
光谱学技术应用于肉类腐败检测的研究进展
2023-09-12 12:44刘庆森董鹏程郝剑刚毛衍伟成海建张一敏
食品科学 2023年15期
刘庆森,罗 欣,董鹏程,郝剑刚,毛衍伟,*,成海建,张一敏,*
(1.山东农业大学食品科学与工程学院,山东 泰安 271018;2.国家肉牛牦牛产业技术体系乌拉盖试验站,内蒙古 乌拉盖 026321;3.山东省农业科学院畜牧兽医研究所,山东 济南 250000)
我国2021年全年猪牛羊禽肉产量8 990万 t[1]。如此巨大产量的肉类不可避免地会受到微生物、添加剂、兽药、非法添加等污染,其中超过25%的肉类不合格率是由微生物引起的[2-3]。在肉的贮藏过程中,由微生物生长及其代谢活动而引起的腐败是造成肉浪费的最主要原因之一,给肉类产业造成巨大的经济损失。肉在腐败过程会产生异味、肉色变暗、表面发黏的现象,使消费者无法接受。一旦微生物总数或某种致病菌超过一定数量,食用后还会给人体带来健康隐患[4]。
传统的微生物检测过程是先通过微生物富集,再进行生化分析确认,最常用的方法为平板培养计数法。虽然该方法具有目的性强、成熟可靠等优点,但其周期长、劳动强度大、会对被测物造成破坏等缺点不容忽视[5]。现代食品质量与安全检测方法要求更加方便、快捷,这也迫切需要引入新型的检测技术,而光谱学技术在快速无损、实时在线等需求方面表现出巨大优势[6]。已有学者对拉曼光谱预测肉色、pH值、嫩度、剪切力等肉类品质指标进行综述[7],虽然涉及肉类腐败预测,但对相关文献总结不够全面。此外,Khaled等[8]对肉类品质和安全的快速检测技术进行了综述,但主要针对食源性致病菌的检测。目前,缺乏关于光谱学技术检测肉类腐败微生物的综述。
本文在分析肉源腐败微生物致腐机理及其预测模型建立过程的基础上,对拉曼光谱、红外光谱和光谱成像技术的原理及研究和应用情况进行阐述,以期为光谱学技术在肉类腐败微生物检测中的应用和进一步研究提供思路和理论指导。
1 肉中的腐败微生物及代谢活动
1.1 初始腐败微生物及优势腐败菌
肉中的初始微生物水平对其货架期具有至关重要的影响,较为常见的腐败微生物有莫拉氏菌、假单胞菌、嗜冷杆菌、肉毒杆菌、沙雷氏菌、索丝菌、埃希氏菌、不动杆菌等[9-10]。在肉类贮藏及流通过程中,不同包装方式(如托盘包装、真空包装以及气调包装(modified atmosphere packaging,MAP))所提供的环境会筛选出特定种类的优势腐败菌[11-12]。
肉在有氧环境中贮藏时,优势腐败菌通常为假单胞菌、热死环丝菌[11-12]。而在无氧条件下,肉中的优势腐败菌主要为乳酸菌、腐败希瓦氏菌、肠杆菌[11-12]。通常情况下,优势腐败菌数量大,提供的信息足以满足快速预测技术实现测定的需求。但要实现对肉类微生物菌群结构的预测,对非优势腐败菌的快速检测变得非常重要。
