
具杀虫功能的蛋白类生物材料研究进展
2023-10-19 10:51徐重新金嘉凤刘贤金
农药学学报 2023年5期
徐重新, 金嘉凤, 沈 成, 朱 庆, 张 霄, 刘 媛, 刘贤金
(1.江苏省农业科学院 农产品质量安全与营养研究所/省部共建国家重点实验室培育基地——江苏省食品质量安全重点实验室,南京 210014;2.江苏大学 食品与生物工程学院,江苏 镇江 212013;3.南京农业大学 植物保护学院,南京 210023)
害虫是农作物生产过程中最具危害性的风险因素,对其有效防治直接关乎农作物健康生产乃至提质增产,是农业领域需要持续研究的重大课题。施用农药是害虫防治最直接的有效手段,在世界范围内为保障农业生产发挥了关键作用,其来源广、种类多,现已从传统无机农药、有机合成农药快速发展到了日新月异的生物农药创制和应用阶段,但随着传统农药特别是化学农药长期大范围使用,其本身所普遍具有的毒副作用[1]和由其诱发的靶标害虫抗药性[2-3]以及威胁非靶标生物[4]和生态环境安全[5]等问题日益凸显。害虫绿色防治是农业现代化可持续发展的必然要求,施用农药作为害虫防治必不可少的重要手段,在其创新研发和应用上,由传统的对人类以及其他非靶标生物和生态环境高毒、高残留迈向新型、低毒、低残留乃至无毒无残留的发展道路势不可挡。
生物农药由于生物源的基本属性,其在生产和应用上普遍较传统化学合成农药对人类更为安全,对环境更为友好[6],是目前害虫绿色防治的重要手段。较为典型的生物农药如微生物源性的阿维菌素 (avermectin)[7]、白僵菌素 (beauverin)[8]以及植物源性的苦参碱 (matrine)[9]、蛇床子素(osthole)[10]等对多种特异性靶标害虫具有广谱杀虫功效,均已实现了产品化应用。而具杀虫功能的蛋白类生物活性材料因兼具生物安全性高和可循环量产药剂乃至直接转基因作物改造应用的多重优势,一直是生物农药创新研发和应用的热点,其中最具代表性的如源自苏云金芽孢杆菌 (Bacillus thuringiensis, Bt) 的伴孢晶体蛋白Bt 毒素,相关杀虫药剂和转基因作物在鳞翅目 (Lepidopter)[11]、鞘翅目 (Coleoptera)[12]等多种常见农业害虫防治上发挥了重要作用,在世界范围内产生了巨大的经济价值和社会生态效益[13];此外,凝集素 (lectin) 类杀虫蛋白[14]、多肽类杀虫蛋白如节肢动物毒素[15]、防御素 (defensin)[16]和蛋白酶抑制剂 (protease inhibitor)[17]) 等也都有较为成熟的研究和应用。值得注意的是,作者所在团队近年来创新探索思路,依托抗体“免疫网络学说 (immune network theory)“中Ab2β 类型抗独特型抗体 (anti-idiotype antibody,Anti-Id) 具备模拟抗原结构和功能的特性[18]为理论依据,结合噬菌体展示基因工程抗体库及抗原特异性基因工程抗体高通量富集与筛选鉴定技术,通过设计Bt Cry 毒素抗体为靶点包被抗原,初步实现了具备模拟Bt Cry 毒素杀虫功能的全新效应物杀虫抗体靶向创制[19],可为蛋白类生物杀虫材料功能模拟物靶向创制提供全新思路和实例参考。
