
补阳还五汤通过调控PI3K/Akt、JAK2/STAT3信号促进BMSC趋化迁移对外伤性脊髓损伤大鼠神经元活性及认知功能的影响
2023-10-07 12:21宋颖军李旭刘小舟张国福
中国老年学杂志 2023年17期
宋颖军 李旭 刘小舟 张国福
(江西中医药大学附属医院,江西 南昌 330006)
外伤脊髓损伤是中枢神经系统的严重损伤,多发生于交通事故、坠落等,是发病率及致残率较高的一种疾病。脊髓损伤后,脊髓内部岀血、水肿,出现神经元凋亡、坏死等现象,进而导致神经系统功能破坏〔1,2〕。目前认为,神经细胞凋亡是脊髓损伤后神经功能障碍的主要原因,磷脂酰肌醇-3激酶/蛋白激酶B(PI3K/Akt)具有调控细胞增殖及凋亡的作用。研究表明〔3〕PI3K、磷酸化(p-)Akt在脊髓损伤发展过程中表达升高,与脊髓损伤机体脊髓细胞凋亡有关。基因调控的启动需要复杂而周密的信号转导系统,内源性酪氨酸激酶(JAK)/信号传导和转录启动因子(STAT)通路是近年来备受关注的一条通路,具有调节细胞生物学行为及免疫功能的作用。研究表明〔4〕,脊髓损伤后可异常激活该通路,从而导致神经细胞凋亡。补阳还五汤是中国清代著名医家王清任所创立的经典方剂,该方由黄芪、赤芍、当归、地龙、川芎、桃仁和红花药物组成。已有报道证实〔5〕该方剂在促进损伤神经再生修复等方面有其独特的效果。本文探讨补阳还五汤通过调控PI3K/Akt、JAK2/STAT3信号促进骨髓间充质干细胞(BMSCs)趋化迁移对外伤性脊髓损伤大鼠的神经元活性及认知功能的影响。
1 材料和方法
1.1实验动物与分组 选取健康大鼠53只,购自北京亿鸣复兴公司提供〔生产许可证号:SCXK(京)-2019-0034〕。大鼠体质量220~300 g,常规饲养,自由饮食饮水。随机分为健康组(健康大鼠常规饲养)、损伤组(建立脊髓损伤模型)、干预组(脊髓损伤模型+补阳还五汤)、对照组(脊髓损伤模型+甲泼尼龙),每组12只,剩余5只大鼠用于补阳还五汤含药血清制备。本研究经医院动物伦理委员会批准。
1.2实验试剂 结晶紫溶液(南京生航公司);PI3K、p-PI3K、Akt、p-Akt一抗(广州柏赛柯公司);辣根过氧化物酶二抗(西安齐岳公司);伊红染色液(北京金克隆公司);TUNEL反应液(苏州优逸兰迪公司);苏木素染色液(大连芮禹公司);RIPA裂解液(武汉卡诺斯公司)。
1.3药物制备 补阳还五汤由医院制剂室加工,1 ml汤药含生药4 g,即4 g/ml质量浓度,无菌封瓶,-20 ℃冰箱保存备用。甲泼尼龙购自广州贝尔卡公司。