1.2 腐败微生物的代谢活动
在肉的贮藏和流通过程中,微生物会利用碳水化合物(主要利用糖原分解产生的葡萄糖)、蛋白质、脂肪等营养物质产生氨、硫化物、酮和醛等一系列代谢物,最终导致肉的腐败[13-14]。大多数腐败微生物首先利用肉中残存的葡萄糖产生乙酸、乙偶姻、乙丁酸和3-甲基丁醇等异味物质[15-16]。当微生物繁殖到一定数量,葡萄糖被利用殆尽。腐败微生物开始利用胞外酶将蛋白质降解为氨基酸,氨基酸进一步被分解为胺、氨、硫化物等,而脂肪及脂肪酸会被微生物来源的脂肪酶分解为甘油、醛、酮等化合物,产生难闻的腐败气味[13-14]。
不同的腐败微生物会利用不同的营养物质,并产生特定的代谢产物。假单胞菌是低温有氧贮藏肉中的优势腐败菌,它们优先利用葡萄糖,而后利用氨基酸并产生硫化物、二甲基硫等物质,使肉发生腐败[17]。但假单胞菌在无氧条件下可利用丙酮酸、葡萄糖酸盐、氨基酸进行生长繁殖,产生乙酯、二甲基硫化物[18]。肠杆菌、乳酸菌、热死环丝菌同样是导致肉类腐败的常见微生物,在利用完葡萄糖和葡萄糖酸盐-6-磷酸之后,选择性利用乳酸、葡萄糖酸、丙酮酸、氨基酸、甘油等[19]。乳酸菌、热死环丝菌多产生乙偶姻、乳酸等最终产物,肠杆菌则多与硫化氨类化合物的产生有关[20-21]。
微生物生长代谢导致肉类发生腐败的过程中,以肉为主体的“生态体系”会同时发生许多复杂变化,主要是肉中蛋白质二级结构(α-螺旋、β-折叠)的变化、氨基酸的合成和分解、微生物数量的增加(图1),此类变化是判断肉类腐败的重要信息。葡萄糖是腐败微生物生长繁殖的重要营养物质,可作为微生物快速检测的内标物质,而氨水平的上升同样可成为判定肉类腐败的重要信息。不同的优势腐败菌在生长代谢过程中会产生复杂的代谢副产物,如何寻找优势腐败菌的特异性代谢产物成为实现肉类腐败快速检测的重点及难点。
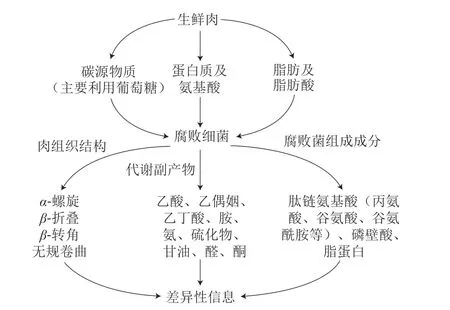
图1 肉类腐败快速检测的物质基础Fig.1 Material basis for the rapid detection of meat spoilage
2 光谱学技术在肉类腐败检测中的应用
2.1 光谱数据处理及模型建立
光谱学技术可以结合化学计量学方法,利用被测物质分子独特的结构及组成成分,将生物学差异信号转变成可视化物理信号数据[22],并进一步建立腐败微生物快速预测模型(图2)。原始光谱既包含目标信息,同时也存在大量的干扰信息。这些噪音主要来源于环境中的杂光、荧光效应、电荷耦合元件(charge coupled device,CCD)检测器噪音、固体颗粒散射等[23]。所以建立预测模型的第一步为降噪处理,方法通常包括基线校正、平滑处理、一阶导数、二阶导数、多元散射校正、标准正太化、去趋势化处理等[24-25]。