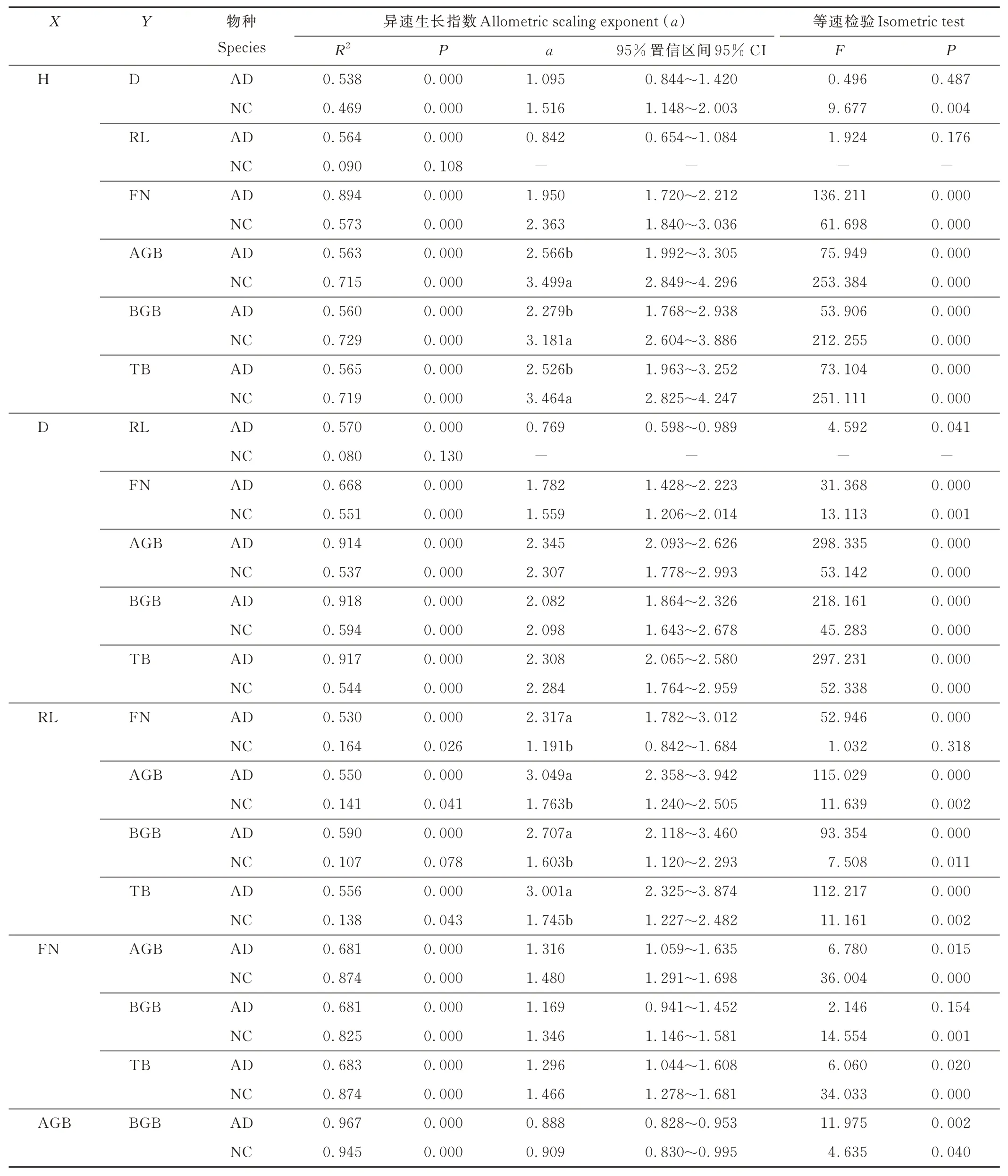
表2 硬萼软紫草和假狼紫草性状间的异速生长指数及等速生长检验Table 2 Allometric scaling exponent and isocratic test among traits of A. decumbens and N. caspica
2.4 2 种短命植物功能性状间的总体关系
PCA 分析表明(图2),硬萼软紫草功能性状第一主成分占总变异量的61.33%,其中D、BGB、AGB、TB、FN、H 及RL 对第一主成分的贡献度较大,这些性状间呈较强的协变关系,但R/S 与上述性状协变趋势相反;第二主成分占总变异量的18.00%,2 个比值性状D/H 与H/RL 的协变方向相反。对假狼紫草而言,其PCA 第一主成分占总变异量的58.27%,D、BGB、AGB、TB、FN、H、D/H 及RL 间具有较为紧密的协变关系,其中R/S 与其他性状协变趋势不同;第二主成分占总变异量的17.82%,其中RL 与H/RL、R/S 的协变趋势相反。上述结果表明,2 种植物功能性状具有不完全一致的协变特征,且具有相同协变趋势的性状群中性状间的关联强度也不同。

图2 硬萼软紫草(左图)和假狼紫草(右图)的主成分分析Fig. 2 Principal component analyses of A. decumbens (left figure) and N. caspica (right figure)
PTN 分析表明,在网络总体参数方面,硬萼软紫草性状网络的边密度较假狼紫草高,而平均路径长度低于假狼紫草,表明硬萼软紫草各性状间的整体协同性较好,有较高的资源利用效率和生产效率(图3 和表3)。在网络节点参数方面,硬萼软紫草性状D 的度和介数都最大,即D 与其他性状的连接最多,因此D 是性状网络中的中心性状也是连接功能模块的桥梁。假狼紫草功能性状中FN 的度和介数均为最大,即FN 是假狼紫草形态性状网络中的中心性状。可见,2 种短命植物性状网络节点参数和网络整体参数均有明显不同,表明2 个同科不同属的短命植物功能性状出现了差异化协变特征。
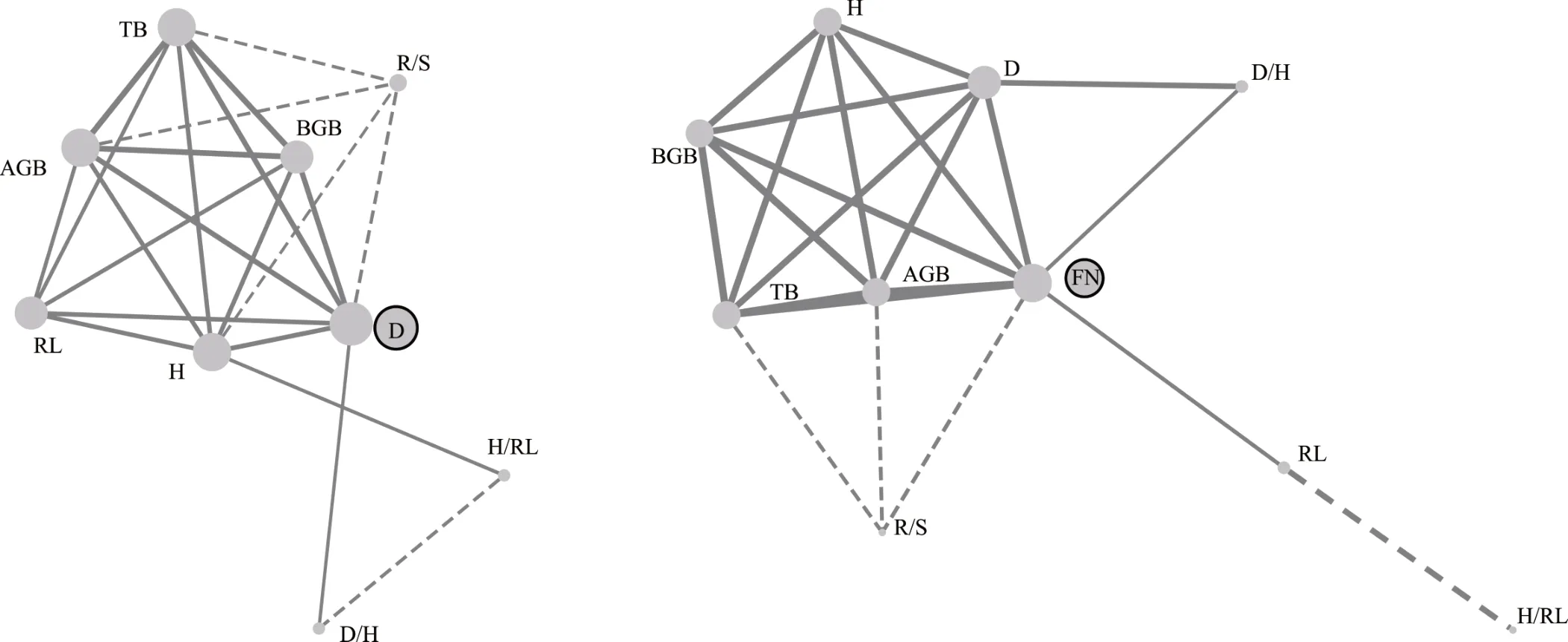
图3 硬萼软紫草(左图)和假狼紫草(右图)的植物性状网络(PTNs)Fig. 3 Plant trait networks (PTNs) of A. decumbens (left figure) and N. caspica (right figure)

表3 硬萼软紫草和假狼紫草的植物性状网络参数Table 3 Parameters of plant trait networks of A. decumbens and N. caspica