1.4模型建立及给药 健康组大鼠无处理,其余各组大鼠戊巴比妥钠麻醉,俯卧位固定在T8棘突中心作长约3 cm切口暴露硬脊膜,采用Allen装置以20 g×25 mm损伤力度行重物打击法制作大鼠外伤性脊髓损伤模型。模型制备成功判定标准:脊髓打击后,损伤处脊髓出血、水肿,大鼠出现摆尾反射,双下肢及躯体回缩样扑动,麻醉清醒后双下肢成弛缓性瘫痪〔6〕。建模成功后损伤组无处理,干预组给予制备好的补阳还五汤1 ml灌胃,1次/d,28 d〔7〕,对照组经腹腔注射给予甲泼尼龙30 mg/kg,1次/d,28 d。
1.5BMSCs分离养鉴定 根据文献〔8〕大鼠原代BMSCs提取方法,戊巴比妥钠麻醉处死大鼠,于75%酒精中浸泡消毒5~10 min。使用眼科剪将10只转T克隆绿色荧光蛋白(GFP)基因的大鼠双侧股骨及胫骨取出,浸泡于预冷的培养基,使用10 ml注射器冲洗骨髓腔直至股骨胫骨变白,1 000 r/min离心5 min。使用含有10%胎牛血清和1%双抗体(青霉素-链霉素)完全培养基重悬浮,置于T12的培养瓶中培养24 h后更换培养基。之后每隔2 d更换培养基。当细胞融合率达到70%~80%时进行细胞传代。光镜下检测细胞形态。
1.6脊髓组织提取 取各组大鼠,麻醉后处死,从背部原切口进入,切取2 cm损伤部位及其上下段脊髓组织,置于EP管中,-70 ℃备用。
1.7补阳还五汤含药血清的制备 取5只大鼠给予补阳还五汤3 ml灌胃,2次/d,共1 w,末次灌胃1 h后腹主动脉取血,离心后取上清,置于-20 ℃保存〔9〕。
1.8检测指标
1.8.1流式细胞术鉴定BMSCs细胞 取第三代细胞通过流式细胞术鉴定细胞表面标记CD29、CD90、CD45和CD31以检测细胞纯度。纯度评估后,使用第三代的细胞以1×105/ml密度在成骨和成脂专用培养基中进行BMSCs定向成骨细胞、成脂、成神经细胞分化诱导培养。
1.8.2Transwell小室法检测补阳还五汤对大鼠BMSCs迁移的影响 取BMSCs放入完全培养基内培养,于12 h观察BMSCs迁移情况。取第三代BMSCs,分为空白组(常规培养),含药血清组(加入2.6 g/ml补阳还五汤含药血清〔9〕),药物对照组(加入10-7mol/L甲泼尼龙〔10〕),培养24 h后,调整细胞密度为5×104/孔,100 μl接种于Transwell上室;下室加入完全培养基600 μl,37 ℃培养12 h后取出Transwell,吸掉上下室培养液,用磷酸盐缓冲液(PBS)洗3遍,4%多聚甲醛固定30 min。晾干,擦去上层未迁移细胞。结晶紫染色20 min。使用光学显微镜进行观察,随机选择5个视野进行细胞计数。重复3次,每次3个复孔。
