
宁都黄鸡GH 基因SNP 与鸡冠性状的关联分析
2023-09-21 15:28:46马荆鄂曾成瑶谭玉文朱学农饶友生许继国
广东农业科学 2023年7期
马荆鄂,曾成瑶,周 敏,谭玉文,朱学农,饶友生,许继国
(南昌师范学院生物技术研究院/江西省地方鸡种遗传改良重点实验室,江西 南昌 330032)
【研究意义】宁都黄鸡是江西省特优质型黄羽肉鸡,具有“三黄”(喙、胫、羽)、“五红”(冠、髯、脸、眼圈、耳叶)[1]、性早熟、体小、矮脚等特征,以及肉质鲜美、口感丰富等优点。其肌肉中的蛋白质及必需氨基酸含量、单位面积肌纤维数量等指标均优于国内多个肉鸡品种[2-4],因此深受广大消费者喜爱,但宁都黄鸡繁殖性能低,影响其产业的健康发展。鸡冠性状不仅是消费者选购禽肉产品的重要参考因素,同时冠长、冠高与产蛋量、平均蛋重等繁殖性状有显著相关性[5-7]。因此,对鸡冠性状进行分子标记辅助选择,是优质肉鸡选育的重要任务之一。【前人研究进展】鸡冠是鸡头部的皮肤衍生物,缺乏羽毛,公鸡鸡冠十分发达,是其第二性征,是衡量鸡性成熟的重要指标[8-9]。鸡冠始于上喙的一个区域,位于前鼻突的后方,由表皮、真皮和中央层组成。表皮具有角质层和生发层,角质层位于表皮最外层,对鸡冠皮下组织起保护作用,防止皮下组织受到感染[10]。鸡冠是一类变异丰富且易于观察的外观性状,其相关研究多从基因的角度进行遗传分析,目前已发现一些常见冠型的改变均源自基因序列变异和异位表达[11]。研究发现,AR、BMP2、CHADL、DRD1、EDN1、GARNL1、HSPA5、HSPA8、STK32A、PIK3R1、VIP1、VIP2和WNT9A基因具有调节鸡冠生长发育的作用,是鸡冠性状选育的重要候选基因[12-16]。【本研究切入点】生长激素(Growth hormone,GH)基因能够调节糖类、脂肪和蛋白质的代谢,促进机体各个组织的生长发育[17]。GH基因序列中存在多个SNP[18-19],部分多态性位点与开产体质量、开产日龄以及生长性状有显著关联性[20]。目前关于GH基因SNP 与宁都黄鸡公鸡鸡冠性状的关联分析少见报道。【拟解决的关键问题】研究GH基因上游调控区域(5'-侧翼区)和内含子1 区域序列单核苷酸多态位点对宁都黄鸡公鸡鸡冠性状的影响,寻找地方鸡种鸡冠性状选育的分子标记。选取前期筛选的GH基因SNP,采用SAS 软件分析鸡冠性状关联的SNP,解析优势基因型,为宁都黄鸡鸡冠性状的分子辅助选育积累理论基础,进而为改良宁都黄鸡的繁殖性状奠定基础。
1 材料与方法
1.1 试验材料
供试的499 只4~16 周龄宁都黄鸡公鸡的鸡冠表型数据来源于实验室前期记录[3],包括7 个周龄(分别为4、6、8、10、14、16 周龄)的冠高、冠长、冠厚、肉垂长、肉垂厚和冠齿数 6 个鸡冠性状指标。
1.2 试验方法
1.2.1 基因频率和基因型频率 假设基因座两个等位基因分别为A、a,基因型为AA、Aa、aa,则基因频率(p、q)、基因型频率(PAA、PAa、Paa)计算公式如下:
式中,n表示该基因型个体数。
