
四川凉山地区多份荞麦种子萌发抗寒性评价*
2023-09-19 02:08:46朱明昆方忠艳陈钰楠杜含梅王安虎周永红吴丹丹
中国农业气象 2023年9期
郑 文,朱明昆,方忠艳,陈钰楠,杜含梅,王安虎,周永红,2,吴丹丹,2**
四川凉山地区多份荞麦种子萌发抗寒性评价*
郑 文1,朱明昆1,方忠艳1,陈钰楠4,杜含梅3,王安虎3,周永红1,2,吴丹丹1,2**
(1.四川农业大学小麦研究所,成都 611130;2.西南作物基因资源发掘与利用国家重点实验室,成都 611130;3.西昌学院攀西特色作物研究与利用四川省重点实验室,西昌 615000;4.四川农业大学农学院,成都 611130)
荞麦是四川凉山等高海拔地区的主要粮食作物,为探究荞麦的抗寒能力,筛选适应凉山州极端气候的优异荞麦种质资源,以该地区推广和种植面积最大的5份甜荞(Moench.)和13份苦荞[(L.) Gaertn.]为材料,在4℃低温条件和22℃常温对照下进行种子萌发期抗寒性评价,测定胚根长、发芽率和发芽势等7项种子萌发生长指标,结合隶属函数分析、主成分分析和聚类分析对其抗寒性进行综合评价。结果表明:低温条件下,所有荞麦品种种子发芽延迟、发芽速度减缓;相比于苦荞,甜荞的7项萌发生长指标的平均相对值都保持较高水平,表明甜荞更有利于应对低温环境;隶属函数综合评价得出,荞麦各品种萌发期耐低温能力最强的为九寨沟栽培甜荞,其次为品甜3号,而对低温最为敏感的则是西荞6号。
凉山;荞麦;低温萌发;抗寒性;综合评价
低温是重要的环境胁迫因子之一,对植物的生长发育、空间分布和作物生产力都有不利影响。在植物萌发及幼苗期,低温会导致发芽率降低、生长缓慢、叶面积减小、植株黄化并伴随生长发育受损而导致坏死[1]。低温胁迫抑制植物细胞内光合作用、呼吸作用和生化途径酶的活性,打破植物细胞内活性氧清除机制和激素平衡,最终导致细胞的死亡以及作物减产[2]。近年,全球变暖放缓,气候变化进入新的停滞期,气温变化异常[3]。极端气候事件频繁出现,低温冷害冻害日趋严重,大量作物生长受到影响。研究表明玉米、大豆、棉花、香蕉和番茄等在低温下(10~15℃)均会出现冻害而导致减产[4-5]。低温已成为作物生产和农业发展面临的严峻挑战[6-7]。
荞麦属于蓼科(Polygonaceae)荞麦属(Mill.),又称花麦、三角麦,一年生或多年生草本或半灌木植物[8]。从地理分布来看,以四川、云南、贵州和西藏为主的中国西南地区是世界公认的荞麦属植物起源中心、分布中心和多样化中心。甜荞(.)和苦荞(.)作为栽培种,在世界范围内广泛种植,尤其在东亚和东欧的高海拔地区[9]。甜荞常栽培于西北、西南等地平原或者低海拔山区农田,海拔为0−1000m,而苦荞主要栽培于西南山区农田,海拔为500−3900m。