1.8.3高架十字迷宫和Morris水迷宫实验检测大鼠认知功能 脊髓损伤后8 w进行。常温,调节灯光,测试5 min,每只测试1次,安抚大鼠使之适应环境。记录开放臂进入次数及迷宫内进行的总路程。用Smart2.0软件分析数据。
于损伤后第7周对大鼠进行水迷宫实验训练,于第8周测试。测试分为定位航行:4 d,从迷宫4个象限入水,每天第1次训练均从目标象限对侧放入迷宫内;空间探索:3 d,取消平台,在1.5 min内记录大鼠穿越平台次数。Smart2.0软件分析数据。
1.8.4苏木素-伊红(HE)染色检测脊髓组织病理形态 取各组脊髓组织,脱蜡、水洗后,苏木素染色15 min。经酸水及氨水中分色数秒。流水冲洗后入经70%、90%乙醇脱水。再置于伊红染色液染色2~3 min。染色后的切片经无水乙醇脱水,二甲苯透明后滴加树胶,封片固定,于光学显微镜下观察。
1.8.5TUNEL检测脊髓组织神经细胞凋亡 脊髓组织石蜡切片经二甲苯、梯度乙醇洗脱后,PBS冲洗,置于含2%蛋白激酶K消化15 min,PBS冲洗2次。滴加TUNEL反应液,避光孵育1 h,PBS冲洗2次,加入DAPI、22%甘油缓冲液封片。荧光显微镜下观察并计算神经细胞凋亡数。
1.8.6免疫组化检测p-JAK2、p-STAT3水平 取各组大鼠脊髓组织标本,常规切片,按SP试剂盒说明书进行操作:烤片、脱蜡至水、高压修复、3%过氧化氢(H2O2)去离子水处理、封闭、滴加一抗过夜(p-JAK2 1∶50、 p-STAT3 1∶100),次日滴加二抗、金黄色葡萄球菌蛋白A-辣根过氧化物酶(HRP)标记抗原工作液,二氨基联苯胺(DAB)显色,蒸馏水终止显色,PBS冲洗,苏木素复染、冲洗、梯度酒精脱水,透明,中性树胶封片。光镜观察,以细胞质棕黄色着色为阳性细胞,取 3个视野,分别计数每个视野的阳性细胞数,取平均值。
1.8.7Western印迹检测脊髓组织PI3K、p-PI3K、Akt、p-Akt水平 取脊髓组织,RIPA裂解液裂解,离心。收集上清液,二喹啉甲酸(BCA)法测定蛋白含量。取适量蛋白质,加入上样缓冲液,95 ℃煮沸5 min,进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)。电泳后,将分离的蛋白条带转至聚偏氟乙烯(PVDF)膜,5%脱脂牛奶室温封闭2 h之后加入PI3K、p-PI3K、Akt、p-Akt一抗,4 ℃孵育过夜。TBST清洗,加入HRP二抗,37 ℃孵育2 h-TBST洗膜,加入电化学发光(ECL)液避光显影。ImageJ图像软件分析各蛋白条带的灰度值。
1.9统计学分析 采用SPSS17.0软件进行单因素方差分析、t检验。