本文在系统梳理具杀虫功能的蛋白类生物材料研究和应用现状的基础上,重点结合作者所在团队在杀虫抗体靶向创制与创新应用模式的最新研究成果和积累经验,进一步探讨具杀虫功能的蛋白类生物材料在未来创新研发与应用方面的潜在可行路径,为农业害虫绿色防治相关的蛋白类生物材料研究提供最新文献资料和潜在启发思路。
1 具杀虫功能的蛋白类生物材料主要来源
生物源性是具杀虫功能的蛋白类生物材料的最鲜明特质,自然界中微生物、植物和低等级动物等是原始蛋白类杀虫材料的最主要来源。按材料特性,可大致分为Bt 毒素类杀虫蛋白、凝集素类杀虫蛋白以及包括动物毒素、防御素、蛋白酶抑制剂在内的小分子多肽类杀虫蛋白,此外还有少数目前尚无法归类的其他杀虫蛋白;相关材料主要类型及典型代表见表1。微生物源性的苏云金芽孢杆菌Bt 毒素类杀虫蛋白,是当前生物农药领域最具代表性的蛋白类生物杀虫材料,国际Bt 毒素权威统计数据库(http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/) 显示,截至2023 年4月,已获命名的Bt 毒素家族杀虫蛋白亚型种类多达1005 个,包括Bt Cry1~78 毒素家族亚型818个、Bt Cyt1~3 毒素家族亚型40 个、Bt Vip1~4 毒素家族亚型147 个。凝集素类杀虫蛋白主要集中在植物凝集素上,研究较多的如刀豆Canavalia virosa凝集素、大蒜Allium sativum凝集素及雪花莲Galanthus nivalis凝集素等,此外在个别细菌、真菌蘑菇以及海洋软体动物中也发现有相应凝集素类杀虫蛋白。多肽类杀虫蛋白材料分子量大多在80 个氨基酸以内,其种类丰富多样,来源更是涵盖了动植物和微生物[15],其中陆地节肢动物毒素 (如蜘蛛毒素、蝎毒素、蜈蚣毒素、蚁类毒素、蜂类毒素) 和低等级海洋动物毒素 (如海葵毒素、水母毒素、芋螺毒素) 是多肽类杀虫蛋白材料的最主要来源;此外,植物源性的防御素和蛋白酶抑制剂也是多肽类杀虫蛋白材料的重要来源。
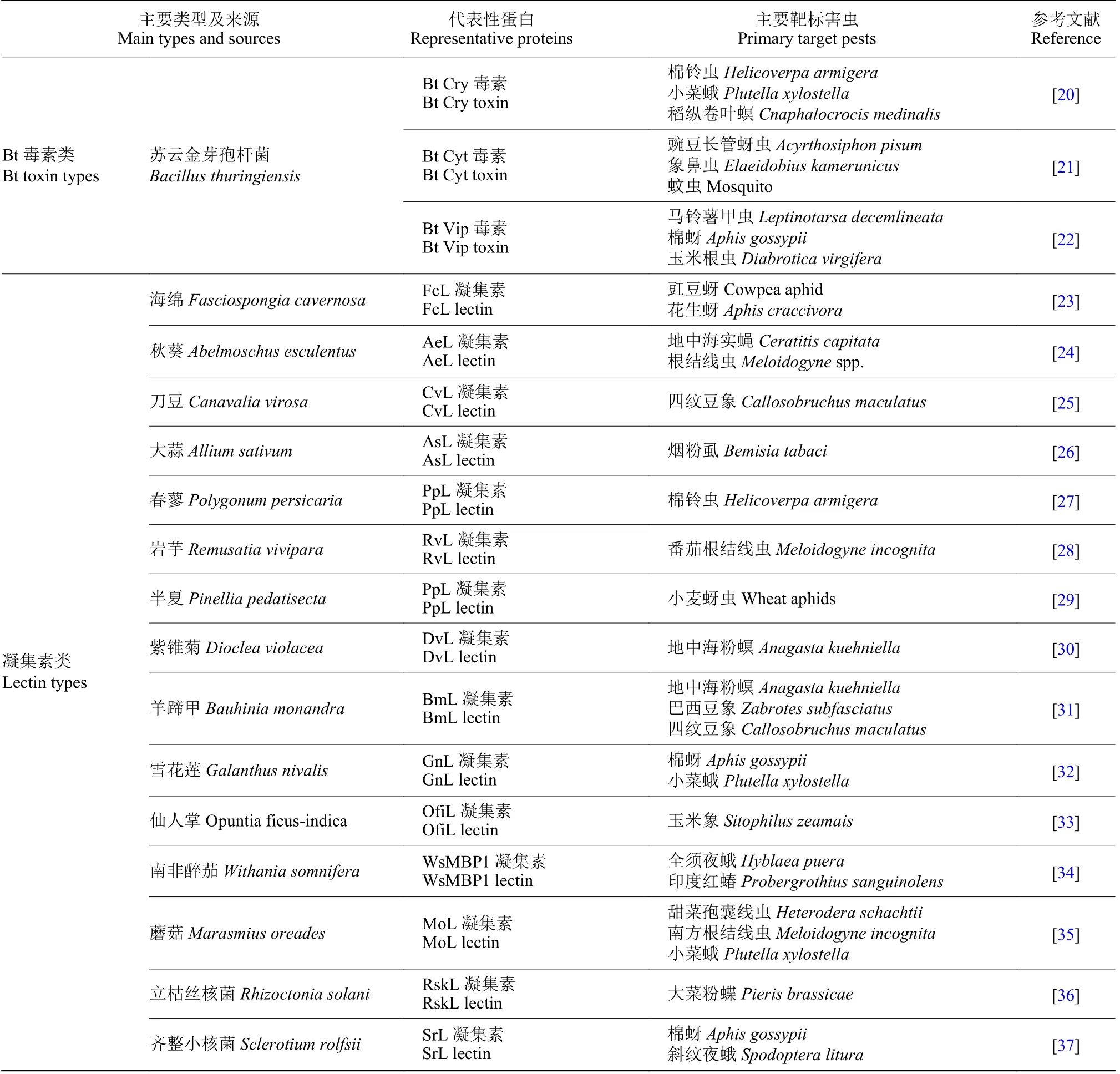
表1 具杀虫功能的蛋白类生物材料主要类型及典型代表Table 1 The main types and typical representative of protein-type biological materials with insecticidal function
2 具杀虫功能的蛋白类生物材料作用机制
Bt 毒素作为最具代表性的蛋白类杀虫生物材料,其对靶标害虫的作用机制是竞先探究的热点,目前尤以Bt Cry 毒素家族杀虫蛋白的作用机制研究得相对较为透彻,相关机理认识也较为清晰。主流观点认为,靶标害虫摄入Bt Cry 毒素后,中肠分泌的蛋白酶将其水解形成分子质量约为60 kDa 大小的活化态,进而与中肠刷状缘膜囊(brush border membrane vesicles,BBMV) 上的钙黏蛋白 (cadherin,CAD) 受体靶向特异性结合,诱发活化态毒素蛋白进一步水解形成单体寡聚化,接着与糖基磷脂酰肌醇锚定蛋白连接的细胞膜上的氨肽酶-N、碱性磷酸酶等关键受体强力结合,从而牵引毒素蛋白穿插到细胞膜中造成穿孔,导致肠道细胞裂解引发害虫死亡[78];也有部分观点认为,Bt Cry 毒素进入靶标害虫中肠与BBMV 上的CAD 受体结合后,刺激肠道细胞膜蛋白G 蛋白偶联受体系统诱发系列级联生化反应,引起环磷酸腺苷过表达,从而激化蛋白激酶A 信号通路,导致肠道细胞裂解引发害虫死亡[79]。
