
川渝地区塔罗科血橙新系炭疽病的病原鉴定及其遗传多样性分析
2023-09-12 00:57黄仁军冯小华何锦辉
西南农业学报 2023年7期
陈 泉,羊 炼,黄仁军,冯小华,何锦辉
(1.重庆三峡农业科学院,重庆 万州 404155;2.重庆市万州区植物保护与果树技术推广站,重庆 万州 404155)
【研究意义】柑橘是世界第一大水果,中国柑橘种植面积和产量均居全球第一。塔罗科血橙新系(Citrussinensiscv.Tarocco)属晚熟甜橙类柑橘[1]。近年来,川渝两地大力发展塔罗科血橙新系,成为我国塔罗科血橙新系的主产区,种植规模达1.5×105hm2。柑橘炭疽病是世界性分布的真菌病害[2],遍布于高温多湿的热带和亚热带柑橘产区[3]。在我国,炭疽病在所有柑橘产区均有分布为害,在重庆、湖南、广东、四川等地的柑橘尤其是甜橙上发生严重[4]。炭疽病发病后,柑橘叶片有圆形、不规则形或“V”字形病斑,中央浅灰褐色或灰白色,病部散生或轮纹状排列黑色小粒点,多雨高湿时产生肉红色分生孢子,落叶严重,枝梢枯死,果实发病表现出干疤和泪痕,还会在采后导致全果腐烂[5-6]。条件适宜时,炭疽病危害柑橘果实后可造成减产90%。因柑橘炭疽病在嫩叶、枝梢、花器和果实等部位都能发生,且在柑橘的生长期、结果期、贮存期均可发病,因此较难防控[7],是限制川渝两地塔罗科血橙新系产业发展的重要因素。柑橘炭疽菌丰富的遗传多样性使菌种发生不同程度的遗传变异,其对各种杀菌剂的抗性不同,是导致川渝地区近年柑橘炭疽病危害严重的重要原因。【前人研究进展】较早报道柑橘炭疽病原为胶孢炭疽菌(ColletotrichumgloeosporioidesPenz)和尖孢炭疽菌(ColletotrichumacutatumSimmonds)[8-9],2个种可用种特异性引物CgInt/ITS4和CaInt/ITS4区分[10],随后首次发现了平头炭疽菌(Colletotrichumtruncatum)。胶孢炭疽菌分布在我国各个省市,是绝对优势种群,易引起柑橘采后贮藏期的炭疽病,还会危害叶片和枝梢[3]。在我国早期的报道中,尖孢炭疽菌仅在云南瑞丽的莱檬上有发现,引起莱檬炭疽,在其他柑橘类型上只引起花后落果[3,11]。随后在四川广安的龙安柚(Citrusgrandisvar.longanyou)上发现一个致病性很强的橘红色慢生型尖孢炭疽菌菌株,易引起采后果实腐烂[12]。平头炭疽菌分离自云南和广东,仅占总量的3.7%[3]。在与寄主植物长期斗争过程中,病原菌的群体结构普遍会发生分化[13],对不同药剂的抗药性也不同。