近年来,由于越来越多的人采用无麸质饮食来对抗麸质不耐受症状,荞麦引起了全世界的关注[10]。荞麦在中国栽培历史悠久,至今仍是华北、西北、西南以及一些高寒地区、少数民族聚居区、边远山区不可替代的作物,并一直作为中国西南凉山地区的主要粮食作物[11]。研究表明在荞麦的生长发育阶段,特别是在发育和开花的早期阶段,对环境条件要求高,尤其对霜冻高度敏感[12]。据统计调查,凉山州自2018年以来极端低温天气频发,在苦荞出苗关键期3−4月,极端低温多次降至4℃以下,2021年甚至跌至−9℃,严重影响了苦荞的存活及生长发育。了解荞麦对低温的耐受性意义重大,但目前关于荞麦的低温研究报道较少,特别是针对萌发期的低温抗性研究还很缺失。
本研究选取四川省凉山州推广和种植面积最大的5份甜荞和13份苦荞种子共计18份材料,进行种子萌发期4℃低温处理与常温对照处理试验,对其进行多项生长指标的测定并结合隶属函数分析、主成分分析和聚类分析对其抗寒性进行综合评价,旨在筛选抗寒性强的荞麦种质,为抗寒品种的选育提供基础材料。
1 材料与方法
1.1 试验材料
荞麦种子信息如表1所示。试验用种子均由西昌学院王安虎教授提供,包括5份甜荞和13份苦荞,其中甜荞包括2份育成种和3份地方种。苦荞均为育成种,包括川荞系列(4份)、西荞系列(8份)和九江苦荞(1份)。
1.2 试验方法
对18份荞麦材料进行低温处理。选取籽粒饱满的种子用15%的NaClO消毒5min,用超纯水冲洗5次,挑选30粒种子放于铺有湿润的两层双圈定性滤纸的培养皿中,将培养皿分别放入4℃低温(荞麦生长临界温度)培养箱和22℃常温(对照)培养箱中,光照周期均为每日6:00-22:00以14000Lx的光照,湿度均为60%,共培养10d。所有品种分别取10颗种子放于培养皿中进行萌发期试验,每个品种3次重复。
1.3 观测项目
每日20:00-21:00进行数据观测并统计种子的萌发情况,以胚根长度达到种子长度的一半(约0.5cm)作为萌发标准。第10天萌发结束后,每个处理挑选出10颗长势一致的发芽种子,用游标卡尺进行胚根长RL(Root length)的测定。同时用精度为0.0001g的电子天平称量其鲜重FW(Fresh weight),然后放入105℃烘箱中杀青0.5h后75℃烘干3d至恒重,再次用精度为0.0001g的电子天平称量其干重DW(Dry weight)。
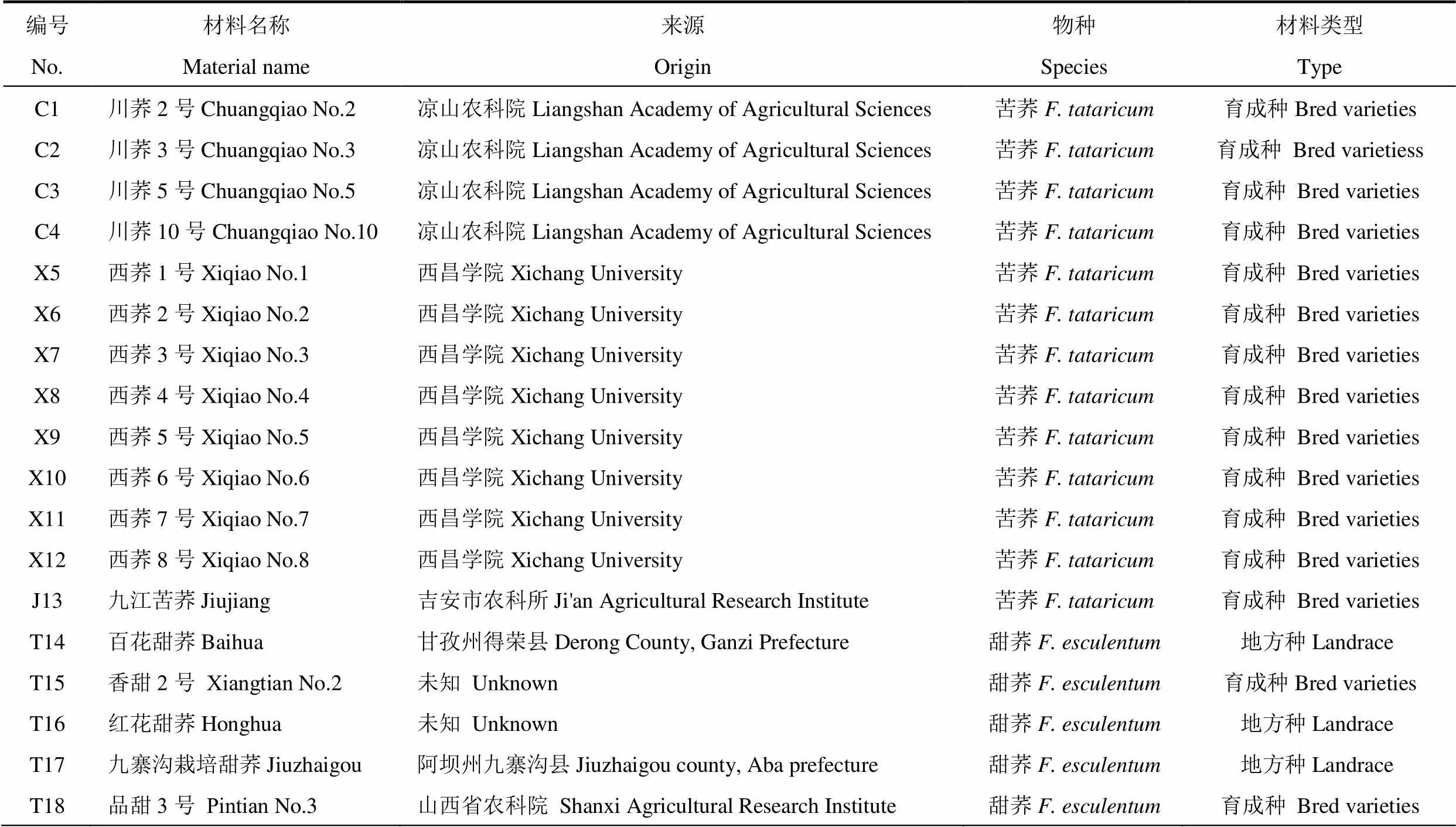
表1 参试荞麦种子信息
1.4 种子发芽指标计算
参考文献[13−14]计算种子发芽率(GR)、发芽势(GE)、发芽指数(GI)、活力指数(VI),计算公式分别为
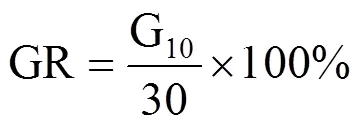
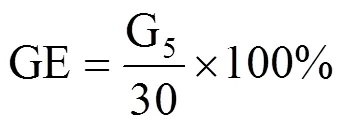
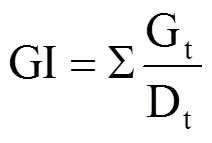
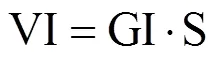