为进一步筛选目标信息和减少信息的冗杂程度,需要进行特征波段的筛选,例如,竞争性自适应重加权算法(competitive adaptive reweighting algorithm,CARS)[26]可以通过偏最小二乘法(partial least squares,PLS)筛选出与预测目标高度相关的变量并组成新的子集,进而提高模型的运行速度和准确性。主成分分析(principal component analysis,PCA)可以解决变量之间的多重线性相关问题,进而降低数据维度。最后,通过人工神经网络、支持向量机(support vector machines,SVMs)、随机森林、偏最小二乘回归(partial least squares regression,PLSR)等机器学习方法[27-28]建立预测模型。通常,按照3∶1划分训练集和验证集,模型性能的评价指标以决定系数(R2)、相关系数(r)和均方根误差(root mean standard error,RMSE)为主。由于人工神经网络初始权重的随机性、SVMs可配置不同的核函数和惩罚因子等,往往使模型运行结果具有不唯一性,可通过结合遗传算法(genetic algorithm,GA)、连续投影算法和蚁群算法找到机器学习方法的最优解以解决上述问题。随着研究的不断深入,研究人员开始将单个或多个传感器提供的特征波段与实验数据进行分析,如以红外光谱和拉曼光谱所提供的信息互为补充。
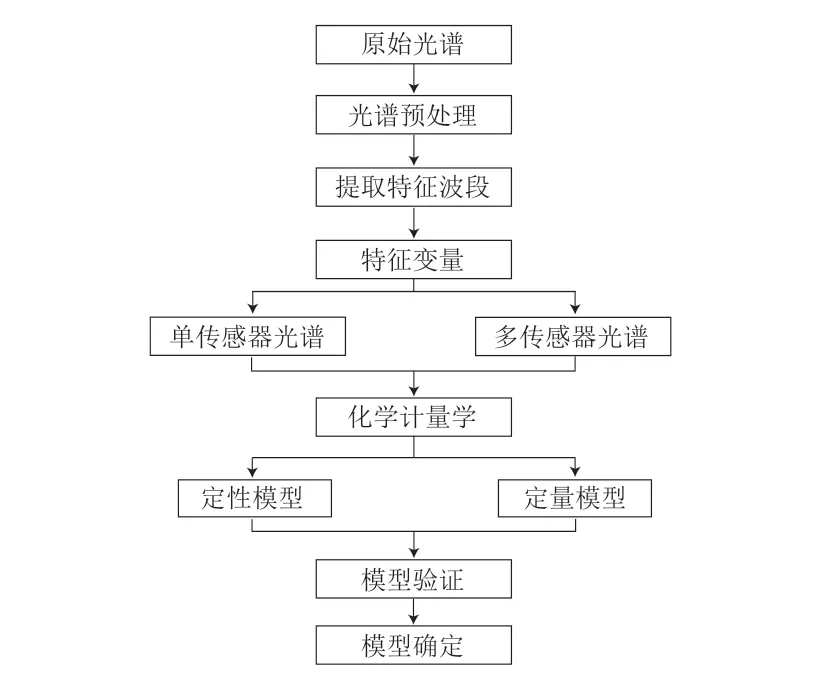
图2 利用光谱技术建立预测模型一般步骤Fig.2 General steps for establishing a prediction model using spectroscopic techniques
2.2 拉曼光谱基本原理及其在肉类腐败检测中的应用
2.2.1 拉曼光谱原理及特征峰
拉曼光谱是分子散射光谱,当待测物中的分子被激发光照射时,分子吸收能量从原始基态跃迁到不稳定的虚态,虚态的分子会释放能量回到基态。如果此时的基态是原始基态,则为瑞利散射;若基态与原始基态存在能级差,则为拉曼散射[29]。拉曼光谱鉴定肉的腐败主要是根据微生物数量和菌群结构的变化、微生物产生的特异性代谢产物以及微生物导致肉本身结构和成分发生的改变。这些变化又导致肉中化学键发生不同的振动,从而使光谱中出现不同的特征峰。例如,酰胺I带(1 645~1 685 cm-1)、酰胺III带(1 200~1 235 cm-1)常常与肉类品质有关。而酰胺I带可反映α-螺旋(1 650~1 658 cm-1)、β-折叠(1 665~1 680 cm-1)、β-转角(1 680 cm-1)和无规卷曲(1 660~1 665 cm-1)信息[30]。