2 结 果
2.1BMSCs鉴定结果 传代后的培养细胞呈旋涡状或放射状贴壁生长,细胞多呈星形、梭形或三角状,培养3代后,细胞贴壁加快、形态均一,呈旋涡状或单层放射状生长。见图1。
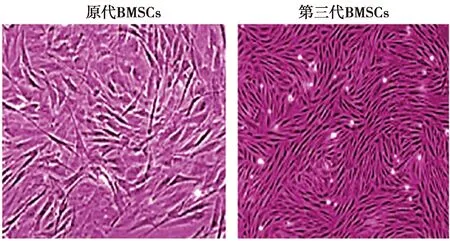
图1 各组BMSCs细胞形态(×200)
2.2BMSCs迁移结果 空白组BMSCs细胞迁移数量〔(243.32±25.68)个〕显著低于含药血清组〔(309.45±34.81)个〕与药物对照组〔(304.72±38.27)个〕(P<0.05)。含药血清组与药物对照组细胞迁移数量无统计学差异(P>0.05)。见图2。
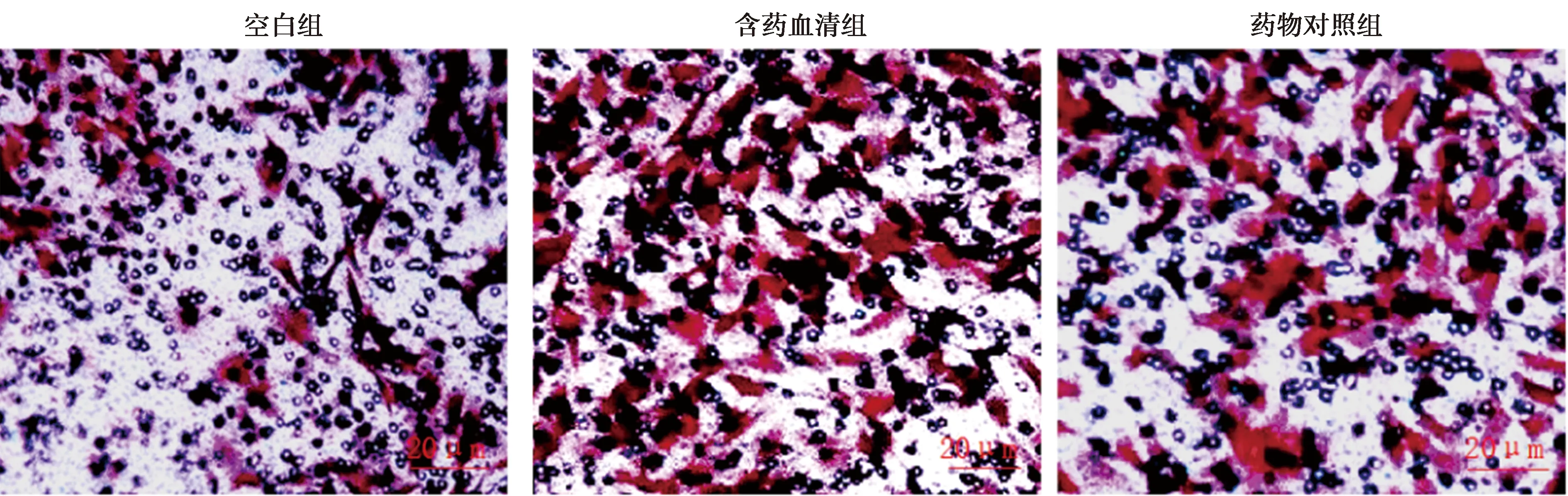
图2 各组BMSCs迁移(结晶紫染色,×400)
2.3各组认知功能检测 与健康组相比,损伤组总路程、进入开臂次数、穿越平台次数显著降低,不同时间的潜伏期显著升高(P<0.05)。与损伤组相比,干预组和对照组总路程、进入开臂次数、穿越平台次数显著升高,不同时间的潜伏期显著降低(P<0.05)。干预组与对照组各指标对比无统计学差异(P>0.05)。见表1。
表1 各组认知功能对比
2.4HE染色结果 健康组脊髓组织结构完整。损伤组脊髓组织疏松水肿,有细胞空泡变性产生。相较于损伤组,干预组与对照组脊髓组织病理形态有所改善。见图3。
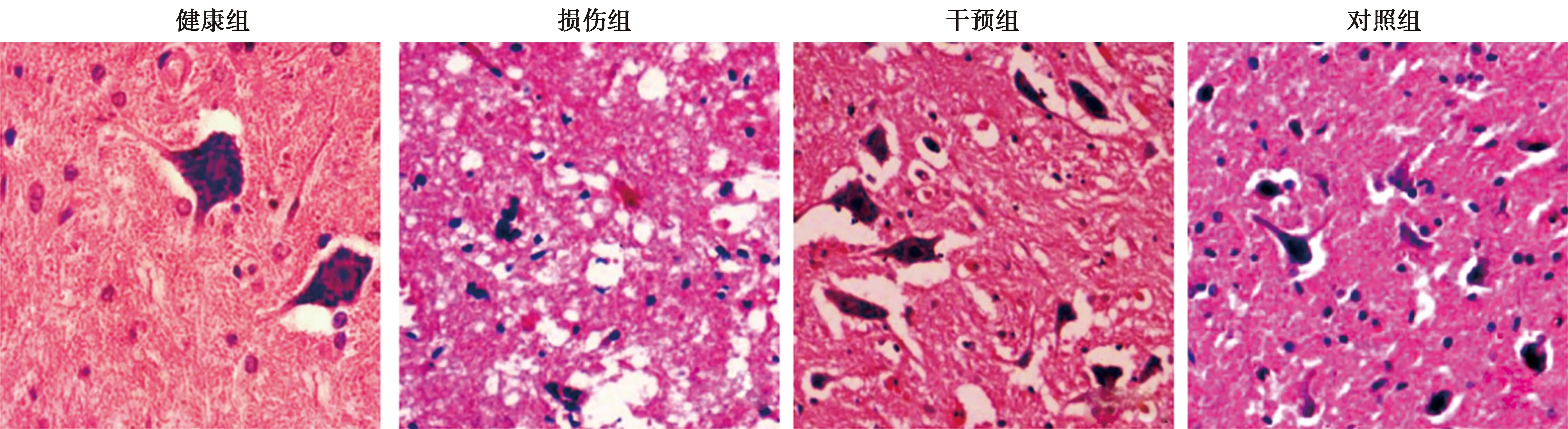
图3 各组脊髓组织病理学形态(HE染色,×200)
2.5神经细胞凋亡率检测结果 健康组脊髓组织神经细胞凋亡率〔(1.28±0.64)%〕显著低于损伤组〔(31.42±4.97)%〕(P<0.05)。损伤组凋亡率显著高于干预组〔(11.04±2.13)%〕和对照组〔(12.46±2.07)%〕(P<0.05)。干预组凋亡率与对照组无统计学差异(P>0.05)。见图4。
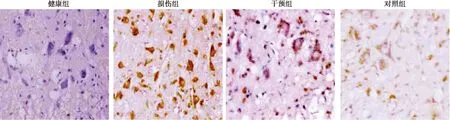
图4 各组脊髓组织神经细胞凋亡(TUNEL染色,×400)
2.6p-JAK2、p-STAT3水平检测结果 健康组脊髓组织中p-JAK2、p-STAT3阳性细胞数显著低于损伤组(P<0.05)。损伤组p-JAK2、p-STAT3显著高于干预组和对照组(P<0.05)。干预组与对照组各指标对比无统计学差异(P>0.05)。见表2、图5。
表2 各组脊髓组织p-JAK2、p-STAT3阳性细胞数及PI3K、p-PI3K、Akt、p-Akt水平对比
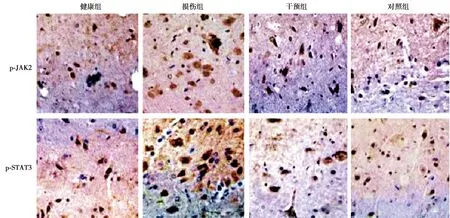