式中,Gt为在t天内的发芽数(个),30为供试种子数,Dt为相应的发芽天数(d),S为胚根长(cm)。GR为10d内的发芽率,GE为5d内的发芽势,GI为发芽指数,VI为活力指数,均为无量纲。
1.5 品种抗寒能力综合评价值计算[15]
1.5.1 抗寒系数
为消除材料不同造成的差异,首先将各指标进行相对化处理,用抗寒系数表示。RRL为胚根长的抗寒系数,RFW为鲜重指标的抗寒系数,RDW为干重指标的抗寒系数,RGR为发芽率的抗寒系数,RGE为发芽势的抗寒系数,RGI为发芽指数的抗寒系数,RVI为活力指数的抗寒系数。其通用计算公式为
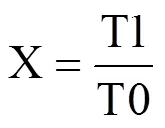
式中,X为各指标的抗寒系数,T1为该指标在低温胁迫下的测定值,T0为该指标在对照处理中的测定值。
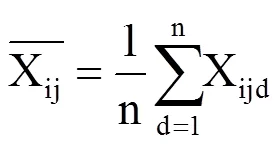
1.5.2 综合指标得分
利用各品种计算的各指标平均抗寒系数及其权重计算品种的综合抗寒指标。即
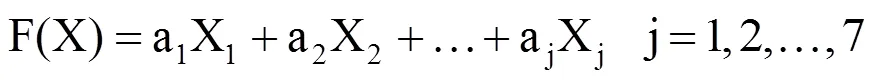
式中,F(X)某品种抗寒指标综合得分。aj为指标权重,用主成分分析法确定。Xj为每个指标经标准化处理后的值。
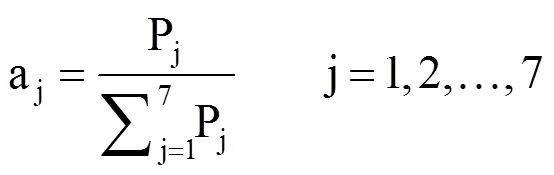
式中,aj表示第j个指标的权重系数,Pj为主成分分析中第j个指标的综合贡献率。
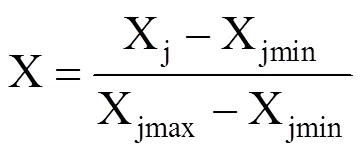
式中,Xjmin为第j个指标的最小值,Xjmax为第j个指标的最大值。
则不同品种荞麦的抗寒能力综合评价值(D)可表述为
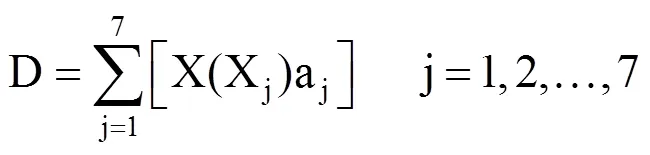
1.6 数据处理
数据用Excel整理分析,利用SPSS 26.0进行相关性分析、主成分分析和聚类分析,Origin 2019b进行绘图。
2 结果与分析
2.1 低温条件下荞麦种子发芽速度分析
由图1可见,与常温培养箱(对照,22℃)相比,低温条件下(4℃)所有(18个)荞麦品种的种子萌发速度都受到影响,但低温条件对不同材料发芽速度的影响程度不同。低温下,川荞系列(图1a-d)发芽集中在第6−8天,在第7−10天种子发芽基本完成;西荞系列(图1e-l)发芽集中在第5−8天,在第7−10天种子发芽基本完成;甜荞系列(图1n-r)发芽集中在第3−6天,在第6−10天种子发芽基本完成;九江苦荞(图1m)发芽集中在第6−7天,于第10天完成全部发芽。总体表现为前期发芽率低、后期发芽率逐渐升高的特点,至第10天发芽率达到与CK同样水平,说明低温条件下种子发芽延迟、发芽速度减缓。
2.2 低温条件下荞麦种子活力指标分析
由图2可见,在低温条件下,18份荞麦材料中,甜荞系列的平均相对胚根长、平均相对鲜重、平均相对干重、平均相对发芽率、平均相对发芽势、平均相对发芽指数和平均相对活力指数7项指标均高于苦荞系列。所有供试材料在低温条件下,上述7项指标除了平均相对发芽率以外,其余指标的绝对值均小于1,说明低温条件下所有品种种子的活力指标降低,反映了低温对荞麦种子的萌发产生了不利影响。