1.2.2 Hardy-weinberg 平衡状态检验 对处于Hardy-weinberg 平衡状态下的群体,可以通过χ2适合性检验判定。
式中,Ei为理论期望值,Oi为实际观察值,n为等位基因数。
1.2.3 遗传多样性分析 采用群体杂合度(Heterozygosity,He)、群体纯合度(Homozygosity,Ho)和多态信息含量(Polymorphism information content,PIC)等衡量种群内的遗传多样性。
式中,Pi为任意位点上第i个等位基因频率。
1.2.4 基因型与性状关联分析 以实验室前期筛查到的GH基因5'-侧翼区域和内含子1 中的45 个SNP 为鸡冠性状候选标记位点[21],采用SAS8.1 软件GLM 模块,对所筛选SNP 与鸡冠性状进行关联分析,采用的混合模型如下:
式中,Y为表型变异,µ为群体均值,G为基因型效应,D为母体效应,H为批次效应,e为残差效应。
等位基因的加性效应是用2 个纯合子离差的均值计算,加性效应=(AA-BB)/2;等位基因显性效应是用杂合子与2 种纯合子均值的离差来计算,显性效应=AB-(AA+BB)/2。其中,AA、BB 和AB分别为基因型AA、BB和AB的表型最小二乘均值。显著水平检验水平为双侧0.05,对于多重比较采用Bonferroni 法进行校正,在遗传多态性关联分析中,校正次数为同一基因中检验的不同SNP个数。
2 结果与分析
2.1 GH 基因45 个SNP 基因频率和基因型频率分析
分别计算GH基因5'-侧翼区域和内含子1 中45 个SNP 的基因频率、基因型频率,以及He、Ho、PIC,结果显示(表1),有14 个位点的PIC> 0.25;经χ2适应性检验显示,25 个位点的χ2值没有达到显著水平,处于Hardy-weinberg平衡状态。
2.2 GH 基因SNP 与鸡冠性状的相关性
采用SAS 软件将45 个SNP 与不同鸡冠性状指标进行关联分析,选取P<0.05 的位点。结果(表2)表明,13 个SNP 与7 个冠高性状指标显著关联,包括G1271800A、C1272225T、G1272384A、G1272611A、T1271213G、C1271211T、G1271198A、C1271195A、C1270902T、T1270805C、A1270769G、T1270451C 和G1270155A。其中,A1270769G 与7 个周龄冠高指标均显著关联,G1272611A 与5 个周龄冠高指标显著关联,T1271213G、C1270902T 分别与4个周龄冠高指标显著关联。5 个位点与16 周龄冠高显著关联,分别为C1272225T、G1272384A、G1272611A、A1270769G、T1270451C。经过Bonferroni 法校正后,A1270769G 与4 周龄冠高、C1272225T 与16 周龄冠高相关性达到显著水平。

表2 与宁都黄鸡冠高显著相关的13 个SNP 位点P 值Table 2 P value of 13 SNPs significantly related to comb height of Ningdu yellow chickens