表1总结了部分可用于分析肉类腐败的拉曼特征峰。肉在贮藏过程中,蛋白质的α-螺旋和β-折叠等变化会伴随着C=O、C—N伸缩振动及N—H弯曲振动,拉曼光谱技术可以根据此类化学键的变化来检测肉类腐败。此外,苯丙氨酸(1 002、1 205 cm-1)、色氨酸(755、1 551 cm-1)、酪氨酸(826、855 cm-1)等芳香族氨基酸的拉曼位移峰可作为分析肉类腐败的补充信息[31]。除1 700~500 cm-1所提供的特征峰之外,水和脂类的O—H伸缩振动(3 397~3 309 cm-1)、氨基酸和蛋白质及胺类的N—H伸缩振动(3 303~3 291 cm-1)、过氧酸的O—H伸缩振动(3 284~3 250 cm-1)以及游离氨基酸的NH3+(2 141~1 997 cm-1)也被用于构建腐败微生物的预测模型。
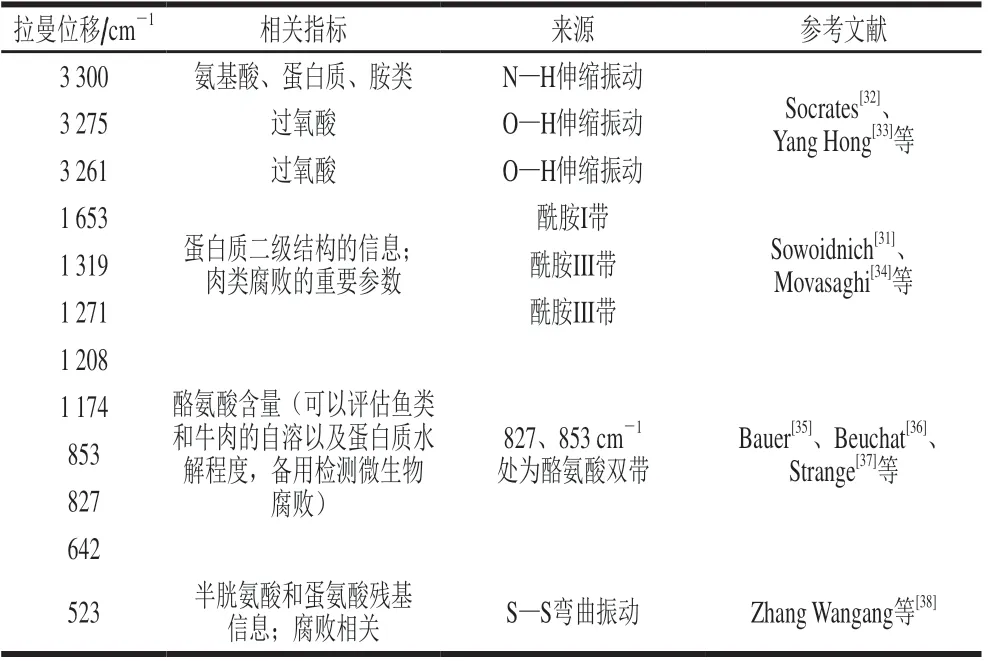
表1 与肉类腐败相关的拉曼光谱特征峰Table 1 Characteristic peaks in Raman spectra associated with meat spoilage
2.2.2 拉曼光谱在肉类腐败检测中的应用
已有学者探究了拉曼光谱技术分析肉类腐败的能力。不同状态鸡胸肉的拉曼光谱可以通过PCA进行区分,进而达到判断新鲜腐败的目的[39]。肉在贮藏过程中会发生变化,进而引起拉曼光谱的改变,寻找与腐败微生物相关的特征峰对预测肉品货架期具有重要意义。Sowoid nich 等[31]通过拉曼光谱结合PCA,并根据1 651 cm-1处的酰胺I带、902 cm-1和935 cm-1处的C—C伸缩振动以及833 cm-1和860 cm-1处的酪氨酸带振动等变化,成功对猪背最长肌和腰大肌的新鲜度进行了判别。仅依靠PCA等方法进行定性分析是远远不够的,开发并建立微生物的定性定量预测模型是关键。
Argyri等[40]尝试利用GA优化人工神经网络预测人工感官评分结果,优化后总体分类精确度达到83.08%。此外,Argyri等[40]利用支持向量回归、PLSR、遗传编程和神经网络对菌落总数(total viable count,TVC)、乳酸菌数量、肠杆菌数量进行定量预测,发现R2集中在0.6~0.8(个别方法R2<0.5),对TVC的预测效果总体好于单菌属[40]。上述结果表明,肉类表面复杂的微生物种类会对单一菌属的预测产生影响。