甜荞系列品种平均相对胚根长、平均相对鲜重、平均相对发芽势、平均相对发芽指数和平均相对活力指数这5项种子活力指标数值高于川荞系列、西荞系列以及九江苦荞。平均相对胚根长中,甜荞系列的T17(九寨沟栽培甜荞)最高(0.882),而西荞系列的X8(西荞4号)最低(0.112)。平均相对鲜重中,甜荞系列的T15(香甜2号)最高(0.586),而T16(红花甜荞)最低(0.238)。平均相对干重中,川荞系列的C4(川荞10号)最高(0.318),而西荞系列的X7(西荞3号)最低(0.105)。平均相对发芽率中,西荞系列的X9(西荞5号)最高(1.2),而川荞系列的C3(川荞5号)最低(0.833)。平均相对发芽势指标中,甜荞系列的T17(九寨沟栽培甜荞)最高(0.95),而最低水平在4个荞麦系列中均有包括,其数值为0。平均相对发芽指数中,甜荞系列的T16(红花甜荞)最高(0.499),而西荞系列的X7(西荞3号)最低(0.195)。在平均相对活力指数这项指标中,甜荞系列的T17(九寨沟栽培甜荞)最高(0.371),而西荞系列的X10(西荞6号)最低(0.027)。由上可见,在低温条件下甜荞系列品种比苦荞更能应对低温环境的不利影响。
2.3 荞麦种子萌发期抗寒性综合评价
2.3.1 指标筛选
对用于抗寒性评价的7项生长指标进行相关性分析,结果见表2。由表可知,达到显著水平的有平均相对鲜重与平均相对胚根长和平均相对活力指数(P<0.05);达到极显著水平的有平均相对发芽势与平均相对胚根长,平均相对发芽指数与平均相对发芽率和平均相对发芽势,平均相对活力指数与平均相对胚根长、平均相对发芽势和平均相对发芽指数(P<0.01)。其中,平均相对发芽势与平均相对活力指数的相关性最高(R=0.874)。说明这些指标可以作为荞麦萌发期抗寒性初步筛选的指标。
2.3.2 指标权重确定
利用主成分分析法确定各指标权重。主成分分析可将多个指标转化为几个独立的指标,从而反映原来多指标的信息。将7个指标耐低温系数进行主成分分析后,得到两个主成分,如表3所示。第一主成分的特征根为3.727,贡献率为53.249%,发芽势、发芽指数和活力指数有较高的载荷,反映了荞麦的萌发情况,所以第一主成分可归纳为“萌发因子”;第二主成分特征根为1.535,贡献率为21.929%,根长有较高的载荷(0.583),反映了荞麦的根部生长状况,可概括为“根部生长因子”。
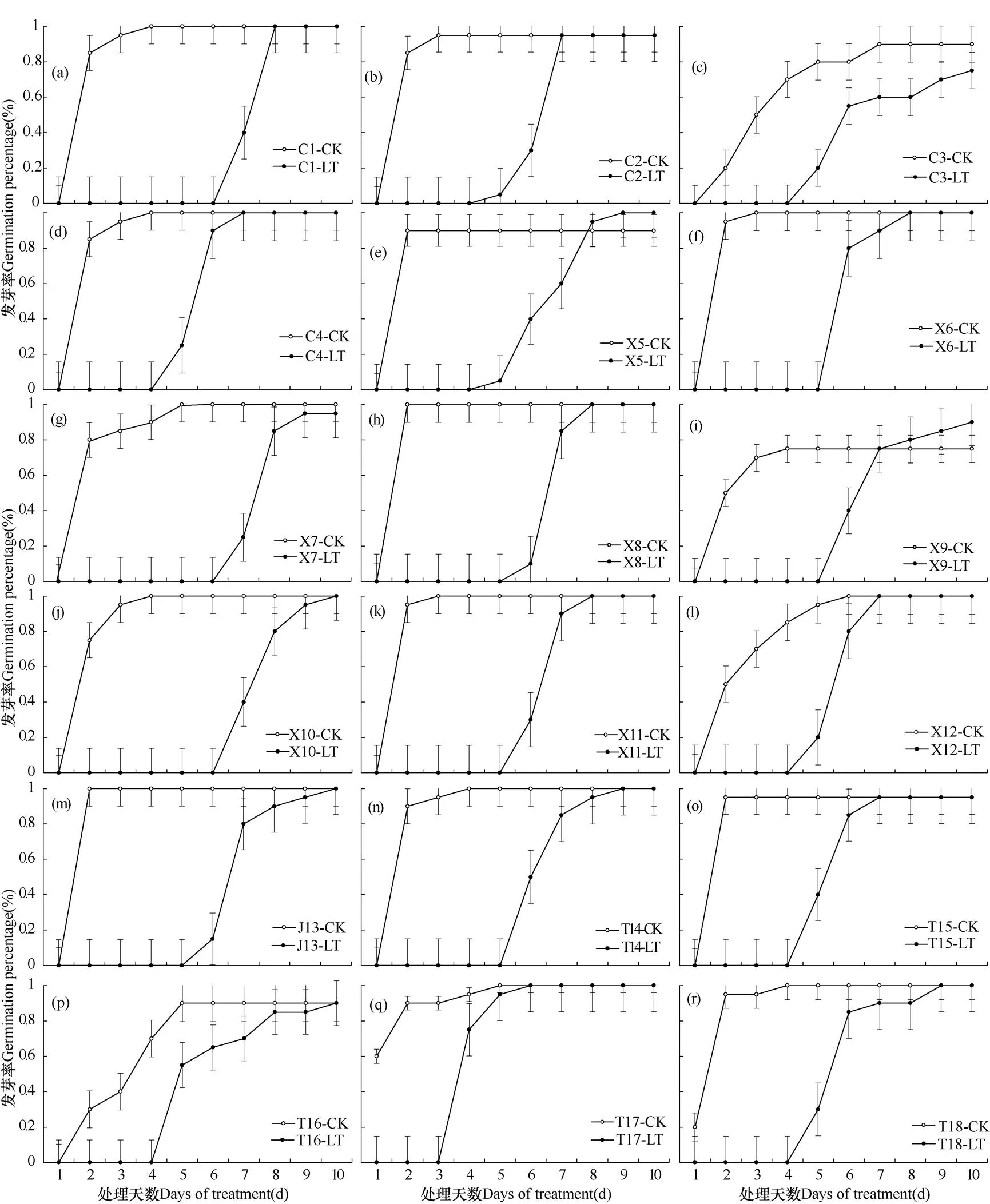
图1 试验期间(10d)不同荞麦材料在常温(22℃,CK)和低温(4℃,LT)条件下发芽率变化过程
注:竖线代表标准误差。
Note: The vertical bars indicate root mean squared error.