10 个SNP 与7 个冠长性状显著关联(表3),分别为C1272225T、G1272611A、C1271211T、C1270902T、A1270769G、G1270640C、T1270578C、G1270503C、T1270451C、C1270032T。其中,C1271211T 与6 个周龄冠长指标显著关联,C1270032T 与5 个周龄冠长指标显著关联,G1272611A 与5 个周龄冠长指标显著关联;与16 周龄冠长显著关联的位点为C1272225T、G1272611A、C1271211T、T1270451C。经 过Bonferroni 法校正后,G1272611A 与16 周龄冠长的相关性达到显著水平。

表3 与宁都黄鸡冠长显著相关的10 个SNP 位点P 值Table 3 P value of 10 SNPs significantly related to comb length of Ningdu yellow chickens
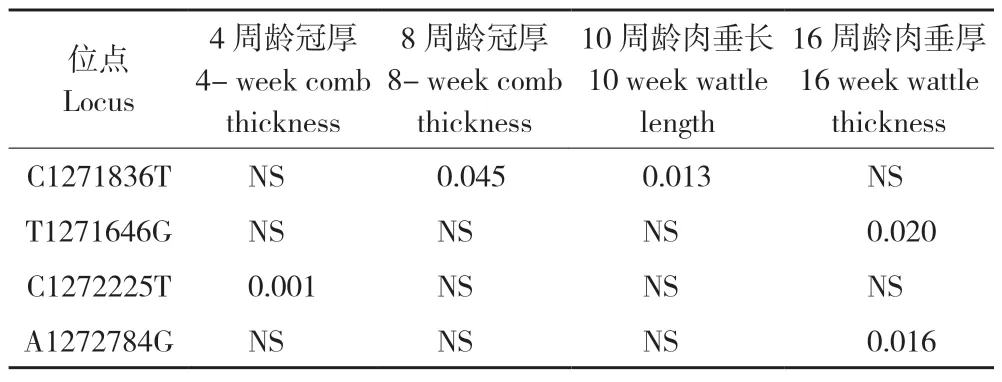
表4 与宁都黄鸡其他鸡冠性状显著相关的4 个SNP 位点P 值Table 4 P value of 4 SNPs significantly related to other comb traits of Ningdu yellow chickens
位点 C1271836T 分别与8 周龄冠厚、10 周龄肉垂长显著关联,T1271646G 与16 周龄肉垂厚显著关联,C1272225T 与4 周龄冠厚显著关联,A1272784G与16 周龄肉垂厚显著关联(表 4)。经过 Bonferroni法校正后,均未达到显著水平。此外,供试的 45个 SNP 与冠齿数性状均无显著关联。
2.3 A1270769G、G1272611A 位点不同基因型与宁都黄鸡冠高性状的相关性
分析宁都黄鸡公鸡GH基因A1270769G、G1272611A 两个位点不同基因型与7 个周龄冠高性状的相关性,并计算其加性效应和显性效应值(表5)。多重比较分析发现,A1270769G 位点的不同基因型中,AA 型个体的4、6、8、10、14、16 周龄冠高显著低于AG 和GG 型个体,12周龄冠高的AA 型个体显著低于AG 型个体;从位点对冠高性状的作用方式看,该位点主要以加性方式起作用,GG 型更加有利。多重比较分析发现,G1272611A 位点的不同基因型中,GG 型个体的5 个周龄冠高均显著高于GA 型个体。

表5 GH 基因 A1270769G、G1272611A 位点与宁都黄鸡冠高(mm)性状的相关性Table 5 Correlation between A1270769G,G1272611A in GH gene and comb height (mm) of Ningdu yellow chickens
2.4 C1271211T、G1272611A 位点不同基因型与宁都黄鸡冠长性状的相关性