凝集素类杀虫蛋白作为一类能与糖及糖类复合物特异性非共价可逆结合的生物大分子材料[14],其对靶标害虫的作用机制往往与其本身所具备的强力结合糖及糖类复合物大分子的特性密切相关,特别是与靶标害虫肠道功能性糖类蛋白互作,抑制相关酶活性 (如α-淀粉酶、β-葡萄糖苷酶等),干扰取食和营养吸收,从而影响虫体生长发育乃至死亡[80]。如立枯丝核菌Rhizoctonia solaniKühn 凝集素对大菜粉蝶Pieris brassicae[36]、羊蹄甲Bauhinia monandra凝集素对四纹豆象Callosobruchus maculatus[31]、附生蕨Microgramma vacciniifolia凝集素对对玉米象Sitophilus zeamais[81]、辣木Moringa oleifera凝集素对地中海粉螟Anagasta kuehniella[82]、南非醉茄Withania somnifera凝集素对全须夜蛾Hyblaea puera[34]等相关研究,均表明供试的凝集素类杀虫蛋白对相应靶标害虫肠道功能造成了损伤,尤其是抑制了消化酶的活性,引起肠道消化吸收功能受阻,导致虫体生长发育迟缓乃至死亡。
多肽类杀虫生物材料对靶标害虫的作用机制则更为复杂多样,其中动物毒素类杀虫多肽的作用机制主要与其普遍具有靶向干扰虫体细胞钠、钙离子通道或强力阻遏如烟碱乙酰胆碱受体 (nicotinic acetylcholine receptors,nAChRs) 等功能性门控通道蛋白的特性有关[15],主要症状通常表现为兴奋性麻痹瘫痪直至死亡。如蜘蛛毒素 μ-SPRTX-Hv2[83]、蝎毒素AaIT[41]、海葵毒素Av3[84]和缎带虫Nemertideα-1 毒素[50]等均是通过干扰钠离子通道的方式分别对美洲大蠊Periplaneta americana、棉铃虫Helicoverpa armigera、果蝇Drosophila 及甘蓝夜蛾Mamestra brassicae等靶标害虫产生抑制或致死活性,蜘蛛毒素OxyTx[38]和HWTX-V[85]杀虫多肽是通过干扰钙离子通道的方式分别对草地贪夜蛾Spodoptera frugiperda和美洲大蠊等靶标害虫产生抑制或致死活性,而蜘蛛毒素GS-ω/κ-HxTx-Hv1[86]和芋螺毒素ImI [87]等杀虫多肽则是通过阻遏nAChRs 活性的方式分别对蚜虫 (aphid) 和黄粉虫Tenebrio molitor等靶标害虫造成不可逆伤害乃至死亡。防御素类杀虫多肽的活性机制一般认为与其普遍富含半胱氨酸且通过二硫键连接形成的稳定结构有关[88],这类特殊结构能特异性破坏靶标害虫肠道功能,阻碍取食行为,影响消化吸收和生长发育。如豌豆PA1b 防御素[89]特异性阻碍靶标害虫肠道V-ATP 酶活性、绿豆VrD1 防御素[51]强力抑制靶标害虫肠道α-淀粉酶活性、百花蛇舌草Kalata 防御素[56]干扰靶标害虫肠道上皮细胞功能等,均是破坏相应害虫肠道功能引发虫体生长发育异常乃至死亡。