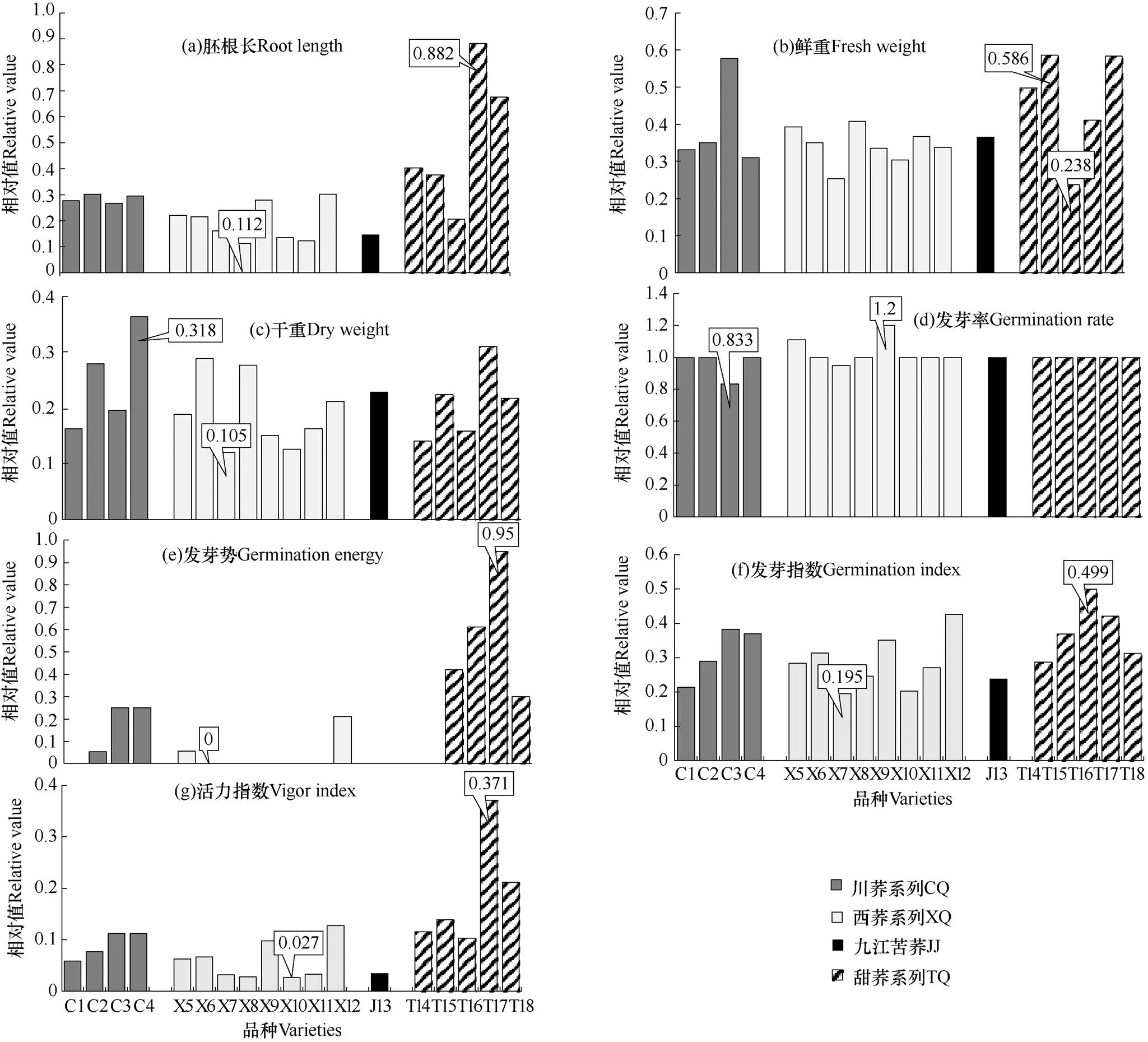
图2 低温条件下不同荞麦材料7个发芽活力指标与CK(22℃)对比的相对值
注:川荞系列(CQ)包括C1−C4,西荞系列(XQ)包括X5−X12,九江苦荞(JJ)包括J13,甜荞(TQ)包括T14−T18。
Note: Chuanqiao series (CQ) includes C1−C4, Xiqiao series (XQ) includes X5−X2, Jiujiang tartary buckwheat (JJ) includes J13, and common buckwheat (TQ) includes T14−T18.
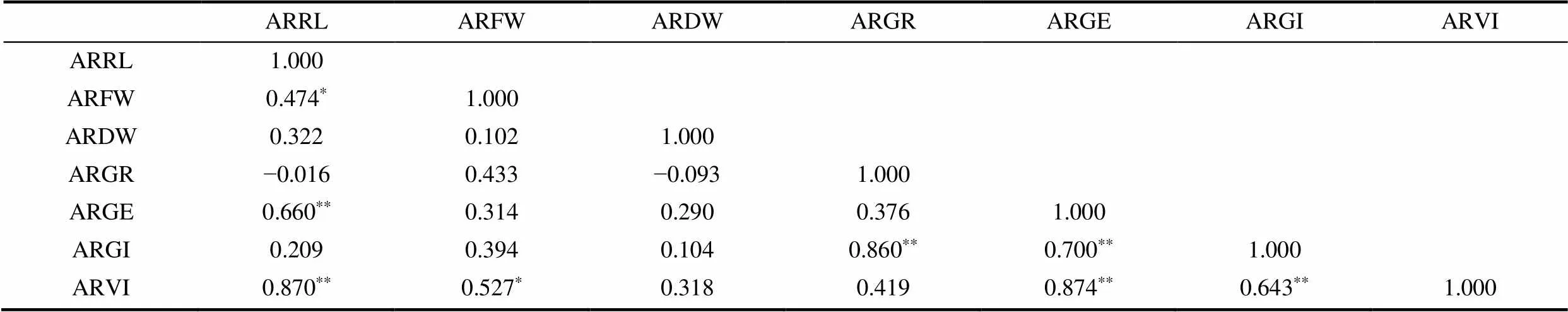
表2 荞麦种子7个抗寒指标间的相关性分析
注:*P < 0.05,**P < 0.01。ARRL为平均相对胚根长,ARFW为平均相对鲜重,ARDW为平均相对干重,ARGR为平均相对发芽率,ARGE为平均相对发芽势,ARGI为平均相对发芽指数,ARVI为平均相对活力指数。下同。
Note:*is P < 0.05,**is P < 0.01. ARRL is average relative root length, ARFW is average relative fresh weight, ARDW is average relative dry weight, ARGR is average relative germination rate, ARGE is average relative germination energy, ARGI is average relative germination index, ARVI is average relative vigor index. The same as below.