炭疽菌群体遗传变异丰富,同一种的不同分离物之间差异很大[10]。目前,炭疽菌遗传分类多数采用多基因系统学结合形态学的方法[14],而ITS序列在炭疽菌属分子系统学中已得到广泛运用。例如李娜[15]、王雪莲[16]通过形态学和ITS序列分别将引起湖北和广东柑橘炭疽病的病原鉴定为胶孢炭疽菌。随着分子系统学研究的不断深入,人们发现ITS不能有效用于近似种的识别[17-18],因此,更多基因逐渐被引入到炭疽菌的分类中,有大量学者采用β-微管蛋白(TUB2)、谷氨酸合成酶(GS)、甘油醛-3-磷酸脱氢酶(GDPH)、钙调蛋白基因(CAL)、核糖体DNA内转录间隔区(ITS)、肌动蛋白基因(ACT)、几丁质合成酶基因(CHSΙ)及组蛋白基因(HIS3)等进行炭疽菌分类研究[19-23],并且发现部分菌株的遗传分化与寄主和地理来源无关[24-25]。Domm等[26]通过形态学与ITS、TUB2、GDPH基因序列分析,将116个代表菌株分属于7个类群和1个独立的种。Huang等[27]通过6个基因序列对中国柑橘属和金橘属植物上的312株炭疽菌进行分析研究,鉴定出6个种,其中2个新种。Weir等[18]也用同样的方法对400个胶孢炭疽复合种菌株进行分析,区分出 22个种,其中8个新种,同时证明GDPH基因是胶孢炭疽复合种遗传多样性分析的有效基因。陈国庆[3]对我国浙江、江西、广东、福建、湖南、重庆和云南等地的不同柑橘炭疽菌分析,发现GS、GDPH、Cal序列分析有效揭示了柑橘胶孢炭疽菌种下变异水平。谢桃明[28]将247株柑橘菌株用GDPH基因进行鉴定,将供试炭疽菌分为6大类。另外,在柿子、草莓、草本植物炭疽菌方面也有许多将形态学与多基因分子系统学结合进行遗传多样性研究的报道[29-31]。【本研究切入点】关于柑橘炭疽病已有较多研究报道,但针对川渝两地塔罗科血橙炭疽病的病原及其遗传多样性研究未见报道。【拟解决的关键问题】本研究通过采集川渝两地塔罗科血橙新系病样,进行病原分离鉴定,选取广泛用于炭疽菌遗传分类的GDPH和ITS基因分析炭疽菌的遗传多样性,以期为川渝两地塔罗科血橙新系炭疽病的绿色高效防控和抗病品种选育提供理论基础。
1 材料与方法
1.1 试验材料
2019—2021年,在重庆万州、长寿、开州、奉节、云阳、涪陵,四川资中、富顺等地采集50份疑似柑橘炭疽病症状的塔罗科血橙新系叶片、枝条、果实样品,并记录发病症状。
1.2 试验方法
1.2.1 菌株的分离纯化 参考方中达[34]编著的《植病研究法》,单孢纯化后,接种到PDA培养基斜面,25 ℃培养长出菌落后,4 ℃保存备用。