图5 各组p-JAK2、p-STAT3阳性细胞数(免疫组化染色,×400)
2.7PI3K、p-PI3K、Akt、p-Akt水平检测结果 健康组脊髓组织中PI3K、p-PI3K、Akt、p-Akt水平显著高于损伤组(P<0.05)。损伤组PI3K、p-PI3K、Akt、p-Akt水平显著低于干预组和对照组(P<0.05)。干预组与对照组PI3K、p-PI3K、Akt、p-Akt水平无统计学差异(P>0.05)。见表2、图6。
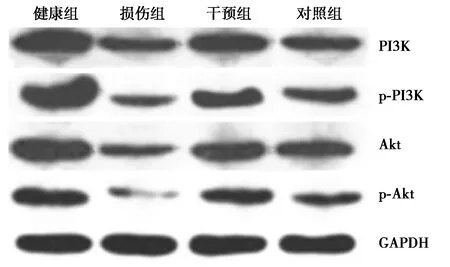
图6 各组PI3K、p-PI3K、Akt、p-Akt水平
3 讨 论
BMSCs是一种具有良好分化能力且较容易分离获取的成体干细胞,作为种子细胞在治疗颅脑损伤和脊髓损伤方面取得了较好疗效。其治疗脊髓损伤的机制主要有作为种子细胞迁移到受损部位分化替代受损的细胞,合成分泌生长因子,如血管内皮生长因子(VEGF)、神经胶质源性神经生长因子等促进组织修复,BMSCs还有良好的抗炎及免疫调节等功能并能够抑制小胶质细胞过度活化的能力。中医认为脊髓损伤属于督脉受损,经脉瘀阻,加之患者多因外伤而大伤元气,导致血瘀气虚之症,近年来已有研究证实〔11〕补阳还五汤对脊髓损伤患者具有一定的治疗作用,且补阳还五汤中当归尾具有活血祛瘀、补血通经、止痛活络等功效;赤芍具有清热凉血、化瘀等功效;桃仁具有活血化瘀、抗炎镇痛等作用。甲泼尼龙主要可以起到消炎的作用,是一种短效激素,起效快,副作用小,治疗脊髓损伤效果较好。秦丰伟等〔12〕运用大鼠建立脊髓损伤模型并进行的实验中,发现淫羊藿苷联合BMSCs通过降低脊髓损伤区域的炎症水平,改善大鼠脊髓损伤病情的严重程度。范瑜洁等〔13〕在对脊髓损伤大鼠的实验中提出,补阳还五汤可促进BMSCs增殖并向神经肝细胞方向分化。本文研究与上述研究结果相似。
以往对于脊髓损伤的实验中,主要灌注的为运动、感觉功能障碍等,对于认知功能等易被忽视。已有研究证实〔14〕,脊髓损伤后患者会出现记忆障碍、焦虑等。本文进一步完善了脊髓损伤大鼠认知功能的行为血评价,并增加了焦虑测试,证实了大鼠在8 w后会出现学习和空间记忆能力衰减,通过高架十字迷宫实验发现,大鼠还具有抑郁、探索兴趣减退等情绪障碍。房涛等〔15〕通过将脑梗死患者分为对照组与观察组并运用补阳还五汤进行治疗发现,补阳还五汤可通过提高患者的血清胰岛素样生长因子(IGF)-1水平,改善神经功能缺损及认知功能。本研究结果与上述研究相似。