此前的研究目标多以肉糜为主,而对于大块肉排的研究较少。Yang Hongbo等[41]利用拉曼光谱结合PLSR对生鲜牛排的TVC、假单胞菌、乳酸菌、热死环丝菌进行预测,结果表明,检测细菌分类越精确,拉曼光谱对单菌属数量预测效果相对较差。虽然拉曼光谱结合化学计量学方法能够判别新鲜肉和腐败肉,并能较好地预测TVC,但对于实现精准、实时在线预测肉类剩余货架期及优势腐败菌(种、属)还存在一定的距离。Lu Weilai等[42]利用人工智能结合共聚焦显微拉曼光谱分析了14 种单细胞微生物的不同生长状态,并得到(95.64±5.46)%的分类正确率。在未来的研究中,拉曼光谱可以结合更加合适的人工智能方法分析肉类表面不同微生物的生长状态从而实现更加准确的检测效果。
拉曼光谱在肉类腐败检测中的应用汇总如表2所示。
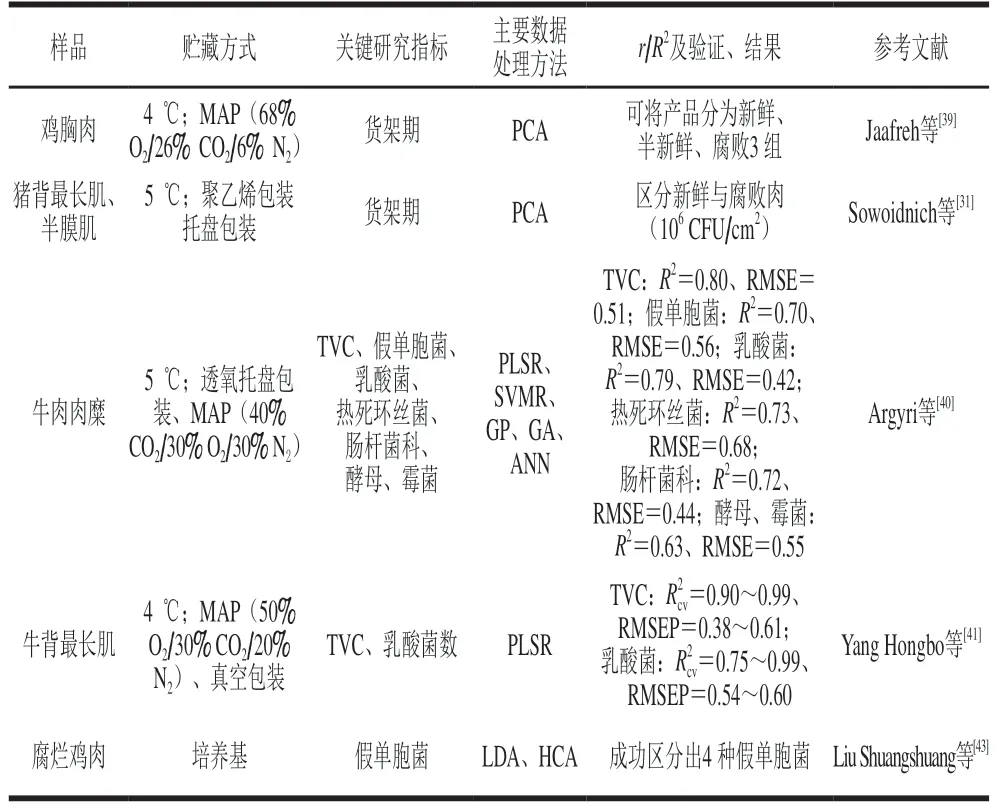
表2 拉曼光谱在肉类腐败检测中的应用Table 2 Application of Raman spectroscopy in meat spoilage detection
2.3 红外光谱在肉类腐败检测中的应用
红外光主要分为近红外光(780~2 500 nm)、中红外光(2 5 0 0 ~2 5 0 0 0 n m)和远红外光(25 000~1 000 000 nm)。近红外光可激发物质分子中的泛音和谐波振动,而中红外光对物质分子的基础振动和相关旋转-振动结构更为敏感,因此多将近红外光谱法和中红外光谱应用于肉类的品质检测[44]。同拉曼光谱法相似,红外光谱法检测肉源细菌主要利用其物质化学键的特征吸收峰(表3)。酰胺I带(1 700~1 600 cm-1)和酰胺II带(1 600~1 500 cm-1)所提供的蛋白质及氨基酸信息是预测微生物的关键,脂肪酸区(3 000~2 800 cm-1)和碳水化合物吸收带(1 500~1 200 cm-1)也可以作为预测模型的主成分。
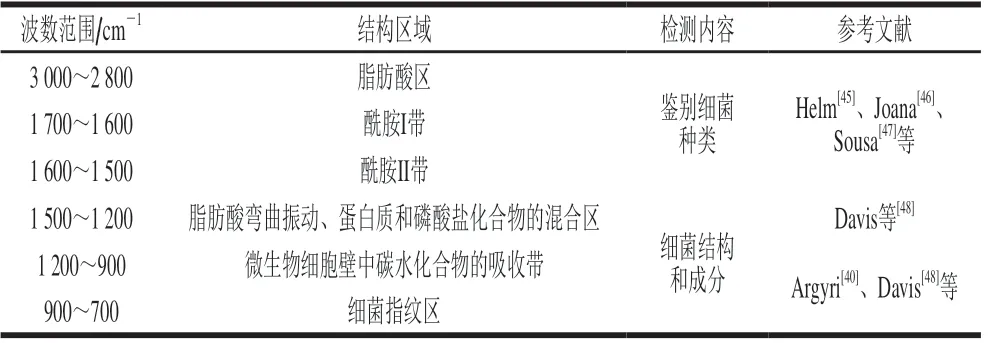
表3 与细菌相关的红外光谱吸收区Table 3 Infrared absorption regions associated with bacteria
2.3.1 以腐败细菌数量为预测指标的研究进展
自2001年起就有学者利用红外光谱法预测肉糜的腐败,主要研究对象为牛肉和禽肉,并初步对TVC进行定量分析[49-50]。随着研究的进一步深入,研究人员开始建立腐败细菌(属、种)的红外光谱定性预测模型。
如表4所示,红外光谱和肉中腐败微生物具有较好的相关性。有学者利用近红外光谱结合PCA成功区分接菌和未接菌的鸡胸肉,但未能成功区分表面接种的英诺克李斯特菌、荧光假单胞菌、恶臭假单胞菌、门多萨假单胞菌和大肠埃希氏菌[51]。与拉曼光谱相同,红外光谱离实现鉴别菌种水平的腐败微生物仍有一定距离,不断提高光谱处理方法是提取重要信息的关键。Kodogiannis等[52]通过反卷积处理红外光谱的洛伦兹宽度,发现游离氨基酸含量与鸡肉腐败程度密切相关。