1.2.2 形态鉴定 将分离纯化的菌株于25 ℃下培养3~7 d,记录菌落形态、颜色,统计平均生长速率。OLYMPUS BX40光学显微镜观察孢子大小,计算平均值。
1.2.3 直接PCR分子检测 试验所用引物见表1[2,15]。先采用通用引物ITS1/ITS4进行PCR测定,初步判定菌种是否为炭疽菌,然后采用种特异性引物Cglnt/Calnt测定炭疽菌种的种类。PCR反应体系:2×Taq PCR Master Mix 15 μL,上下游引物各0.5 μL,ddH2O 14 μL,菌丝少量。扩增反应条件:预变性95 ℃ 3 min;95 ℃ 10 s,59 ℃ 30 s,72 ℃ 20 s,44个循环;72 ℃ 5 min,4 ℃保存。引物GDF1/GDR1采用降落PCR程序,反应条件为:预变性95 ℃ 3 min;95 ℃ 10 s,退火60 ℃ 30 s,延伸72 ℃ 45 s,10个循环,每个循环退火温度下降1 ℃;变性95 ℃ 10 s,退火50 ℃ 30 s,延伸72 ℃ 45 s,35个循环;延伸72 ℃ 5 min,4 ℃保存。
1.2.4 致病性检测 用75%酒精对塔罗科血橙新系的健康果实和保湿培养盒表面进行杀菌处理。在果实最大横径部位选取3处用针刺伤(针刺5点,均匀分布在直径0.5 cm的圆内),取直径0.5 cm的菌块,将菌丝面贴在果实伤口处,放入保湿培养盒,保持湿度95%以上,25 ℃培养7 d,以空白PDA作对照,记录发病状况。每种类型菌株随机选取2个菌株用于致病性检测,每个菌株接种3个果实,重复3次。
1.2.5 序列分析 将ITS、GDPH的PCR扩增产物切胶回收后连接于pMD 18-T载体上,转化大肠杆菌,每个菌株选择3个阳性转化子送上海生工生物测序、BLAST比对。分别将来自柑橘(登录号HM575235.1、HM575241.1、HM575236.1、HM575249.1、AJ301988、AM947679)、柿子(登录号AY791893、AY791894)、草莓(登录号HM575266.1、HM5752 65.1)和草本植物(登录号AY266373、AY266379、AY266382、AY266383)胶孢炭疽菌ITS基因序列与本研究测得的41个ITS基因序列(登录号OL587950~OL587990),利用MEGA 4.0以邻接法(Neighbor-Joining),bootstrap设置为1000,构建系统进化树。将来自柑橘(登录号HM575284、HM575290.1、HM575285)、柿子(登录号GU133383、GU133392、GU133397)和草莓(登录号M93427、HM575315、HM575314.1)胶孢炭疽菌GDPH序列与本研究测得的40个GDPH序列(登录号OP558430~OP558469),利用MEGA 4.0以邻接法(Neighbor-Joining),bootstrap设置为1000,构建系统进化树。