3 讨论
3.1 不同短命植物功能性状的差异性
为适应不同的环境条件,植物的功能性状及其组合会表现出差异[41]。而植物在形态上的适应是应对不良环境时最主要的适应机制[42],也是在野外调查中所获取的最直观的信息。植物为适应不同的环境,产生了不同的生活型,不同生活型的区别可通过调整植物构件生物量比例的关系来进行改变。植物为寻找最适合种群扩散和最有利于自身发育的方式改变各构件生物量的分配格局,进而产生了各异的形态特征,形成了丰富的植物系统[43]。在短命植物类群中的不同种间,植物对环境的适应在形态特征方面并没有一致的定律,比较硬萼软紫草和假狼紫草之间的形态特征可以发现,硬萼软紫草和假狼紫草在形态特征上表现出不同的特点,硬萼软紫草的H(株高)、RL(根长)及R/S(根冠比)大于假狼紫草,FN(花的数量)、D(冠幅直径)、AGB(地上生物量)、BGB(地下生物量)及TB(总生物量)均小于假狼紫草。这表明两种植物在相同环境中呈现不同的形态,硬萼软紫草植株更高、RL 更长,而假狼紫草D 更大,地上部分冠幅面积较大且个体质量更重。
生物量分配是植物构件之间资源分配的核心,生物量分配在不同生活型物种之间差异显著[16,43]。根系对于植物吸收土壤中的水分很重要,但高的R/S 并非普遍表现在所有荒漠植物中,生活型或温度对R/S 的影响可能更强于干旱程度对其的影响[44]。在不同生活型和环境的影响下,植物的R/S 呈多样化。植物在地下根的竞争也如地上部般有生态位的不同划分,对于利用浅层土壤的植物来说,它们没有深根系植物如梭梭、疏叶骆驼刺(Alhagi sparsifolia)那样发达的根系[45-46]。就草本而言,李雪华等[43]对科尔沁沙地70 种草本植物生物量分配的研究发现,一年生植物的R/S 均值为0.156(0.027~0.394),稍高于谢然等[35]对4 种一年生荒漠植物的生物量分配研究得出的R/S(0.083~0.158),表明同一生活型在不同环境下的适应性略有差异。同是生长在荒漠环境下的不同生活型植物的R/S 也表现出差异,类短命植物的BGB 比例较大,6 种类短命植物的R/S 为0.355~3.066[8],既高于一年生草本植物也高于本试验的短命植物。生活史短的植物则倾向于将更多的能量投入到地上构件的生长,因短命植物和一年生植物均在当年完成整个生活史,所以不会为保存地下多年生根系而分配更多的生物量到地下部分;相反多年生植物和类短命植物除在地上产生种子外还将较多的生物量分配到地下,实现地下多年生的策略,投入较多的能量用于地下根的生长,具有更大的R/S。这种生物量分配模式的差异是其对环境长期适应而形成不同生活型的结果[29]。类短命植物与短命植物虽然生境相同或相似,但从R/S 来看,二者表现出了不同的响应和适应机制。