除定性分析外,研究人员也展开了假单胞菌、乳酸菌、肠杆菌和热死环丝菌等优势腐败菌的快速定量分析,并实现了较好的预测效果[53-54]。Argyri等[50]通过不同的数据处理方法对TVC、假单胞菌、乳酸菌、热死环丝菌进行定量分析,R2集中在0.6~0.9(个别方法R2<0.4)。在傅里叶变换红外光谱(Fourier transform infrared spectroscopy,FTIR)结合化学计量学方法对益生菌发酵香肠检测微生物状况的研究中,PLSR模型预测TVC和乳酸菌的RMSE分别为0.641和0.633[55]。目前,尚未有基于FTIR建立益生菌发酵香肠预测接种乳酸菌数量的其他模型,所以结合人工神经网络、SVMs等方法建立预测模型可能会进一步提高该技术的性能。Spyrelli等[56]不但利用PLSR对假单胞菌进行了定量预测,还采用LDA、二次判别分析(quadratic discriminant analysis,QDA)、SVMs进行分析,结果表明QDA结合FTIR光谱预测新鲜腐败状态的效果最好。Spyrelli等[57]进一步通过不同机器学习方法预测鸡胸肉表面的TVC和假单胞菌数量,发现选择合适的数据处理方法对于模型性能具有至关重要的意义。综上,选择合适的实验目标菌相和化学计量学方法可提高预测模型的准确度,这也使红外光谱在肉类腐败判别中表现出更大的潜力。目前,红外光谱结合多元回归法建立肉源腐败微生物的定量预测模型仍处于探索阶段。

表4 红外光谱在肉类腐败检测中的应用Table 4 Application of infrared spectroscopy in meat spoilage detection
2.3.2 以挥发性盐基氮含量作为预测肉类腐败指标的研究进展
总挥发性盐基氮(total volatile basic nitrogen,TVB-N)是我国国家标准中规定的判定肉类腐败的物质,其产生与细菌的代谢活动密切相关。姜新华[59]的研究表明TVB-N含量和TVC的皮尔逊相关系数高达0.902。如表5所示,Wu Xiaohong等[60]在拟合近红外光谱图与TVB-N含量的过程中发现,Adaboost算法可以显著提高模型的判别性能,其结合非相关性判别分析(uncorrelated linear discriminant analysis,ULDA)可使分类正确率达到100%。此外,特征波段的选择尤为重要,波段过长会出现数据冗杂、后期模型运转速度慢等问题,但波段过短会使数据信息缺失,进而导致模型的准确度下降。