脊髓损伤是临床上常见的中枢神经系统疾病,致残率较高。脊髓损伤的主要病理变化是神经细胞凋亡,神经细胞凋亡以核染色质固缩、DNA断裂、电泳呈梯带分布为特征,是脊髓损伤后神经细胞死亡的主要方式。研究发现,在中枢神经系统缺血性和机械性损伤模型中,广泛存在DNA损伤,可表现为神经细胞凋亡 、DNA的片段化和转录水平修复相关基因表达等〔16〕。TUNEL可以检测细胞基因组DNA断裂等,从而反映细胞凋亡情况。本研究提示,补阳还五汤能够在一定程度上减弱损伤后脊髓神经元凋亡的现象,从而进行神经保护 。黄芪与当归是补阳还五汤中的两味药材。李虎虎等〔17〕在实验中提出,黄芪具有抑制糖尿病大鼠神经细胞凋亡的作用。朱丽娟等〔18〕在实验中提出,当归通过激活细胞外调节蛋白激酶(ERK)信号通路可抑制缺血再灌注神经细胞的凋亡。本研究显示,在经过补阳还五汤药物干预后,神经细胞凋亡减少,提示其具有保护神经的作用。
有研究显示〔19〕,神经元损伤后,可进行自我修复,但此过程需要多种基因参与调控,包括神经生长因子和凋亡相关基因等,而这些基因又受到信号转导通路的调控。研究发现〔20〕,PI3K/Akt通路不仅在胰岛素调节糖代谢中发挥重要作用,还能促进神经细胞的存活和抗凋亡,并调节神经细胞的分化和运动。Akt是PI3K的主要靶蛋白之一,Akt通常磷酸化后被激活,磷酸化的Akt可通过激活蛋白激酶C 和线粒体ATP敏感性钾离子通道,改善线粒体功能,抑制线粒体释放凋亡因子,在细胞凋亡调控过程中发挥关键作用。有研究提出〔21〕,丹参酮ⅡA可能通过提高脊髓损伤大鼠脊髓组织PI3K、Akt mRNA和蛋白表达水平促进脊髓神经功能的恢复。另研究表明〔22〕,PI3K/Akt通路激活通过调节抗凋亡和自噬反应在脊髓损伤中起着重要的神经保护作用。 Akt磷酸化可削弱神经细胞凋亡相关下游分子的表达,如裂解的含半胱氨酸的天冬氨酸蛋白水解酶(Caspase)-3、6、9,B淋巴细胞瘤-2基因(Bcl-2)和Bcl-2相关X蛋白(Bax),导致Bcl-2/自噬基因苄氨素(Beclin)-1复合物的解离,该过程释放Beclin-1并导致自噬反应。在吴晓光等〔23〕对脑出血模型大鼠的实验研究中提出,补阳还五汤通过激活PI3K并与其下游因子Akt结合,使Akt从细胞质向细胞膜转移的同时,改变其构象,使Akt的Thr308位点及Ser473位点发生磷酸化,活化的Akt通过调控Bcl-2等凋亡相关但表达的表达,从而抑制脑出血神经细胞凋亡,保护脑组织。本研究发现,在脊髓损伤中PI3K、Akt水平降低,经过药物干预后PI3K、Akt水平升高。陈博威等〔24〕对脑出血小鼠的实验中提出,补阳还五汤通过窖蛋白(Cav)-1调控PI3K/Akt信号通路活性,改善病情严重程度。