
不同抗性甘蓝品种对黑腐病的生理响应
2023-10-09 11:17:52张鑫鑫
种子科技 2023年16期
张鑫鑫
(山西运城农业职业技术学院,山西 运城 044000)
结球甘蓝简称甘蓝,是一种十字花科芸薹属蔬菜作物,栽培历史悠久,在世界各地广泛栽培[1]。随着甘蓝在世界各地的普遍推广,复种指数居高以及重茬严重,各种病害相继出现,由野油菜黄单胞菌引起的黑腐病为其中一种主要病害[2]。
甘蓝黑腐病常发生于温暖、潮湿的气候条件下[3]。近年来,甘蓝黑腐病在我国蔬菜生产区普遍发生,病征表现为:初期,叶片边缘出现黄褐色V 形病斑。随着病原菌在植株体内蔓延,出现维管束变黑、植株枯死的情况,且常伴随软腐病的发生,严重影响甘蓝的产量和质量[4-9]。
国内外许多学者研究发现,小麦、花椰菜、大豆、烟草、花生、菜豆、水稻、黄瓜、大白菜等多种植物感染病原菌后,叶片和根系的生理生化活动会发生一些独特的变化,这些变化与植物的抗病性相关[10-12]。但是,目前关于结球甘蓝感染黑腐病菌后叶片及根系发生的生理生化反应的研究报道较少。因此,本研究选用5 个抗性不同的甘蓝材料,分别比较结球甘蓝在感染黑腐病菌后根系以及叶片生理生化指标的变化与品种抗病性的关系,旨在为深入研究结球甘蓝对黑腐病的抗病机理和加快抗病品种选育工作提供依据。
1 材料与方法
1.1 材料
1.1.1 供试菌株
菌株由西北农林科技大学园艺学院甘蓝抗病育种研究室提供,编号YU,致病力较强[13],该菌株分离自陕西省榆林市田间自然发病的甘蓝病叶。
1.1.2 供试甘蓝材料
选择田间抗甘蓝黑腐病差异显著的5 个品种:高抗材料QK2502(HR)、抗病材料QG70(R)、中抗材料LQ66(MR)、感病材料QG50(S)、高感材料B02(HS)杂种一代种子,均由西北农林科技大学园艺学院甘蓝抗病育种研究室提供。
1.2 试验方法
1.2.1 幼苗培养
试验在西北农林科技大学科研温室进行。挑选供试甘蓝种子,选择籽粒饱满、整齐一致的种子,先在75%酒精中处理1 min,再用灭菌蒸馏水冲洗2~3 次,在25 ℃催芽箱中催芽2 d,选取发芽一致的种子播种于50 孔穴盘灭菌基质中,播种后的育苗盘置于温室中正常管理。
1.2.2 黑腐病菌悬浮液制备
菌株YU 在接种前18 h 转接到牛肉膏蛋白胨液体培养基中,摇床28 ℃、190 r/min 培养18 h,再用无菌水调节菌液浓度至1×108cfu/mL。
1.2.3 接种
待供试材料长至3 叶1 心期进行接种。接种前24 h 将材料浇透并覆盖塑料薄膜保湿,第2 天用小型喷雾器将菌液均匀喷洒到植株叶片上,以叶片无液滴滴落为度。喷施无菌水作为空白对照。每个处理20 株苗,设置3 次重复。接种后继续用塑料薄膜保湿36 h,后揭去塑料薄膜,将幼苗放在温度26、19 ℃,湿度90%条件下正常管理,光照时长14 h[14]。
1.2.4 取样
从接种当天起,分别于0、1、3、5、7 d 的固定时间对甘蓝幼苗叶片进行取样,取样后将样品保存于-80 ℃冰箱中备用,其中,地下部相关生理指标的测定取样时间是1、7 d。