2.3.3 综合指标计算
因子载荷值除以该成分的特征根的平方根得到特征向量,将7个指标的平均抗寒系数进行数据标准化后,利用式(7)得到两个主成分的综合指标值。即
F1=0.724ARRL+0.636ARFW+0.324ARDW+0.606ARGR
+0.878ARGE+0.801ARGI+0.955ARVI(11)
F2=0.583ARRL-0.078ARFW+0.559ARDW-0.747ARGR
+0.124ARGE-0.510ARGI+0.208ARVI (12)
式中,ARRL为平均相对胚根长,ARFW为平均相对鲜重,ARDW为平均相对干重,ARGR为平均相对发芽率,ARGE为平均相对发芽势,ARGI为平均相对发芽指数,ARVI为平均相对活力指数。均为无量纲。
根据式(8)和式(9)得到的权重系数与隶属函数值μ,再按照式(10)得到各低温处理的综合评价值(D),对各品种抗寒能力进行综合评价,表4显示,D值的变化范围在0.064~1.000,其中甜荞系列的T17(九寨沟栽培甜荞)的D值最大,该品种抗寒性最强,而西荞系列的X10(西荞6号)的抗寒性最弱。

表3 种子萌发期抗寒性指标的主成分分析

表4 不同荞麦材料的抗低温处理综合评价值(D)
2.3.4 综合抗寒能力聚类分析
采用欧式距离法对18份荞麦材料的抗寒能力综合评价值(D值)进行系统聚类分析,结果如图3所示。在欧式距离5.0处,18份材料可以分为4大类:第I类材料低温耐性最强,为T17(九寨沟栽培甜荞);第Ⅱ类为耐低温较强的两份甜荞材料,即T18(品甜3号)和T15(香甜2号);第III类为中等耐低温材料,包括甜荞系列的T16(红花甜荞)、T14(百花甜荞),以及川荞系列的C3(川荞5号)、C4(川荞10号)和西荞系列的X12(西荞8号);第Ⅳ类低温敏感材料,包括有川荞系列、西荞系列以及九江苦荞在内的10份材料。因此,甜荞的耐低温能力普遍强于苦荞,苦荞系列中,川荞的整体低温耐性强于西荞。

图3 不同荞麦材料抗寒能力综合评价值(D值)的聚类分析
3 结论与讨论
3.1 讨论
适宜的温度是种子萌发最重要的环境因子之一,营养物质的分解和其他一系列的生理生化活动,都需要在适宜的环境下才能进行[16]。本研究中,低温条件下荞麦种子发芽延迟,发芽速度减缓,这与彭运翔等[17]的研究结果一致,这可能是因为种子呼吸作用及种子内含物的分解代谢和生理活动受到了低温抑制[18]。本研究对用于抗寒性评价的7项生长指标进行相关性分析,结果表明,平均相对发芽势和平均相对活力指数相关性最高。发芽势可直观表现荞麦种子的发芽情况,活力指数可表现种子的发芽质量和活力,这是衡量植物抗寒能力的两个重要指标,这与王晓龙等[19]研究结果一致,低温条件下,不同品种的荞麦种子的发芽势和活力指数均受到不同程度的降低,这可能是因为低温对种子内部酶活性、可溶性糖和淀粉等生理代谢过程起到了抑制作用[20],导致种子活性降低。本研究利用主成分分析得出两个主成分,将其归纳为发芽势和发芽指数载荷更高的“萌发因子”和胚根长更高载荷的“根部生长因子”。胚根是植物从土壤中吸收水分的主要器官,同时也是贮存营养物质的重要部位,所以也用胚根长来衡量种子的萌发情况,这与侯龙鱼等[21]研究结果一致,低温条件下,荞麦根长生长受到抑制,可能是由于荞麦种子内部营养物质分解、转化和一系列代谢活动受到影响[22]。
植物的抗寒性是由多个因子控制的数量性状,不同物种或种质材料对不同指标的响应规律不同,为全面评价植物的抗寒性强弱,许多学者采用综合多个指标的隶属函数法,以消除用单一指标鉴定的片面性。贾祥等[23]通过对4份豆科牧草材料发芽率和发芽势等6个生长指标的隶属函数综合分析,评价了其抗寒性,筛选出紫花苜蓿耐寒性最高,刘伟等[24]利用隶属函数法对3份辣椒材料的耐寒性进行了综合评价,结果为紫杆美早佳>强龙>长辣香妃。本研究则主要通过隶属函数综合评价18份荞麦材料萌发期抗寒性,初步判定甜荞的耐低温能力优于苦荞。在低温胁迫下,甜荞材料除百花甜荞外,其余甜荞材料的发芽势相较于苦荞材料都处于较高水平,表明甜荞种子对于低温抑制的适应能力较强。其原因可能是为甜荞主产区大多在陕西、山西一带,处于秦岭以北,入冬时间早、冬季长且气温低。甜荞主要栽培于海拔1000m以下的平原或是山区农田[25],栽培条件适宜,吸引了许多农户和育种家的兴趣。在农户和育种家的选育条件下,研究所用的甜荞经过长期低温驯化,表现出了更为耐低温的特性。而苦荞的主产区在四川、云南一带的高海拔山区,未经过漫长的寒冬驯化,在抗寒方面明显逊色于甜荞。九寨沟栽培甜荞和红花甜荞的发芽指数最高,表明在低温胁迫下,其种子活力依旧保有较高水平,受低温胁迫的影响比大多数苦荞材料小,说明本研究评价的甜荞材料的低温耐受能力优于苦荞材料。若要对甜荞和苦荞的抗寒性做出更全面的评价,需要收集大量的农家种和当地野生种做出系统性评价。
3.2 结论
本研究对荞麦在低温条件下的根长、发芽率和发芽指数等7项指标进行主成分分析,发现发芽势、发芽指数和活力指数作为“萌发因子”反映了荞麦的萌发情况,根长作为“根部生长因子”反映了荞麦的根部生长状况。采用隶属函数法,对7个指标进行抗寒性综合评价,结果表明,九寨沟栽培甜荞的抗寒性最高,品甜3号次之,西荞6号的抗寒性最弱,受低温的抑制作用最为明显。
[1] Manasa S L,Panigrahy M,Panigrahi K,et al.Overview of cold stress regulation in plants[J].The Botanical Review, 2021(1):1-29.