蛋白酶抑制剂类杀虫多肽对靶标害虫的活性机制与防御素具有一定相似之处,其主要也是通过抑制靶标害虫消化道特殊功能蛋白酶活性,造成肠道细胞受损或消化吸收受阻,从而扰乱虫体生长发育乃至致死。如巴西木蝶豆CFPI[62]、印度黑豆VmPI [63] 及苦荞麦TBTI-II[67]等蛋白酶抑制剂均抑制中肠胰蛋白酶活性、木耳Clitocypin[68]蛋白酶抑制剂抑制肠道半胱氨酸蛋白酶活性,泡盛曲霉AaPI[69]蛋白酶抑制剂抑制消化道α-糖苷酶活性等,均造成相应靶标害虫消化功能受阻,影响虫体生长发育乃至死亡。
3 具杀虫功能的蛋白类生物材料应用现状
制剂和转基因抗虫作物改造是具杀虫功能的蛋白类生物材料在靶标害虫防治应用中的两种最主要形式。以制剂形式应用方面,Bt 毒素无疑是世界范围内生产和使用最广泛的蛋白类杀虫材料,根据农业农村部农药检定所权威统计信息 (中国农药信息数据库http://www.chinapesticide.org.cn/),截至2023 年4 月,我国登记的有效成分含Bt 毒素的生物农药产品已超过240 余种;多肽类杀虫材料也有一些成熟的商品化制剂产品在应用,较为典型的如蜘蛛毒素源性的Spear®杀虫剂[90]和植物防御素源性的Sero-X®杀虫剂[91]。此外在美国、澳大利亚均有类似多肽类杀虫材料作为杀虫剂进行农药产品登记[15];而凝集素类和其他类型的杀虫蛋白材料目前尚无相关制剂产品和农药登记。以转基因抗虫作物形式应用方面,Bt 毒素同样是世界范围研究最深入、应用最成熟的杀虫蛋白材料,国际农业生物技术应用服务组织 (International Service for Acquisition of Agri-Biotech Applications,ISAAA) 统计数据显示,全球每年种植的涵盖棉花、水稻、玉米、小麦、大豆、马铃薯、油菜、甘蓝、甜菜及烟草等常见经济作物在内的转Bt 毒素基因的抗虫作物已超过1 亿公顷,产业增值超过190 亿美元[13]。凝集素类和多肽类杀虫蛋白材料在转基因抗虫作物上的应用,目前多数处于室内探索的初级研发阶段,成熟的具备可商品化推广应用的转基因抗虫作物品种较少。含转凝集素类杀虫蛋白基因的抗虫作物研究现已涵盖棉花[37]、小麦[92]、水稻[93]、烟草[94]和芥菜[61]等类型,相应转基因作物分别对斜纹夜蛾Spodoptera litura、小麦蚜虫、黄茎螟(yellow stem borer)、烟草粉蚧Phenococcus solenopsis和芥菜蚜虫等靶标害虫具有良好防治效果;而含转多肽类杀虫材料基因的抗虫作物研究也涉及了水稻[95]、烟草[94]、棉花[96]、豌豆[16]、马铃薯[60]、芥菜[61]等类型,相应转基因作物分别对褐飞虱Nilaparvata lugens、烟草粉蚧、棉贪夜蛾Spodoptera littoralis、豌豆蚜虫、马铃薯甲虫Leptinotarsa decemlineata和芥菜蚜虫等靶标害虫具有良好防治效果。
4 具杀虫功能的蛋白类生物材料潜在问题