1.2.5 测定项目和采用方法
甘蓝材料根系相关指标:根长,采用直接测量法;根系鲜重,采用直接称量法;根系活力,采用TTC 法[15]。
甘蓝材料叶片相关指标:过氧化物酶(POD)活性,采用愈创木酚氧化法[16];超氧化物歧化酶(SOD)活性,采用NBT 光还原法[17];苯丙氨酸解氨酶(PAL)活性,采用王敬文和薛应龙(1981)[18]的方法。
1.3 数据处理
各处理重复3 次,结果计算平均值,然后采用Microsoft Office Excel 2010 和SPSS 17.0 对试验数据进行统计分析,用Duncan’s 多重比较分析差异显著性。
2 结果分析
2.1 接种黑腐病菌对不同抗性甘蓝材料根系的影响
由表1 可知,与未接种黑腐病菌的5 个甘蓝材料幼苗相比,在接种后第1 天,接种幼苗根长和根系鲜重变化均较小,均未达到显著水平(P<0.05);QG70(R)幼苗根系活力表现出显著升高。在接种后第7 天,经接种后5 个材料的根长、根系鲜重均不同程度低于未接种材料,其中QG50(S)、B02(HS)经过接种的幼苗根长明显低于未接种幼苗,并且差异可达到极显著水平(P<0.01),QG70(R)、QG50(S)和B02(HS)经接种处理的幼苗根系鲜重显著低于未接种幼苗;接种的QK2502(HR)、QG70(R)、LQ66(MR)、QG50(S)的根系活力均高于未接种幼苗,B02(HS)的根系活力低于未接种幼苗,其中QK2502(HR)经过接种的幼苗根系活力增加率为21.04%,与未接种幼苗差异达到极显著水平(P<0.01),LQ66(MR)经过接种的幼苗根系活力显著高于未接种幼苗,增加率为15.27%。

表1 接种黑腐病菌对甘蓝幼苗根系的影响
比较5 个甘蓝材料可知,在第7 天,接种比未接种增加率表现为:根长增加率由低到高依次为B02(HS)、QG50(S)、QK2502(HR)、LQ66(MR)、QG70(R);根系鲜重增加率由低到高依次为B02(HS)、QG50(S)、QG70(R)、LQ66(MR)、QK2502(HR);根系活力增加率由低到高为B02(HS)、QG50(S)、QG70(R)、LQ66(MR)、QK2502(HR),其中B02(HS)根系活力增加率为负值。图1 为5 个甘蓝材料接种黑腐病菌和未接种幼苗第1 天和第7 天根系形态对比情况。

图1 黑腐病菌对甘蓝幼苗根系的影响(第1天和第7天)
2.2 接种黑腐病菌对不同抗性甘蓝材料叶片POD活性的影响
由表2 可知,接种黑腐病菌后,5 个材料幼苗叶片POD 活性均不同程度增加,增加率表现出先升高(第1、3、5 天)后降低(第7 天)的趋势。QK2502(HR)在接种后1、3、5、7 d 叶片POD 活性均高于未接种幼苗,且达到极显著水平(P<0.01),增加率分别为29.77%、35.80%、38.83%、33.79%。QG70(R)在接种后1、5、7 d 叶片POD 活性均高于未接种幼苗,且达到极显著水平(P<0.01);在接种后3 d 叶片POD 活性显著高于未接种幼苗,未达到极显著水平(P<0.01)。