分析宁都黄鸡公鸡GH基因C1271211T、G1272611A 两个位点与7 个周龄冠长性状的相关性,并计算其加性效应和显性效应值(表6)。多重比较分析发现,C1271211T 位点的不同基因型中,CC 型个体的4、6、8、10、12、16 周龄冠长均显著高于CT 和TT 型个体,14 周龄冠长的CC 型个体只显著高于CT 型个体;从该位点对于4、6、8、10、12、14、16 周龄冠长的作用方式看,该位点加性方式起作用,CC 型更加有利。G1272611A 位点的不同基因型中,GG型个体的8、10、12、14、16 周龄冠长显著高于GA 型个体。
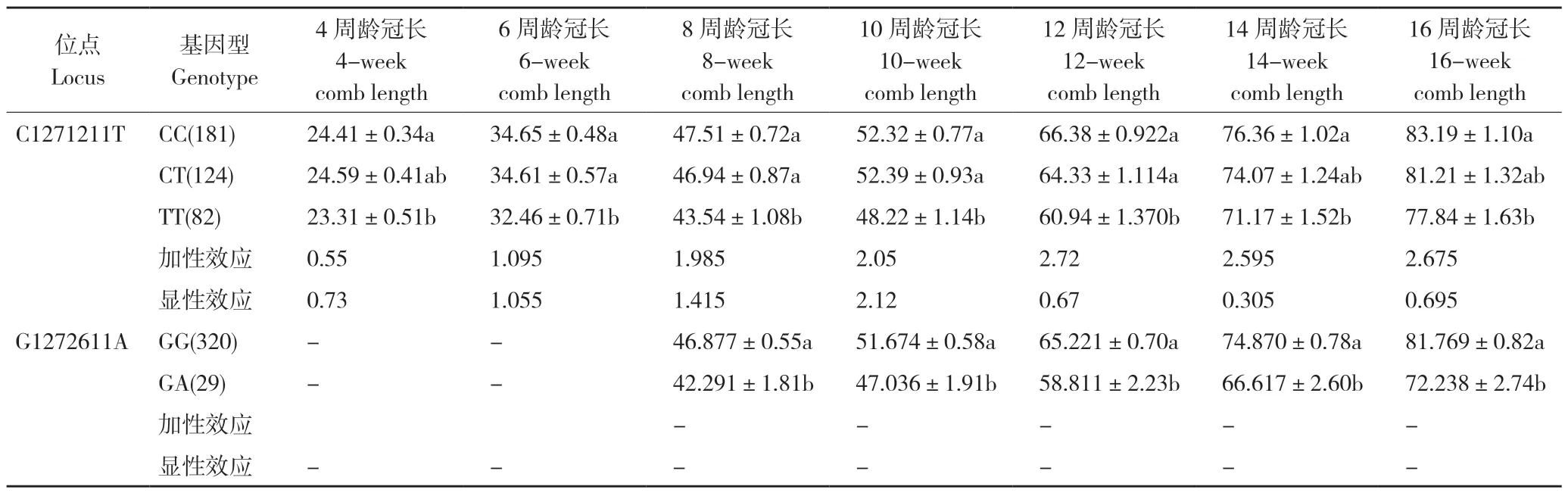
表6 GH 基因C1271211T、G1272611A 位点与宁都黄鸡冠长(mm)性状的相关性Table 6 Correlation between C1271211T and G1272611A in GH gene and comb length (mm) of Ningdu yellow chickens
3 讨论
本研究采用前期记录的宁都黄鸡公鸡生长早期(4~16 周龄)的冠高、冠长、冠厚、肉垂长、肉垂厚和冠齿数性状数据,与GH基因5'-侧翼区域和内含子1 区域的SNP 进行关联性分析,首先分析了与不同周龄鸡冠性状分别关联的SNP,发现较少位点与鸡冠厚、肉垂长和肉垂厚存在关联性,而与冠长和冠高关联位点较多,推测GH基因主要调控鸡冠的长度和高度发育;进一步选取与5 个连续周龄冠长或冠高性状指标均关联且与16 周龄显著关联的位点进行不同基因型多重比较分析,发现3 个位点对应的优势基因型分别为A1270769G 位点GG 基因型、C1271211T 位点CC 基因型、G1272611A 位点GG 基因型。经过Bonferroni 法校正后,A1270769G 与4 周龄冠高、G1272611A 与16 周龄冠长相关性达到显著水平。Hardy-weinberg 检验结果表明,A1270769G、C1271211T 均处于平衡状态,而G1272611A 未达到平衡状态。综上,A1270769G 可为鸡冠性状的潜在关键候选分子标记。
本课题组前期研究时发现,G1272611A 位点与16 周龄肉鸡的睾丸性状存在显著关联[3],表明位点G1272611A 既与鸡冠性状又与睾丸性状显著关联,推测该位点为调控鸡冠性状和睾丸性状的功能位点,对该位点不同基因型对应结合转录因子类型进行分析发现,GG 基因型对应的转录因子主要有p53、myogenin,AA 基因型对应的转录因子主要为En-1、C/EBPalpha[21],鸡冠性状的优势基因型为GG,推测转录因子p53、myogenin 更有利于鸡冠发育通路的激活,这些位点的调控功能还有待进一步验证。综合分析,G1272611A 可作为地方鸡繁殖性状选育的重要候选标记,可进一步开展大群体验证。本研究还发现,C1272225T 与12、14、16 周龄的冠高和冠长均显著相关,且该位点与16 周龄冠高的相关性经过Bonferroni 法校正后达到显著水平;C1271211T与6 个周龄的冠长显著关联,而这2 个位点也与16 周龄睾丸性状显著关联,说明这些位点变异可能影响GH基因表达,进而调控鸡冠和睾丸发育。