靶标害虫抗药性和非靶标生物的毒副作用几乎是所有农药在应用过程中都不可避免会面临的两大潜在共性问题,前者是农药长期或不合理使用所诱导靶标害虫抗药性进化产生的,而后者则是由农药本身药理性质所决定的。Bt 毒素作为最具代表性的蛋白类杀虫材料,其应用时间长、使用面广,因此暴露的靶标害虫抗药性和非靶标生物的毒副作用问题也相对更突出。自从小菜蛾成为野外第一个被发现对Bt Cry 毒素产生抗药性的害虫以来[97],目前至少包括小菜蛾、棉铃虫、棉红铃虫Pectinophora gossypiella、草地贪夜蛾Spodoptera frugiperda、灰翅铃夜蛾Helicoverpa punctigera、粉纹夜蛾Trichoplusia ni、玉米螟Ostrinia furnacalis和山杨叶甲Chrysomela tremulae在内的7 种鳞翅目和1 种鞘翅目害虫已对转Bt Cry1Aa/Ab/1Ac/Ah/1B/Cry1Ca/1F/1J/2Ab/2Ae/3Aa 等类型毒素基因作物中的一种或多种产生了抗性[98],形势异常严峻。合理轮换药剂是目前包括对Bt 毒素产生抗性在内的抗性治理最为直接有效的方式,如茚虫威 (indoxacarb) 与Bt 毒素尚未形成交互抗性,其对Bt 毒素产生抗性的害虫仍然保持良好的敏感性,因此在相关害虫抗性治理上效果显著[99]。此外,借鉴农药复配策略,对Bt 毒素进行异源融合改造也是治理其抗性的潜在手段,如将Bt Cry1Ac 毒素的N 末端与蓖麻凝集素(ricin B-chain) 基因融合,形成的全新杀虫复合物的毒力较Bt Cry1Ac 毒素提升了4 倍,具备治理相关抗性害虫的应用潜力[100]。而对大量非靶标生物的毒副作用风险监测表明,尽管Bt 毒素的生物安全性普遍很高,但仍然有部分Bt 毒素类型对包括家蚕Bombyx mori[101]、二星瓢虫Adalia bipunctata[102]、秀丽隐杆线虫Caenorhabditis elegans[103]以及土壤微生物[104]在内的特定非靶标生物存在直接或间接安全隐患。尽管对凝集素类和多肽类杀虫蛋白材料的研究报道也较多,但总体仍处于室内探索的起步阶段,由于实际应用较少,相关的靶标害虫抗药性问题目前均未暴露出来。已有的非靶标生物毒副作用风险监测表明,部分凝集素类杀虫蛋白在一定程度上存在风险隐患,如刀豆凝集素Con A 能显著减少其在转基因作物生境中的非靶标原生动物种群数量[105]、雪花莲凝集素GNA 对蜜蜂幼虫[106]和绿草蛉Chrysoperla carnea成虫生殖系统[107]均能造成危害,辣木凝集素WSMoL 对斑马鱼胚胎和幼体具有致死和亚致死毒性[108]等,而多肽类杀虫蛋白材料及其转基因作物目前尚未发现类似风险。
5 具杀虫功能的蛋白类生物材料创新研发与应用动向
自然界生物体是挖掘具杀虫功能的蛋白类生物材料的原始资源库,但随着世界范围内不断大规模探寻,近年来直接从生物体中获取相关新材料显得尤为吃力。依托现有材料对其进行活性改造或功能修饰,是具杀虫功能的蛋白类生物材料创新研发的持久性热点,特别是围绕Bt 毒素杀虫蛋白的活性改造和功能修饰上,相关研究较多,也最具参考借鉴价值。