本研究中,硬萼软紫草和假狼紫草的R/S 也有所差异,其中硬萼软紫草地下生物量占总生物量的8.8%~16.1%,假狼紫草地下生物量占总生物量的6.4%~11.2%,硬萼软紫草的R/S 为0.147,显著大于假狼紫草的R/S(0.091),这符合之前对短命植物的研究结果[26,30]。假狼紫草R/S 显著低于硬萼软紫草,表示在荒漠极端气候下假狼紫草更注重于地上生长和繁殖,而地下根系投入较少。硬萼软紫草的TB 较假狼紫草轻,而投入到地下部分的资源却比假狼紫草多,这体现2 种短命植物迥异的生物量分配策略。
2 种紫草科短命植物的形态差异较大,但硬萼软紫草和假狼紫草大部分性状间均具有协同生长关系。随植物个体变大,2 种植物R/S 均呈现一定的下降趋势,且在个体较小时下降快,个体较大时下降速度趋缓。2 种短命植物BGB 分配比例随AGB 的增大而减小的结论与郭浩等[47]的研究结果一致,与对准噶尔荒漠6 种短命植物和4种一年生荒漠草本的研究结果也类似[8,35],再次印证短命植物生长过程中存在个体发育漂变[48]。研究结果显示假狼紫草TB 显著高于硬萼软紫草,实际中硬萼软紫草植株较硬挺,假狼紫草植株较柔软易弯,2 种植物的株型差异明显。对于假狼紫草来说,较大的个体质量遇上柔软的茎秆组织,这可能是其长不高的直接原因。由此,2 种同科的短命植物不仅生物量分配存在差异,其表型发育特征也存在截然不同的策略。
3.2 短命植物功能性状间异速生长关系的差异性
植物可通过改变其种子大小和数目、生长率、繁殖时间、竞争能力、胁迫耐受等,采取不同的生活史策略以应对自然选择和环境带来的不同压力,因此植物在响应自然选择压力和自身发育限制时可采取的异速比例模式多种多样[8,49]。诸多研究表明,不同物种间及同物种在不同生长环境中存在一个相对普遍的生物量分配模式[12,14-15,50]。异速生长关系通过数据定量描述了构件性状间的关系,展现出资源分配的模型[15]。同种植物的不同构件性状间具有不同的异速比例关系,性状间呈现差异化的异速生长关系,表明性状之间可能存在不同的协变关系。依据理论预测,全球木本植物地上、地下生物量间均为等比例关系,但其他生物学特征与构件生物量之间多为异速生长关系,其中个体长度与生物量间具有1/4 的异速关系[51]。本试验中硬萼软紫草和假狼紫草的地上、地下生物量间呈现α<1 的异速比例关系,不符合理论预测,但接近在藻结皮、地衣结皮生境中生长的尖喙牻牛儿苗(Erodium oxyrhinchum)的地上、地下生物量间的异速关系[52]。而且,2 种植物R/S 随个体大小的增大均呈下降趋势,这与此2 种植物AGB、BGB 间不呈等速生长相印证。