在家禽生产中,鸡冠性状是遗传育种选育中重要的表型性状,与重要的经济性状如开产性状、睾丸重等有密切联系[13,20]。鸡冠是家禽的第二性征,生长发育受遗传因素、激素及光照环境等影响。睾酮的分泌影响鸡冠和肉垂的发育[22],其与鸡冠表皮层和真皮层的受体结合影响鸡冠发育。鸡冠的的生长状况在一定程度上反映鸡的性功能是否优良[13,23-24]。韦金兑[25]发现清远麻鸡和广西麻鸡各冠型各日龄公鸡的冠高、冠长和肉垂长之间均呈较高程度正相关,相关系数在0.498~0.763 之间。肖俊[26]对清远麻公鸡性早熟进行研究,发现鸡冠和睾丸的生长发育有很大的关联性。周敏等[3]、王琨等[9]和姬杰菲等[27]的研究验证了性成熟前鸡冠与公鸡睾丸性状、母鸡开产性状具有较强的相关性。通过这些结果推测,以冠高或冠长为参数进行分子选育时可达到改良鸡繁殖性能性状的效果。而本研究发现的G1272611A 位点与冠高、冠长性状、睾丸性状均显著关联,进一步体现了鸡冠、睾丸性状的相关性,可能是因为鸡冠、睾丸的发育过程受到同一基因的调控。鸡冠是一种复杂性状,由多基因共同调控,目前已知鸡冠性状候选基因有AR、BMP2、CHADL、DRD1、EDN1、GARNL1、HSPA5、HSPA8、STK32A、PIK3R1、VIP1、VIP2和WNT9A等[12-17],GH基因序列变异关联鸡冠性状的挖掘将为解析鸡冠性状的多基因调控网络提供新的参考。
4 结论
GH基因可作为调控鸡冠长度和高度发育的重要候选基因,位于该基因5'-侧翼的G1272611A 位点及位于内含子1 区域的C1271211T、A1270769G位点与宁都黄鸡冠长或冠高性状显著关联。A1270769G、G1272611A 均 对8、10、12、14、16周龄冠高性状指标的影响达到显著水平(P< 0.05),这两个位点可作为冠高性状的分子辅助选育候选分子标记;G1272611A、C1271211T 均对8、10、12、14、16 周龄冠长性状的影响达到显著水平(P< 0.05),这两个位点可作为冠长性状的分子辅助选育候选分子标记。G1272611A 位点与5 个连续周龄冠长或冠高性状指标均关联,可作为调控鸡冠性状的候选分子标记。
猜你喜欢
小猕猴学习画刊·下半月(2023年4期)2023-06-20 19:29:14
科普童话·学霸日记(2021年3期)2021-09-05 16:42:08
作文大王·低年级(2021年6期)2021-08-02 18:04:24
辽金历史与考古(2021年0期)2021-07-29 01:06:34
作文小学中年级(2019年10期)2019-11-04 00:39:52
岭南音乐(2019年5期)2019-10-29 11:37:28
幼儿100(2017年13期)2017-06-15 20:29:45
东方少年·快乐文学(2016年7期)2016-08-01 19:18:06
军事历史(1999年4期)1999-08-20 07:36:02
军事历史(1991年6期)1991-08-16 02:22:02