表1 引物信息
2 结果与分析
2.1 病原菌分离及菌株形态
从50份样品中分离到41个菌株,其中10个分离自重庆万州的样品,5个分离自重庆长寿的样品,4个分离自重庆云阳的样品,5个分离自重庆开州的样品,4个分离自重庆涪陵的样品,5个分离自重庆奉节的样品,4个分离自四川富顺的样品,4个分离自四川资中的样品。根据菌落形态、颜色、孢子形态等特征,可以将这些菌株划分为4种类型(图1)。
Ⅰ型菌株菌丝颜色为浅粉色到白色,气生菌丝较少,易产生大量拟菌核和橘红色分生孢子堆,生长速率0.7~0.8 cm/d,分生孢子一端尖,另一端钝圆,大小为(14.7~16.7)μm×(4.9~5.6)μm,此类菌株数量占分离菌株总数的73.17%,是主要类型。
Ⅱ型菌株菌丝颜色为灰色至灰白色,菌丝茂密,菌丝表面不产拟菌核,培养基底部可观察到拟菌核,生长速率1.2~1.3 cm/d,易产生大量橘红色分生孢子堆,分生孢子呈棍棒状,两端钝圆,大小为(14.8~15.2)μm×(4.3~5.5)μm,此类菌株数量占分离菌株总数的17.07%。
Ⅲ型菌株菌丝颜色为深灰色,菌丝茂密,生长速率0.5~0.6 cm/d,色素分泌多,能产生大量的拟菌核和分生孢子堆,分生孢子呈长棍棒状,两端钝圆,大小为(14.8~16.5)μm×(4.3~5.2)μm,此类菌株数量占分离菌株总数的4.88%。
Ⅳ型菌株菌丝颜色为青灰至灰黑色,生长速率1.4~1.5 cm/d,没有产生或少量产生拟菌核,没有观察到橘红色分生孢子堆,分生孢子呈椭圆或梭形,两端钝圆,大小为(13.8~14.1)μm×(4.3~5.0)μm,此类菌株数量占分离菌株总数的4.88%。
2.2 分子鉴定结果
测序比对结果显示,ITS序列均与已报道的柑橘炭疽菌同源性达到99%以上,表明其为柑橘炭疽病菌。Cglnt/ITS4均能扩增出500 bp左右的条带,Calnt/ITS4则均无法扩增到条带,将测序结果进行Blast比对发现,获得序列与柑橘胶孢炭疽菌的ITS序列(AJ301988、AM947679)相似性达到99%以上,说明分离到的柑橘炭疽菌均为胶孢炭疽菌,未分离到炭疽菌其他种。
2.3 致病性测定结果