Ouyang Qin等[61]使用4 种不同的特征波段选择方法,分别建立了预测TVB-N含量的组合区间偏最小二乘法(synergy interval partial least square,Si-PLS)、变量组合总体-分析-偏最小二乘法(variable combination population analysis partial least squares,VCPA-PLS)、CARS-PLS和随机青蛙-偏最小二乘法(random frog partial least squares,RF-PLS)模型,其中RF-PLS预测模型的性能最佳,因为RF选择的特征波段提供了更多与—NH2相关的谱峰。
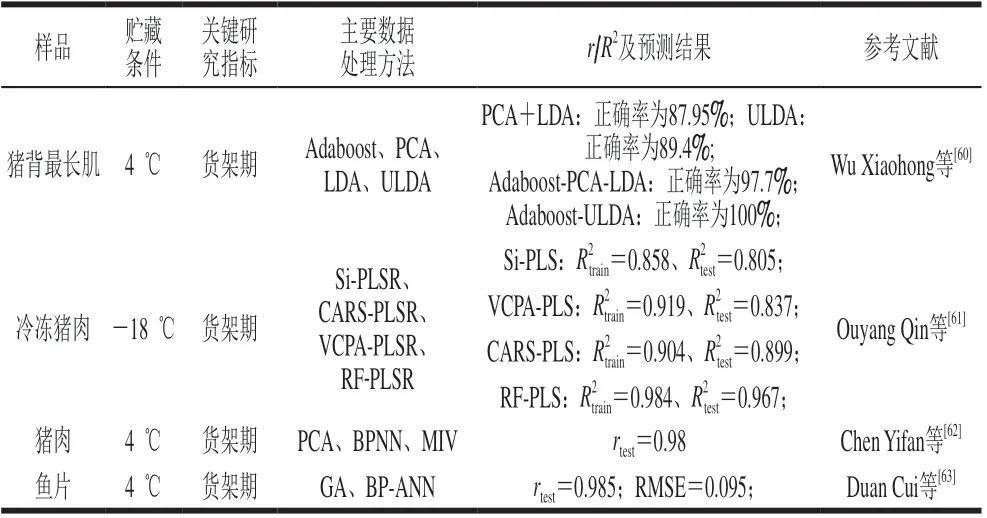
表5 红外光谱预测肉中TVB-N含量的应用Table 5 Application of infrared spectroscopy in predicting TVB-N content in meat
在大多数的研究中,近红外光谱多结合PLSR建立线性回归模型,很少有研究建立非线性预测模型。Chen Yifan等[62]首先利用PCA将数据进行降维处理,然后结合BPNN建立TVB-N含量的预测模型(r=0.96),然后采用MIV预处理方法选择特征波段,提高了模型的预测性能(r=0.98),说明TVB-N含量具有较强的非线性效应。利用便携式近红外光谱仪能够较好地预测鱼片腐败,特别是利用GA优化BP-ANN的方法最为有效,r和RMSE分别为0.985和0.095[63]。
2.4 光谱成像在肉类腐败检测中的应用
2.4.1 光谱成像的分类及特点
光谱成像技术可分为多光谱成像技术、高光谱成像技术和超光谱成像技术[64]。多光谱成像技术同时包括光谱信息和图像信息,相比于拉曼光谱和红外光谱技术,其具有更小的波段和更加明显的特征波段[65]。而高光谱成像技术可获得点、线和面扫描图像[66],其成像方式主要有漫反射、透射和反射成像[67]3 种。超光谱成像技术的优势在于同一时间可测试样品整个区域内所有点的信息,光谱的分辨率也可达到一个更精确的数量级,但对其用于肉类腐败微生物的检测还鲜有研究。在食品微生物检测中,光谱成像主要选用近红外光波段和可见光波段[68]。物质分子结构的特异性决定其在近红外光谱中存在不同的吸收带,进而能够通过差异性图像光谱结合化学计量学方法实现可视化分析。
2.4.2 多光谱成像在肉类腐败菌检测中的应用
基于多光谱成像技术的优势,该技术不仅可应用于肉类色泽、纹理等方面的检测,而且在肉类腐败微生物的检测方面也逐渐发展起来。除对新鲜度和TVC进行预测[69],单菌属的预测也是光谱成像技术应用的方向。Panagou等[70]利用PLSR模型预测TVC、假单胞菌和热死环丝菌水平,结果显示,模型对于TVC的预测效果明显优于假单胞菌和热死环丝菌。与拉曼光谱和FTIR技术相似,预测目标越精确,光谱成像所提供的信息越有限。