LQ66(MR)在接种后1 d 叶片POD 活性虽高于对照,但未达到显著水平(P<0.05);在接种后3、5、7 d 叶片POD 活性均极显著高于未接种幼苗,增加率分别为17.70%、21.06%、16.21%。QG50(S)在接种后5 d 叶片POD 活性极显著高于未接种幼苗,增加率达18.65%。B02(HS)在接种后5、7 d 叶片POD 活性极显著高于未接种幼苗,增加率分别为15.35%、7.71%。

表2 接种黑腐病菌对叶片过氧化物酶(POD)活性的影响
比较5 个甘蓝材料可知,接种比未接种幼苗叶片POD 活性增加率表现为:第1 天增加率由高到低依次为QK2502(HR)、QG70(R)、QG50(S)、LQ66(MR)、B02(HS);第3、5、7 天增加率由高到低为QK2502(HR)、QG70(R)、LQ66(MR)、QG50(S)、B02(HS)。
2.3 接种黑腐病菌对不同抗性甘蓝材料叶片SOD活性的影响
由表3 可知,QK2502(HR)、QG70(R)、LQ66(MR)、B02(HS)在接种后幼苗叶片SOD 活性增加率表现出先升高(1、3 d)后降低(5 d)再升高(7 d)的趋势,QG50(S)表现出先升高(1 d)后降低(3、5、7 d)的趋势。QK2502(HR)接种后1、3、7 d 叶片SOD 活性均高于未接种幼苗,且达到极显著水平(P<0.01),增加率分别为4.43%、4.48%、10.51%。QG70(R)接种后3、7 d 叶片SOD 活性均极显著高于未接种幼苗,增加率分别为4.58%、3.09%。LQ66(MR)和B02(HS)接种后幼苗叶片SOD 活性与未接种幼苗相比均有差异,但未达到显著水平(P<0.05)。QG50(S)接种后3、5、7 d 叶片SOD 活性均显著低于未接种幼苗,增加率分别为-5.14%、-10.99%、-12.58%。

表3 接种黑腐病菌对叶片超氧化物歧化酶(SOD)活性的影响
比较5 个甘蓝材料可知,接种比未接种幼苗叶片SOD 活性增加率表现为:第1 天增加率由高到低依次为B02(HS)、QK2502(HR)、QG50(S)、LQ66(MR)、QG70(R);第3 天增加率由高到低依次为QG70(R)、QK2502(HR)、LQ66(MR)、B02(HS)、QG50(S);第5 天增加率由高到低依次为QK2502(HR)、QG70(R)、B02(HS)、LQ66(MR)、QG50(S);第7 天增加率由高到低依次为QK2502(HR)、QG70(R)、LQ66(MR)、B02(HS)、QG50(S)。
2.4 接种黑腐病菌对不同抗性甘蓝材料叶片PAL活性的影响