选择上述4种类型共8个菌株分别接种到塔罗科血橙新系果实上,接种后7 d测量果实病斑直径,比较不同类型菌株的致病力差异。结果如表2、图2所示:4种类型的菌株在塔罗科血橙新系果实上的致病力存在差异。其中,Ⅰ型和Ⅳ型菌株接种后的果实发病率均为88.9%,Ⅰ型菌株接种后果实症状表现为病斑凹陷腐烂,病斑平均直径为1.2 cm,而Ⅳ型菌株仅能形成平均直径为0.6 cm的病斑;Ⅱ型和Ⅲ型菌株发病率均达到100%,其中Ⅱ型菌株刺伤接种后能形成典型的炭疽病斑,病斑呈深褐色,平均病斑直径为1.7 cm,Ⅲ型菌株形成的病斑平均直径为1.1 cm。对照表现不致病,仅观察到1个愈合斑点。方差分析结果显示,4个类型菌株接种塔罗科血橙新系果实后形成的病斑平均直径差异达到显著水平(P<0.05),且Ⅱ型2个菌株(WZ-2和FS-41)与其他3个类型共6个菌株的差异均达极显著水平(P<0.01),Ⅳ型2个菌株(WZ-15和FS-45)与其他3个类型6个菌株的差异也达到极显著水平(P<0.01)。致病性测定结果还显示,不同类型菌株的致病性差异与菌株地理来源、分离部位无明显相关性,不同地理来源和分离部位的菌株可能划到同一类菌株中,相同地理来源和分离部位的菌株也可能划为不同菌株类型。

A.菌丝正面;B.菌丝背面;C.分生孢子。
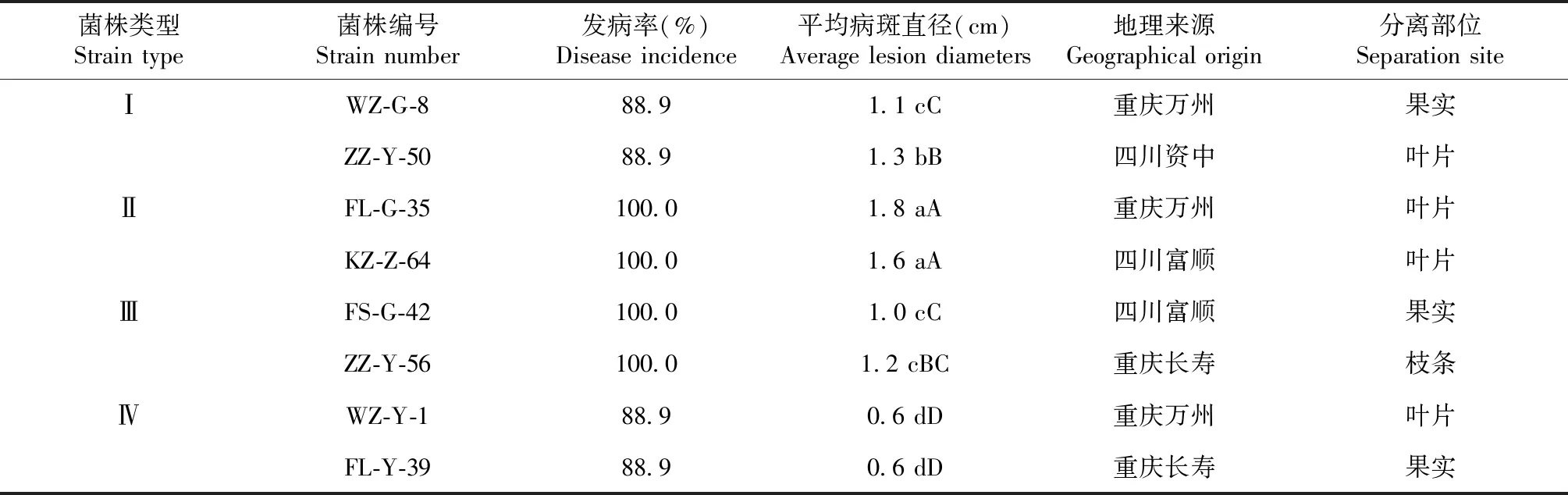
表2 不同菌株对塔罗科血橙新系果实的致病力测定
根据叶片发病症状分为2种类型:第一类症状表现为叶斑型,病斑一般沿叶缘或叶片中部发病,病斑椭圆形或不规则形,病斑直径范围在0.1~3.0 cm,以直径0.5~2.0 cm的病斑居多,病健分界明显,后期病斑中心呈白色纱布透明状,病部散生或轮纹状排列黑色小点,多雨高湿条件下产生肉红色粘质物分生孢子(图3-A,3-B);第二类症状病斑为叶枯型,多在叶片尖端,呈“V”字型,病斑较大,直径在1.0~4.0 cm,病斑后期易干枯或穿孔,多雨高湿时其上产生大量的肉红色粘质物和黑色小点(图3-C,图3-E~3F)。
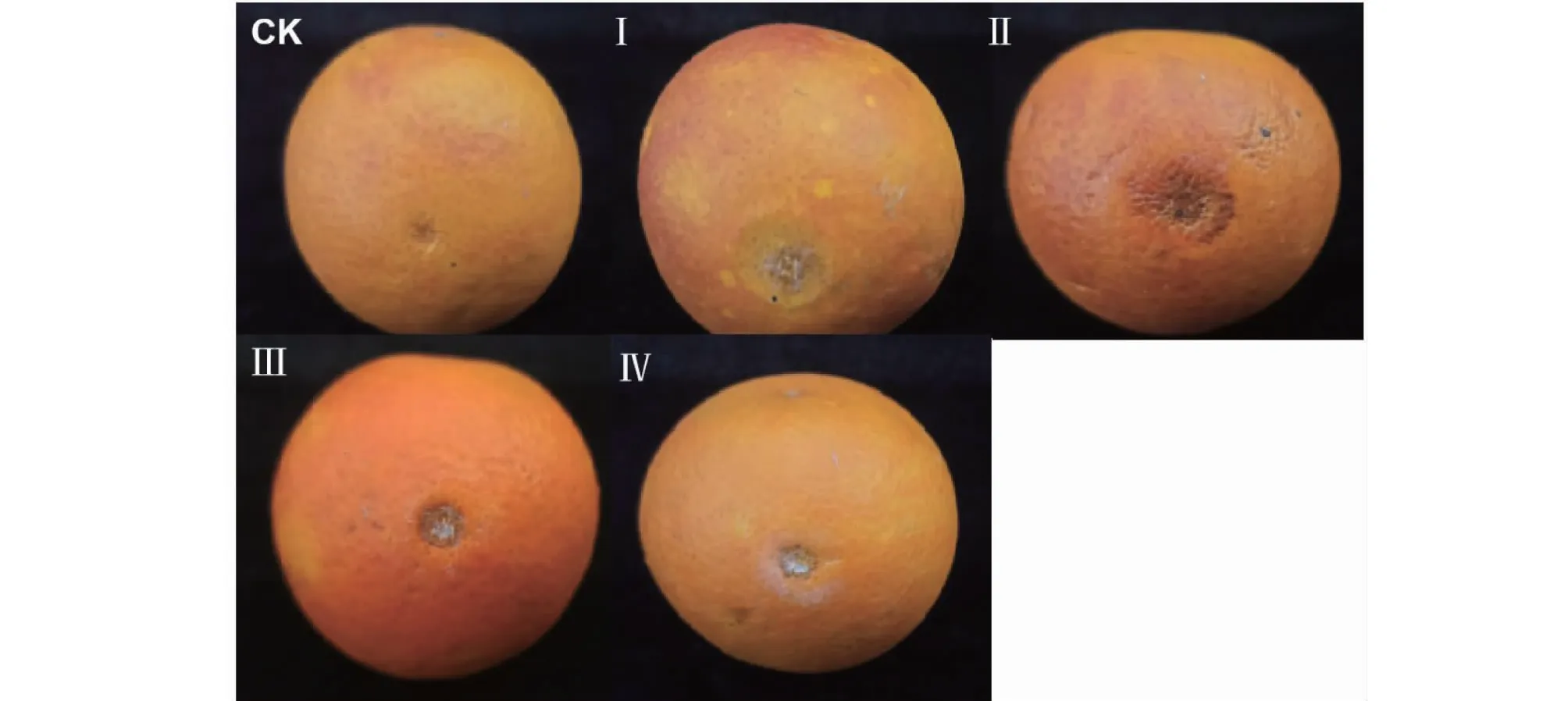
图2 不同类型菌株接种塔罗科血橙新系果实表现

A~B.叶片炭疽病第一类症状;C.叶片炭疽病第二类症状;D.果实炭疽病第一类症状;E.果实炭疽病第二类症状;F.果实炭疽病第三类症状。
果实发病症状也分为3种类型:第一类病斑为干疤型,病斑直径范围在0.2~2.0 cm,圆形或近圆形,黄褐色或褐色,微下陷,呈革质状,发病组织不深入果肉(图3-D);第二类为泪痕型,病斑形状如眼泪,由许多红褐色小凸点组成(图3-E);第三类为软腐型,多在贮藏期发生,一般从果蒂部开始,初期为淡褐色,以后变为深褐色而腐烂(图3-F)。上述叶片和果实症状遇湿度大时,病斑上均可形成橘红色的分生孢子堆。