其他形态性状与生物量间呈α>1 的异速生长关系,符合理论预测,表明形态性状生长速率与有机物积累速率存在差异。硬萼软紫草的株高与生物量间呈2/5 的异速关系,高于理论预测的1/4,假狼紫草的株高与生物量间的异速关系则接近1/4。随生物量增加,硬萼软紫草株高增长速度高于其他植物,表现出其对干旱环境独特的适应特点。与假狼紫草相比,随H 增长,硬萼软紫草生物量积累速率较慢,而随根长增长其生物量增加较快,表明2 种植物间性状的异速生长关系呈现差异化。
形态特征间的异速关系在2 种植物间表现出差异,如硬萼软紫草的D 与H 间呈等比例生长关系,而假狼紫草在该组性状关系中呈α>1 的异速比例关系,即假狼紫草D 的增长速率比H 快得多。2 种植物的FN 与H 间均为α>1 的异速生长关系,但随着H 的变化,假狼紫草FN 的生长速率高于硬萼软紫草,可能与两种植物D 的增长速率的差异有关。通常来说,在森林或草原生态系统中植株越高其竞争光照等资源的能力越强,植物的成年株高和地上生物量是可以直观体现植株个体大小的指标,且二者皆与植物的竞争能力相关[3-4]。但假狼紫草的H 显著小于硬萼软紫草,说明假狼紫草在植株高度并不占优势的情况下也能充分保证自身的繁殖能力。这可能与特殊的环境有关,本研究中短命植物分布在沙漠中,与森林生态系统不同,植被较为稀疏,植物冠幅直径大也可增加植物获取太阳辐射的面积。此外,硬萼软紫草的株高高于假狼紫草,但地上生物量显著低于假狼紫草。结合2 种植物的根长、冠幅直径及总生物量的特点,可得出硬萼软紫草全株的体型呈“细长型”,而假狼紫草全株呈“矮胖型”。在实际采样过程中,可见硬萼软紫草整株呈直立生长的状态,其茎等组织较为坚挺,表皮毛更长更坚硬;而假狼紫草整体形态呈趴伏的状态,其表皮毛较柔软,可黏附沙粒结构增加植物所负担的茎叶的质量,这可能与假狼紫草株型呈现低矮化有关。表皮毛的存在也使植物的蒸腾面积缩小以减少水分的蒸发量[53],本研究中2 种植物具有不同形态的表皮毛,也是二者对荒漠环境不同适应策略的体现。
3.3 短命植物功能性状复杂网络关系的差异性
在分析对象的数据种类较多时,通常可以使用主成分分析法将分析对象降维成2 到3 个数据集,再对其进行分析。马克平等[54]对29 个山区植物区系基本特征进行PCA 分析,探讨了植物区系间的关系是否密切,验证了植物区系纬向地带性的规律。在个体水平,有研究表明对植物特征参数进行PCA 降维处理可用于识别和分类不同植物的叶片[55]。对硬萼软紫草和假狼紫草植物的形态性状进行PCA 主成分分析发现,其降维结果较好,前2 轴的贡献率均大于75%。2 种植物性状间整合的协变关系表现出差异,假狼紫草在D/H 与其他性状的协变关系上强于硬萼软紫草,D 与RL 在硬萼软紫草中具有协变关系而在假狼紫草中关系较弱,这些性状间协变关系的差异表明2 种植物的适应策略出现分异。