如Gómez 等[109]在Bt Cry1Ab毒素Dmain III β-22 引入定点突变,得到的突变体对草地贪夜蛾的毒性显著增强;Jiang 等[110]在Bt Vip3Aa Dmain II 和Dmain III 之间引入点突变,得到的突变体对草地贪夜蛾和棉铃虫的毒性显著增强;Liu 等[111]将Bt Cry1Ab 毒素Dmian III 替换为Bt Cry1Gc 毒素的Dmian III,以该结构域替换体获得的转基因水稻和转基因玉米对二化螟和玉米螟的抗性水平显著提升;而Chen 等[112]更是将Cry1Ab-前导肽、Cry1Ah-Domain I、Cry1Ac Domain II 和Cry1Ca-Domain III 拟合形成全新的结构域重组体Cry1A.2 毒素,其较4 种供体母本毒素的杀虫谱更广,极大提升了应用价值。值得关注的是,近年来以抗体“免疫网络学说”中Ab2β 类型Anti-Id 具有抗原“内影像”效应为理论依据[18],在可模拟抗原部分结构乃至生物功能的效应物抗体材料创制上有了一些初步成功的探索[113],有望成为具杀虫功能的蛋白类生物材料功能替代效应物创新研发的捷径。特别是作者所在团队依托这种理论,通过设计Bt Cry 毒素抗体为包被抗原,通过“投入-结合-洗脱-扩增”等富集淘筛过程,并结合抗原特异性酶联免疫吸附测定(ELISA)、靶标害虫潜在关键受体竞争抑制以及生物测定验证等策略,现已从噬菌体展示抗体库中成功获得出了一系列具备模拟Bt Cry 1Aa/1Ab/1Ac/1B/1C/1F/2Aa 等相应毒素杀虫功能的Ab2β Anti-Id 及对应基因材料[19],其中活性最强的模拟物杀虫抗体对小菜蛾的致死率达到原Bt Cry 毒素的77%,在国内外率先实现了蛋白类杀虫生物材料模拟物杀虫抗体的靶向设计研发,具有重要的理论实践意义和技术参考价值。
借鉴传统化学农药联合复配增效模式,探寻将不同类型的杀虫蛋白基因进行融合,再以表达制剂或转基因抗虫作物改造的方式施用,是不断提升现有杀虫材料对靶标害虫防治效率乃至扩大杀虫谱的最直接有效手段,目前尤以Bt 毒素不同亚型基因融合或与其他类型杀虫基因融合的创新应用居多,而除Bt 毒素以外的其他类型杀虫蛋白基因之间融合的创新应用也有一些报道,相关典型实例及效果见表2。此外,以植物内生菌 (endophyte)为生防载体菌搭载外源杀虫蛋白定殖宿主作物防治靶标害虫,有望实现蛋白类杀虫材料在害虫防治上突破传统制剂或转基因方式迈向更为绿色的创新应用模式,目前已有初步探索成功的报道,如Zhang 等[114]以植物内生细菌Enterobacter cloacae SJ-10 搭载抗白背飞虱的半夏凝集素蛋白定殖水稻,Qi 等[115]以植物内生真菌Chaetomium globosum YY-11 搭载抗蚜虫的半夏凝集素蛋白定殖油菜,均显示构建的重组生防工程菌可以协同宿主作物防治相应靶标害虫。

表2 具杀虫功能的蛋白类生物材料融合创新应用典型实例Table 2 Typical examples of innovative applications for the fusion of protein-based biological materials with insecticidal functions
6 结语与展望
害虫绿色防治是农业绿色发展的必然要求,具杀虫功能的蛋白类生物材料因其生物源的特质属性,对人类以及其他非靶标生物的安全性普遍较高,在害虫绿色防治中扮演着重要角色,相关研究的科学意义和潜在价值不言而喻,是生物农药创新研发和应用的重点。