2.4 菌株的遗传多样性分析
ITS序列比对将供试菌株分为1个类群(图4)。41个菌株与来自柑橘、柿子、草莓和草本植物的胶孢炭疽菌均处于系统发育树的同一个大分支,说明柑橘胶孢炭疽菌与这三类寄主植物的胶孢炭疽菌遗传距离较近,但根据ITS序列无法进行胶孢炭疽菌种下遗传变异的有效区分。根据聚类结果,发现柑橘胶孢炭疽菌的遗传进化与地理来源、分离部位没有明显相关性,不同地理来源和分离部位的菌株都被聚为同一类群中。对比图1和图2,发现柑橘胶孢炭疽菌的遗传进化与菌株形态及致病力也没有明显相关性。
GDPH序列分析结果显示(图5),其菌株聚类与基于ITS序列的分析结果不同,共分为4个类群。与基于ITS序列聚类结果相似,GDPH序列分析结果将来自各采样点的29个菌株与陈国庆[3]报道的柑橘炭疽菌株HB-F01 (HM575284)和HY-UF03(HM575290.1)处于同一分支上,聚为GroupⅠ,为主要菌群;来自重庆涪陵、开州、长寿和四川资中的7个炭疽菌株与陈国庆[3]报道的柑橘炭疽菌株HB-F02(HM575285)聚在一起,为GroupⅡ;来自四川富顺和资中的2个菌株(FS-G-42、ZZ-Y-56)单独聚为GroupⅢ;而来自重庆万州和涪陵的2个菌株与来自柿子的炭疽菌株(GU133383、GU133392、GU133397)聚为GroupⅣ。而来自柿子的胶孢炭疽菌与试验菌株分离度较高,单独聚为一类,说明柿子胶孢炭疽菌与本研究菌株的遗传距离较远。以上结果进一步说明柑橘胶孢炭疽菌遗传变异与地理来源、分离部位没有相关性。与图1、图2测定结果对比,基于GDPH基因序列的遗传聚类结果与基于形态学的分类结果(图1、表2)一致,I型菌株聚到GroupⅠ,Ⅱ型菌株聚到GroupⅡ,Ⅲ型菌株聚类到GroupⅢ,Ⅳ型菌株聚类到GroupⅣ,进化树中的4个Group与形态学和致病性测定结果划分的4个类型菌株组成大体一致,说明不同Group菌株可能与其形态学和致病力相关。

图4 根据ITS序列构建的系统发育树
3 讨 论
柑橘炭疽病是一种为害柑橘生产的重要病害,在我国分布极其广泛。胶孢炭疽菌、平头炭疽菌和尖孢炭疽菌是引起柑橘炭疽病的主要原因,其中胶孢炭疽菌是最主要的类型。本研究从川渝两地50份塔罗科血橙新系疑似炭疽病样品上分离到41个柑橘炭疽菌菌株,通过形态学和ITS序列分析,明确其都为柑橘胶孢炭疽菌。结果表明,胶孢炭疽菌是引起川渝两地塔罗科血橙炭疽病的主要病原菌。