PTN 是一种将植物性状间复杂的网络关系可视化的手段,性状间的连接称为“边”,边的长短粗细是量化了的相关关系的体现[23-24]。本研究中,D 是硬萼软紫草性状网络中的中心性状兼连接功能模块的桥梁,可能有影响整个表型的中心调控作用。FN 是假狼紫草形态性状网络中的中心性状也是连接功能模块的桥梁。2 种植物的中心性状不同,即2 种植物中对表型起调控作用的性状不同。环境对植物功能模块间的协调关系有较大影响[24],其中网络整体参数边密度高、平均路径长度低表明性状间的协同性好[24]。对不同氮磷添加下短命植物条叶庭荠(Alyssum linifolium)网络性状研究表明,磷添加降低了植物性状网络整体参数,而适量氮添加或氮磷共同添加则有助于提升植物的网络稳定性[39]。本研究中2 种植物的生长环境一致,但性状网络中介数最大的性状却不同,且硬萼软紫草性状网络的边密度较假狼紫草高,平均路径长度较假狼紫草低。这表明,硬萼软紫草性状间表现出更好的协同性,有利于多种功能同时发挥作用;而假狼紫草性状间的独立性较高,这也与2 种植物迥异的株型特征相对应。总体看来,即使同是紫草科的短命植物,其功能性状关系也有不同,在基于形态等易测性状的生态适应策略上存在明显差异。
4 结论
紫草科2 种短命植物的功能性状间具有明显差异,不同性状间的异速生长关系也不完全相同。硬萼软紫草的株高、根长及根冠比大于假狼紫草,而生物量及冠幅直径小于假狼紫草;硬萼软紫草的冠幅直径与株高等速变化,而假狼紫草冠幅直径增长的速率远比株高快。整体来看,2 种植物的株型差异明显,这是植物对相同环境采取不同权衡和生长策略的结果。2 种植物功能性状间的协变关系也不完全一致,但整体上硬萼软紫草的各性状间协同性较高而假狼紫草的各性状间独立性较高。除上述差异,2 种植物也存在一些稳定的功能性状关系,如二者根冠比等比值性状随个体大小的变化趋势一致、地上生物量的分配速率均高于地下生物量。由此可见,紫草科2 种短命植物的功能性状及性状间的关系体现出一定的种间特异性。
猜你喜欢
今日农业(2020年19期)2020-12-14
世界科学技术-中医药现代化(2020年2期)2020-07-25
特别健康(2018年9期)2018-07-17
临床医药文献杂志(电子版)(2017年11期)2017-05-17
中学生物学(2016年8期)2016-01-18
中国科技信息(2015年2期)2015-11-16
中国当代医药(2015年10期)2015-03-01
云南中医学院学报(2014年6期)2014-07-31
植物营养与肥料学报(2014年1期)2014-03-11
中国中医药现代远程教育(2014年22期)2014-03-01