[2] Ritonga F N,Chen S.Physiological and molecular mechanism involved in cold stress tolerance in plants[J]. Plants,2020,9(5):560.
[3] Johnson N C,Xie S P,Kosaka Y,et al.Increasing occurrence of cold and warm extremes during the recent global warming slowdown[J].Nature Communications,2018,9(1): 1724.
[4] Wang L J,Zhang P,Wang R N,et al.Effects of variety and chemical regulators on cold tolerance during maize germination[J].Journal of Integrative Agriculture,2018,17 (12):2662-2669.
[5] Senda M,Kawasaki M,Hiraoka M,et al.Occurrence and tolerance mechanisms of seed cracking under low temperatures in soybean()[J].Planta,2018,248 (2):369-379.
[6] 张瑶,杨再强,姜雨函,等.低温寡照条件下黄瓜叶片光合作用减弱的机理分析[J].中国农业气象,2022,43(4): 285- 294.
Zhang Y,Yang Z Q,Jiang Y H,et al.Mechanism analysis on photosynthetic attenuation in cucumber leaves under low temperature and weak light condition[J].Chinese Journal of Agrometeorology,2022,43(4):285-294.(in Chinese)
[7] 吉春容,刘海蓉,罗继,等.低温冻害过程中枣树枝条相关生理指标的变化[J].中国农业气象,2016,37(5):545-554.
Ji C R,Liu H R,Luo J,et al.Changes of physiological indices of jujube branches under process of low temperature and freeze injury[J].Chinese Journal of Agrometeorology,2016, 37(5):545-554.(in Chinese)
[8] 唐宇,邵继荣,周美亮.中国荞麦属植物分类学的修订[J].植物遗传资源学报,2019,20(3):646-653.
Tang Y,Shao J R,Zhou M L.A taxonomic revision ofMill from China[J].Journal of Plant Genetic Resources, 2019,20(3):646-653.(in Chinese)
[9] 李玲,闫旭宇,王延峰.荞麦文献分析及相关进展研究[J].科技通报,2020,36(1):1-7.
Li L,Yan X Y,Wang Y F.Analysis of literature and related research progress of buckwheat[J].Bulletin of Science and Technology,2020,36(1):1-7.(in Chinese)
[10] Zhang L,Li X,Ma B,et al.The tartary buckwheat genome provides insights into rutin biosynthesis and abiotic stress tolerance[J].Molecular Plant,2017,10(9):1224-1237.
[11] 吴朝昕,陈庆富,黄娟,等.不同生态区对不同品种甜荞和苦荞农艺性状和品质性状的影响[J].安徽农业大学学报,2020,47(4):618-626.
Wu C X,Chen X F,Huang J,et al.Effects of different ecological regions on agronomic and quality characters of different varieties of common buckwheat and tartary buckwheat[J].Journal of Anhui Agricultural University, 2020,47(4):618-626.(in Chinese)
[12] Jeon J,Kim J K,Wu Q,et al.Effects of cold stress on transcripts and metabolites in tartary buckwheat ()[J].Environmental and Experimental Botany,2018,155:488-496.
[13] Shun D L,Bian N F,Wang X,et al.Comprehensive evaluation of cold tolerance and germplasm screening of high oleic acid peanut at germination stage[J].Journal of Nuclear Agricultural Sciences,2021,35(6):1263-1272.
[14] Du L L,Bo Y Z,Hong M S,et al.Comprehensive evaluation on cold resistance of early fruiting walnut cultivars[J].Acta Horticulturae Sinica,2015,42(3):545-553.
[15] 徐超,高芮,王明田,等.苗期高温对草莓果实营养品质影响的模糊综合评价及模型建立[J].中国农业气象,2020, 41(12):785-793.
Xu C,Gao R,Wang M T,et al.Fuzzy comprehensive evaluation and model establishment of the effect of high temperature in the seedling stage on the nutritional quality of strawberry fruit[J].Chinese Journal of Agrometeorology, 2020,41(12):785-793.(in Chinese)
[16] 曹慧明,史作民,周晓波,等.植物对低温环境的响应及其抗寒性研究综述[J].中国农业气象,2010,31(2):310-319.
Cao H M,Shi Z M,Zhou X B,et al.A review on response of plant to low temperature and its cold resistance[J].Chinese Journal of Agrometeorology,,2010,31(2):310-319.(in Chinese)
[17] 彭运翔,张力君,陈立波.18个苜蓿品系种子萌发期对温度的响应[J].内蒙古草业,2012,24(2):42-43.