在材料创新研发上,自然界中生物体显然是具杀虫功能的蛋白类生物材料的最原始创新来源,特别是在有毒生物如陆生节肢动物、海洋生物、真菌蘑菇等毒素中蕴藏尤为丰富,但随着长期探寻,近年来从生物体中直接获取新型杀虫材料显得愈发困难,特别是如Bt 毒素的高活性高价值杀虫材料更是凤毛麟角。依托现有材料为模板研发活性更高或杀虫谱更广的突变体、不同类型材料之间的重组体或杂合体,是目前杀虫蛋白材料创新的重要手段,但实现这种创新改造的前提是供试模板本身结构和功能乃至对靶标害虫的活性机制要相对较为清晰,否则盲目性较大,很难达到预期效果,这也是相关研究几乎都是以Bt 毒素为模板的重要因素。与改造路径相比,依托抗体“网络免疫学说”中Ab2β Anti-Id 具备模拟抗原结构乃至生物学功能的特性为理论,指导现有材料功能模拟物创制路径,在适用性上则更广,理论上只要有相应材料的抗体并以其为免疫抗原或包被抗原,通过动物免疫或抗体库筛选并结合Ab2β Anti-Id 分型和相应靶标害虫生测验证,就能获得其功能模拟物,探索空间巨大。目前这方面研究虽已起步,但诸如包被抗体设计以及靶标模拟物筛选效率、模拟功效等一系列问题还有待完善和解决[19],前景可期,但也任重道远。
在材料创新应用上,借鉴化学农药联合复配模式,将不同药理药效的杀虫蛋白基因融合,再通过表达制剂或转基因抗虫作物方式提升对共同靶标害虫的防治效果或扩大杀虫谱,这种创新应用实效性较强,具有极大挖掘潜力。此外,借助植物内生菌搭载杀虫蛋白直接定殖宿主作物协同防治靶标害虫的创新应用模式,有望避开蛋白类制剂型生物农药表达制备和维持活性稳定上的困境[127],也能跳过转基因作物构建过程的复杂技术流程[128]且在不改变宿主作物其他性状的前提下达到与转基因作物类似的抗虫功效,是蛋白类杀虫材料突破传统制剂或转基因应用模式的潜在可行路径,目前相关研究尚处于起步阶段,未来可期。不过该创新应用模式受植物内生菌和其搭载的杀虫蛋白活性双重制约:一方面,植物内生菌作为健康植株体内的共生菌,正常情况下,其在植株体内定殖量必然不会太多,因此需要解决可能由其在宿主作物中的定殖丰度低所导致搭载目标杀虫蛋白总表达和蓄积量不足以防治靶标害虫的问题,如通过循环接种驯化提升其对宿主作物的定殖能力和蓄积丰度;另一方面,也需不断改进供试杀虫蛋白杀虫活性和优化与植物内生菌相适应的表达载体,从而提升单位植物内生菌搭载杀虫蛋白重组生防工程菌的毒力,是弥补植物内生菌对宿主作物定殖能力或蓄积丰度不足的重要手段。
猜你喜欢
军民两用技术与产品(2021年10期)2021-03-16
世界农药(2019年3期)2019-09-10
今日农业(2019年12期)2019-08-15
中国中医药信息杂志(2016年7期)2016-12-01
光学精密工程(2016年4期)2016-11-07
中华老年多器官疾病杂志(2016年7期)2016-04-28
肿瘤影像学(2015年3期)2015-12-09
川北医学院学报(2015年5期)2015-12-05
国际心血管病杂志(2015年5期)2015-02-27
中国质量与标准导报(2014年4期)2014-03-11