由表4 可知,接种黑腐病菌后,5 个材料幼苗叶片PAL 活性表现出不同变化,增加率表现出逐渐降低的趋势。QK2502(HR)在接种后1、3、5、7 d 叶片PAL 活性均高于未接种幼苗,且达到极显著水平(P<0.01),增加率分别为12.48%、12.09%、9.43%、6.12%。QG70(R)在接种后1、3 d 叶片PAL 活性均极显著高于未接种幼苗,增加率分别为9.40%、6.11%;接种后5、7 d 叶片PAL 活性均高于未接种幼苗,但未达到显著水平(P<0.05)。LQ66(MR)在接种后1、3 d 叶片PAL 活性均高于未接种幼苗,且达到极显著水平(P<0.01),增加率分别为6.06%、4.38%;接种后7 d 叶片PAL 活性显著低于未接种幼苗,增加率为-4.50%。QG50(S)在接种后1 d 叶片PAL 活性极显著高于未接种幼苗;接种后3、7 d叶片PAL 活性均低于未接种幼苗,且达到极显著水平(P<0.01)。B02(HS)在接种后1 d 叶片PAL 活性高于未接种幼苗,但差异不显著(P<0.05);接种后3、5、7 d 叶片PAL 活性均显著低于未接种幼苗,且第3、7 天差异均达到极显著水平(P<0.01)。
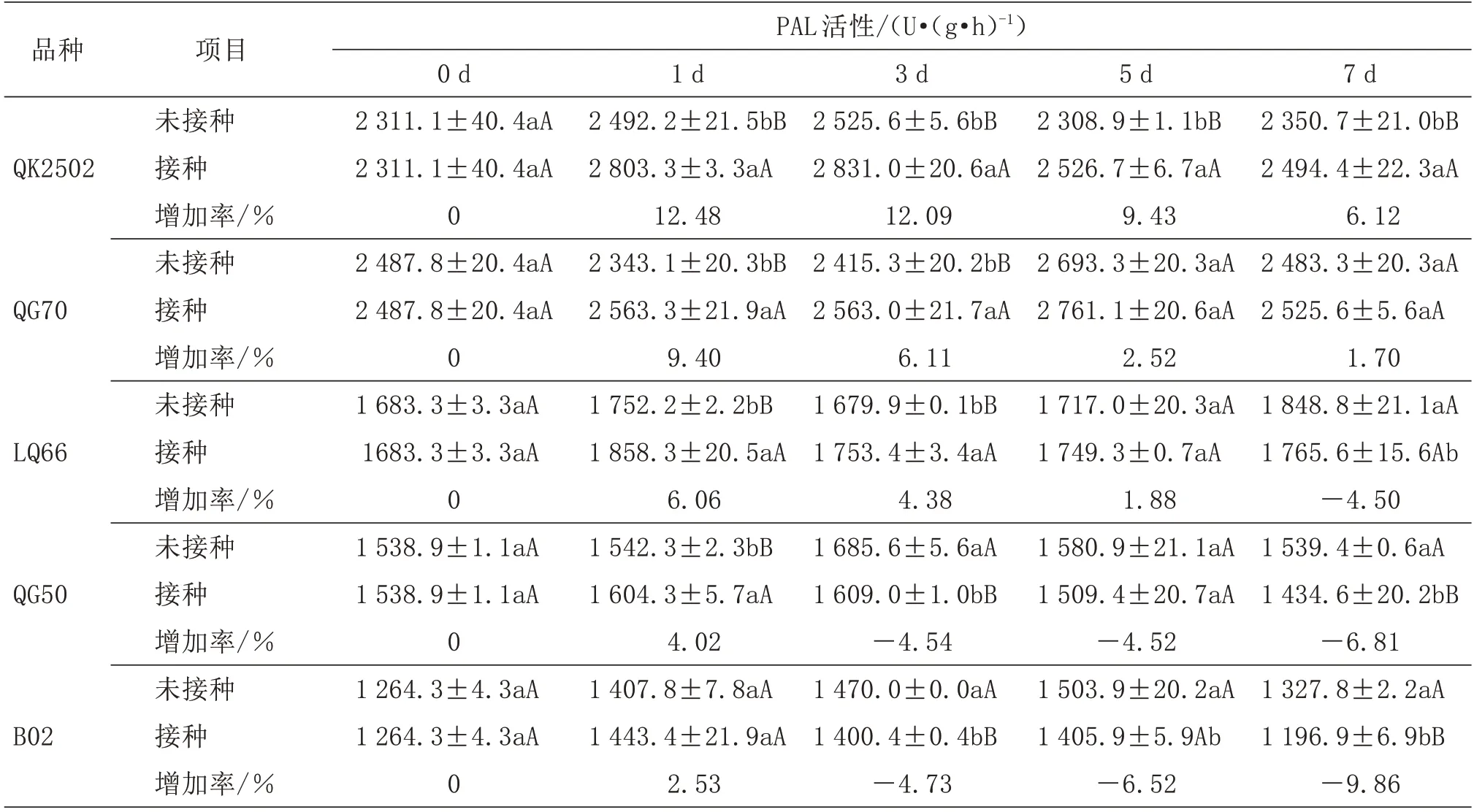
表4 接种黑腐病菌对叶片苯丙氨酸解氨酶(PAL)活性的影响
比较5 个甘蓝材料可知,在接种后1、3、5、7 d,接种比未接种幼苗叶片PAL 活性增加率由高到低为QK2502(HR)、QG70(R)、LQ66(MR)、QG50(S)、B02(HS)。
3 讨论与结论
根系生长状况对植物生长起直接作用,关于抗病性和根系生理生化情况变化之间的关系已有学者作了大量研究。邵金旺等(2001)[19]研究甜菜丛根病不同抗性品种根系活力发现,抗病品种的根系活力明显高于感病品种。吴晓丽等(2011)对接种黑腐病菌后花椰菜根系的形态和生理变化进行研究发现,接种处理的植株根长、根干重明显比未接种的植株(CK)低,但是接种幼苗的根系活力比未接种幼苗的根系活力高。本试验结果表明,接种黑腐病菌的幼苗根长和根系鲜重明显低于未接种幼苗,且抗病品种的根长和根系鲜重降低率低于感病品种;而根系活力则相反,经过接种的幼苗根系活力高于未接种幼苗,且抗病品种的根系活力增加率高于感病品种。这可能是由于黑腐病菌侵染植株,根系生长受到抑制,导致根长和根系鲜重小于未接种植株,抗病品种的防御能力较感病品种强,故抗病品种的根长和根系鲜重降幅小于感病品种;病原菌侵染,植物会启动防御机制,根系也会发生一系列保护性反应,植株根系对于病原菌的防御能力增强,从而出现根系活力升高,且抗病品种根系活力升高幅度高于感病品种的现象。在第7 天,B02(HS)根系活力低于未接种植株,可能是由于此时植株的保护机制已经被破坏。
过氧化物酶(POD)虽无直接的抗菌活性,但其可以通过影响植物体内的多种代谢途径而在抗病性中起到间接作用。当植物受到病原菌侵染时,体内会产生大量的活性氧,SOD 是清除活性氧的主要酶之一。苯丙烷类代谢途径是植物在抗病反应体系中的一个重要途径,而苯丙氨酸解氨酶是该途径的关键酶和限速酶[20]。已有学者对抗病性与POD活性、SOD 活性、PAL 活性的关系进行了研究。吴应海等(2022)[21]研究发现,为百香果接种茎基腐病后,植物体内POD 活性、SOD 活性、PAL 活性均有升高,且抗病性越强酶活性越高。