PengY X,Zhang L J,Chen L B.Response of 18 alfalfa strains to temperature during seed germination[J].Inner Mongolia Prataculture,2012,24(2):42-43.(in Chinese)
[18] Hao X,Wang L,Zeng J,et al.Response and adaptation mechanisms of tea plant to low-temperature stress[M]//Han W Y,Li X,Ahammed G J.Stress physiology of tea in the face of climate change.Singapore:Springer Singapore,2018: 39-61.
[19] 王晓龙,杨曌,李红,等.6个苜蓿品种种子萌发对低温的响应[J].中国草地学报,2022,44(7):79-86.
Wang X L,Yang Z,Li H,et al.Response of seed germination of six medicago sativa varieties to low temperature[J]. Chinese Journal of Grasland,2022,44(7):79-86.(in Chinese)
[20] Wang Y X,Hu Y,Chen B H,et al.Physiological mechanisms of resistance to cold stress associated with 10 elite apple rootstocks[J].Journal of Integrative Agriculture,2018,17(4): 857-866.
[21] 侯龙鱼,任立飞,任丽昀,等.紫花苜蓿低温萌发特征及指标筛选[J].西南民族大学学报(自然科学版),2019,45(6): 564-569.
Hou L Y,Ren L F,Ren L Y,et al.Efects of low temperature on seed germination characteristics of alfalfa ()[J].Journal of Southwest Minzu University(Natural Science Edition),2019,45(6):564-569.(in Chinese)
[22] Theocharis A,Clément C,Barka E A.Physiological and molecular changes in plants grown at low temperatures [J].Planta,2012,235(6):1091-1105.
[23] 贾祥,陈盼盼,王艳琳,等.4种豆科牧草种子萌发期对低温胁迫的响应[J].高原农业,2020,4(2):137-142.
Jia X,Chen P P,Wang Y L,et al.Response of four legume species to cold stress during seed germination[J].Journal of Plateau Agriculture,2020,4(2):137-142.(in Chinese)
[24] 刘伟,黄勇,胡展育,等.低温对辣椒种子萌发的影响及其抗寒性分析[J].江西农业学报,2020,32(10):68-71.
Liu W,Huang Y,Hu Z Y,et al.Effect of low temperature on germination of capsicum seeds and analysis of cold resistance[J].Acta Agriculturae Jiangxi,2020,32(10):68-71. (in Chinese)
[25] Suvorova G,Zhou M.Distribution of cultivated buckwheat resources in the world[M]//Zhou M, Kreft I, Suvorova G,et al.Buckwheat germplasm in the world.New York:Academic Press, 2018:21-35.
Evaluation on Cold Resistance of Buckwheat Germplasm during Germination Stage in Sichuan Province
ZHENG Wen1, ZHU Ming-kun1, FANG Zhong-yan1, CHEN Yu-nan4, DU Han-mei3, WANG An-hu3, ZHOUYong-hong1,2, WU Dan-dan1,2
(1. Sichuan Agricultural University Triticeae Research Institute, Chengdu 611130, China;2. State Key Laboratory of Crop Gene Exploration an Utilization in Southwest China,Chengdu 611130;3. Panxi Crops Research and Utilization Key Laboratory of Sichuan Province, Xichang University, Xichang 615000;4. College of Agronomy, Sichuan Agricultural University, Chengdu 611130)
Buckwheat is the major food crop in high altitude areas such as Liangshan district, Sichuan province, China. In order to explore the cold resistance of buckwheat and screen excellent buckwheat germplasm resources adapted to Liangshan district, 5 common buckwheat (Moench.) and 13 Tartary buckwheat [(L.) Gaertn. ], the most extensive varieties and landrace, were used to evaluate the cold resistance during seed germination stage at 4℃(low temperature condition) and 22℃(control condition). Seven indices related to germination and growth were measured, including radicle length, germination rate and germination potential. Comprehensive cold resistance was reflected by subordinate function analysis, principal component analysis and cluster analysis. The results showed that under low temperature conditions, the seed germination of all buckwheat materials was delayed and the germination rate was retarded. Compared withTartary buckwheat, seven germination and growth indexes of common buckwheat maintained a higher level, indicating that common buckwheat was more tolerant under low temperature environment. The comprehensive evaluation of the membership function results suggested that Jiuzhaigou common buckwheat had the strongest low temperature tolerance ability at the germination stage, followed by Pintian No.3, whereas Xiqiao No.6 was the most sensitive to low temperature.
Liangshan; Buckwheat; Low temperature germination; Cold resistance; Comprehensive evaluation
10.3969/j.issn.1000-6362.2023.09.004
郑文,朱明昆,方忠艳,等.四川凉山地区多份荞麦种子萌发抗寒性评价[J].中国农业气象,2023,44(9):795-804
2022−10−25
攀西特色作物研究与利用四川省重点实验室资助课题(SZKF2201);2021−2023年中国农技协西昌苦荞麦科技小院专项基金;四川省麦类作物创新团队项目(2019scyz02)
吴丹丹,博士,讲师,主要从事植物资源分类与评价工作,E-mail:wudandan@sicau.edu.cn
郑文,E-mail:wenzheng0621@hotmail.com
猜你喜欢
广西林业科学(2021年6期)2022-01-21 07:39:28
今日农业(2021年20期)2021-11-26 01:23:56
山西农业科学(2021年8期)2021-08-17 06:20:10
河北果树(2020年4期)2020-11-26 06:04:28
山西农业科学(2020年1期)2020-02-27 17:00:54
青年歌声(2018年5期)2018-10-29 03:18:40
广东饲料(2016年3期)2016-12-01 03:43:12
广西林业科学(2016年2期)2016-03-20 05:53:26
林业与生态(2016年2期)2016-02-27 14:24:01
当代畜禽养殖业(2014年1期)2014-02-27 07:58:34