Qiu等〔25〕针对脑出血患者的实验中提出,补阳还五汤能够激活人CXC趋化因子受体(CXCR)4-PI3K 自噬轴信号传导通路,既能够激发细胞自噬,又能够调控细胞自噬状态,实现抑制细胞凋亡的脑保护作用。本文研究与上述研究结果相似,推测补阳还五汤可能是通过促进PI3K、Akt激活PI3K/Akt通路,发挥神经保护作用,减少神经细胞凋亡,改善病情。且本文HE染色结果与TUNEL结果也可证实上述结论。
近年来,国内外学者发现,JAK2/STAT3 信号转导通路在免疫系统和肿瘤相关疾病中重要作用。该通路是多种细胞因子和生长因子信号转导的共同通路,在调控细胞生物学行为中具有重要作用。脊髓损伤可激活JAK/STAT 信号转导通路。已有研究证实〔26〕,脊髓损伤后 STAT3 信号转导对神经细胞凋亡有重要的调节作用。相关研究报道〔27〕,脑缺血后可激活JAK2/STAT3 通路,p-STAT3入核后可调控大量基因水平,直接参与神经元凋亡。研究发现〔28〕,JAK2/STAT3 信号通路可诱导大量的大脑皮质神经元细胞凋亡。另有研究提出〔29〕,JAK2/STAT3 信号转导通路在许多实体肿瘤和血液肿瘤中被激活,可能与其促进生长和抗凋亡作用有关。在杜全宇〔30〕研究肺纤维化的实验中提出,补阳还五汤通过抑制JAK2/STAT3 通路水平,抑制上皮细胞和成纤维细胞向肌成纤维细胞转化,提示补阳还五汤可抑制JAK2、STAT3表达。本文研究发现,脊髓损伤后JAK2、STAT3水平升高,在经过药物干预后JAK2、STAT3水平降低。王凯华等〔31〕在对脑出血大鼠的实验研究中提出,补阳还五汤通过抑制JAK2/STAT3 通路,减少神经细胞凋亡,改善神经功能,从而发挥治疗作用。
综上所述,补阳还五汤通过激活PI3K/Akt通路,抑制JAK2/STAT3信号通路的激活,促进BMSCs的迁移,减轻神经细胞的凋亡,起到神经保护的作用,从而改善脊髓损伤大鼠的认知功能。
猜你喜欢
军事文摘(2022年8期)2022-11-03
中成药(2021年5期)2021-07-21
Digital Chinese Medicine(2021年4期)2021-02-14
中国运动医学杂志(2016年3期)2016-07-10
中国组织化学与细胞化学杂志(2016年4期)2016-02-27
中国当代医药(2015年29期)2015-03-01
发明与创新(2015年37期)2015-02-27
癌变·畸变·突变(2014年1期)2014-03-01
中国中医药现代远程教育(2014年22期)2014-03-01
中国中医药现代远程教育(2014年21期)2014-03-01