黄志磊等(2019)[22]研究表明,不同抗性大麦品种在受到叶斑病菌侵害时POD 活性、SOD 活性、PAL 活性均升高并且随着时间延长增幅逐渐增加,且抗病品种蒙啤麦3 号升高幅度均大于感病品种蒙啤麦1 号。本研究对QK2502、QG70、LQ66、QG50、B02 这5 个抗性不同的甘蓝品种接种黑腐病菌后,叶片POD 活性均不同程度增加,其中从第3 天开始抗病品种的增加率均高于感病品种,由高到低依次为QK2502(HR)、QG70(R)、LQ66(MR)、QG50(S)、B02(HS);SOD 活性增加率表现出先升高(1、3 d)后降低(5 d)再升高(7 d)的趋势,QG50(S)表现出先升高(1 d)后降低(3、5、7 d)的趋势。接种初期,抗病性与SOD 活性变化率之间没有表现出较强的相关性,但随着时间推移,抗病品种的SOD 活性增加率普遍高于感病品种,与其他学者的研究相吻合。叶片POD 活性、SOD 活性增加可能是植株为了抵抗黑腐病菌侵害产生的保护性反应,从而增加植株对黑腐病菌的抵抗能力,抗病品种的抵抗力较感病品种强,体现在POD 活性、SOD 活性增加率上则为抗病品种高于感病品种。PAL 活性表现为抗病品种普遍高于感病品种,与其他学者研究一致。接种后第1 天,无论是抗病还是感病品种,PAL活性增加率均达到峰值,之后开始下降,PAL 活性增加率与抗病性呈明显正相关,但3、5、7 d 的QG50、B02 以及7 d 的LQ66 叶片PAL 活性低于对照,这可能是由于植株自身的防御系统被病原菌破坏所导致。
综上,接种黑腐病菌后导致甘蓝幼苗根系生长缓慢,根长和根系鲜重增加较慢,均低于未接种植株,且抗病品种受病原菌侵染的影响小于感病品种;根系活力相反,病原菌侵染会使根系活力提高,且抗病品种根系活力增加率高于感病品种。接种黑腐病菌后,植株的防御机制启动,POD 活性、SOD活性和PAL 活性升高,且抗病品种酶活性增加率普遍高于感病品种。因此,根长、根系鲜重、根系活力、叶片POD 活性、叶片SOD 活性、叶片PAL 活性可以作为甘蓝抗黑腐病种质资源鉴定的辅助指标。
猜你喜欢
浙江林业科技(2021年3期)2021-07-13 09:03:56
新农村(2018年24期)2018-09-14 06:55:38
长江大学学报(自科版)(2018年14期)2018-07-17 08:37:02
中国中药杂志(2017年11期)2017-06-22 11:37:34
中国中药杂志(2017年1期)2017-03-06 21:19:22
河北林业科技(2016年4期)2016-02-05 08:07:21
湖北农业科学(2015年23期)2016-01-08 20:37:10
凤凰资讯报(2014年36期)2014-04-29 00:44:03
食品工业科技(2014年14期)2014-03-01 09:56:48
长江大学学报(自科版)(2013年23期)2013-10-27 08:45:37