本研究将分离的柑橘胶孢炭疽菌菌株分为4个类型,不同类型的菌株形态特点和致病力存在差异,可能导致不同菌株的危害程度不同。因此,研究炭疽菌种内分化,可为炭疽病病害诊断、病原鉴定、精准高效防治和抗病品种选育提供理论依据。胶孢炭疽菌具有复合种和种群集合的特征,许多研究者也已经证实扁桃[10]、桃子[10]、草莓[24,30,33]、柑橘[3,12,27]、荔枝[34]等寄主炭疽菌种间、种内遗传多样性丰富。本研究发现不同类型的胶孢炭疽菌株形态特点和致病力存在差异,说明川渝地区柑橘胶孢炭疽菌菌株存在丰富的遗传多样性,这可能是由于柑橘胶孢炭疽菌种内出现分化所致。
本研究采用ITS和GDPH两个序列分析来进行柑橘胶孢炭疽菌种内分化研究,结合菌株的地理来源、分离部位、形态学特征及致病力等方面的分析,结果表明柑橘胶孢炭疽菌的种内分化与地理来源、分离部位没有明显的相关性,这与刘开启等[24]、蒲占湑等[25]的研究结果一致,这可能是因为柑橘胶孢炭疽菌有潜伏侵染特性,使同一种菌伴随种苗、接穗等在不同地区进行传播。因此,严格种苗、接穗带菌量检测,从源头上控制炭疽病的传播很关键。
本研究结果还表明基于ITS序列的遗传聚类结果对揭示柑橘胶孢炭疽菌种内遗传变异信息有局限性,这与Moriwaki等[17]的研究结果一致。值得注意的是,与ITS不同,GDPH序列分析结果证明柑橘胶孢炭疽菌的种内分化与菌株形态特点、致病力有紧密关系,能获得种内更丰富的遗传多样性信息,说明GDPH可能与菌株的遗传变异相关,但具体与哪一个或几个形态构成因素有关,还需进一步深入研究。
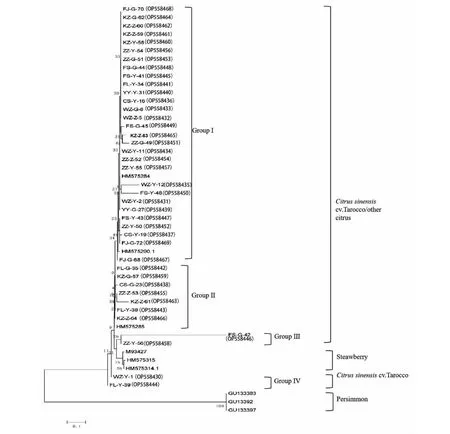
图5 根据GDPH序列构建的系统发育树
4 结 论
本研究明确了川渝两地塔罗科血橙炭疽病的病原菌为胶孢炭疽菌,并且菌株之间存在丰富的遗传多样性,且不同类型的菌株形态特点和致病力存在明显差异,为川渝两地塔罗科血橙新系炭疽病的绿色高效防控和抗病品种选育提供一定的研究基础。
猜你喜欢
今日农业(2022年15期)2022-09-20
现代畜牧科技(2021年4期)2021-07-21
基层中医药(2021年10期)2021-06-05
小猕猴智力画刊(2017年12期)2017-12-27
小猕猴智力画刊(2017年4期)2017-05-04
小猕猴智力画刊(2017年2期)2017-02-18
小猕猴智力画刊(2017年2期)2017-02-18
湖南农业(2016年3期)2016-06-05
浙江柑橘(2016年4期)2016-03-11
营销界(2015年23期)2015-02-28