数据处理方法对于模型的预测能力至关重要,研究不同的数据处理方法是光谱成像技术发展的前提。Alshejari等[71]研究发现,相较于自适应神经网络模糊推理系统(adaptive neural fuzzy inference system,ANFIS)、多层神经网络(multilayer neural networks,MLP)和PLSR模型,基于自适应模糊推理神经网络(adaptive fuzzy inference neural network,AFINN)的非线性回归模型对TVC具有更好的预测能力。除此之外,研究人员也将不同光谱学技术结合进行微生物预测。在进行相同数据处理的基础上,多光谱成像结合FTIR相比单一多光谱成像能够提供更多的TVC预测信息[72]。不同传感器的信息融合可以提高模型的精确度,为未来光谱学技术应用于肉类的腐败预测提供了新的研究方向。
2.4.3 高光谱成像在肉类腐败菌检测中的应用
相对于多光谱成像,拥有更高精确度的高光谱成像[73]结合化学计量方法在微生物检测方面具有明显优势。研究表明,利用可见光和近红外光范围(400~1 100 nm)的高光谱成像技术能够较好地预测牛肉、猪肉的腐败情况[74]。Barbin等[75]利用PLSR建立TVC和嗜冷菌数(psychrotrophic plate count,PPC)的预测模型,得到预测集的R2分别为0.86和0.89。魏菁[76]通过多种数据处理方法预测冷鲜羊肉的TVC,结果表明,基于极限学习机(extreme learning machine,ELM)预测模型的性能优于PLSR、SVM等,在新鲜度分级模型中,基于混沌粒子群算法(chaos particle swarm optimization,CPSO)的SVM模型的预测效果最好。姜新华[59]采用核稀疏图嵌入典型相关分析法(kernel sparse graph embedding canonical correlation analysis,KSGECCA)对高光谱图像数据进行提取,并通过自适反向神经网络分类法使羊肉新鲜度的总体分类精度达到93.93%。除了预测微生物数量随时间的变化,高光谱提供的特征值还可以用于构建关于微生物的二级生长模型。Zhou Binjing等[77]发现457、509 nm波长处的相对反射率与猪肉表面的假单胞菌数量相关性最强,以其作为特征值构建假单胞菌关于温度的二级生长模型,R2在0.849~0.974范围内。同样,拉曼光谱和FTIR在未来的研究中也可以涉及微生物二级生长模型预测。
高光谱成像作为快速检测技术研究的热点,与FTIR等技术进行信息融合建立预测模型是未来的研究方向之一。相比于单一传感器,多传感器融合特征波段的信息可以提高预测模型的精确度[78]。
光谱成像在肉类腐败检测中的应用研究汇总如表6所示。
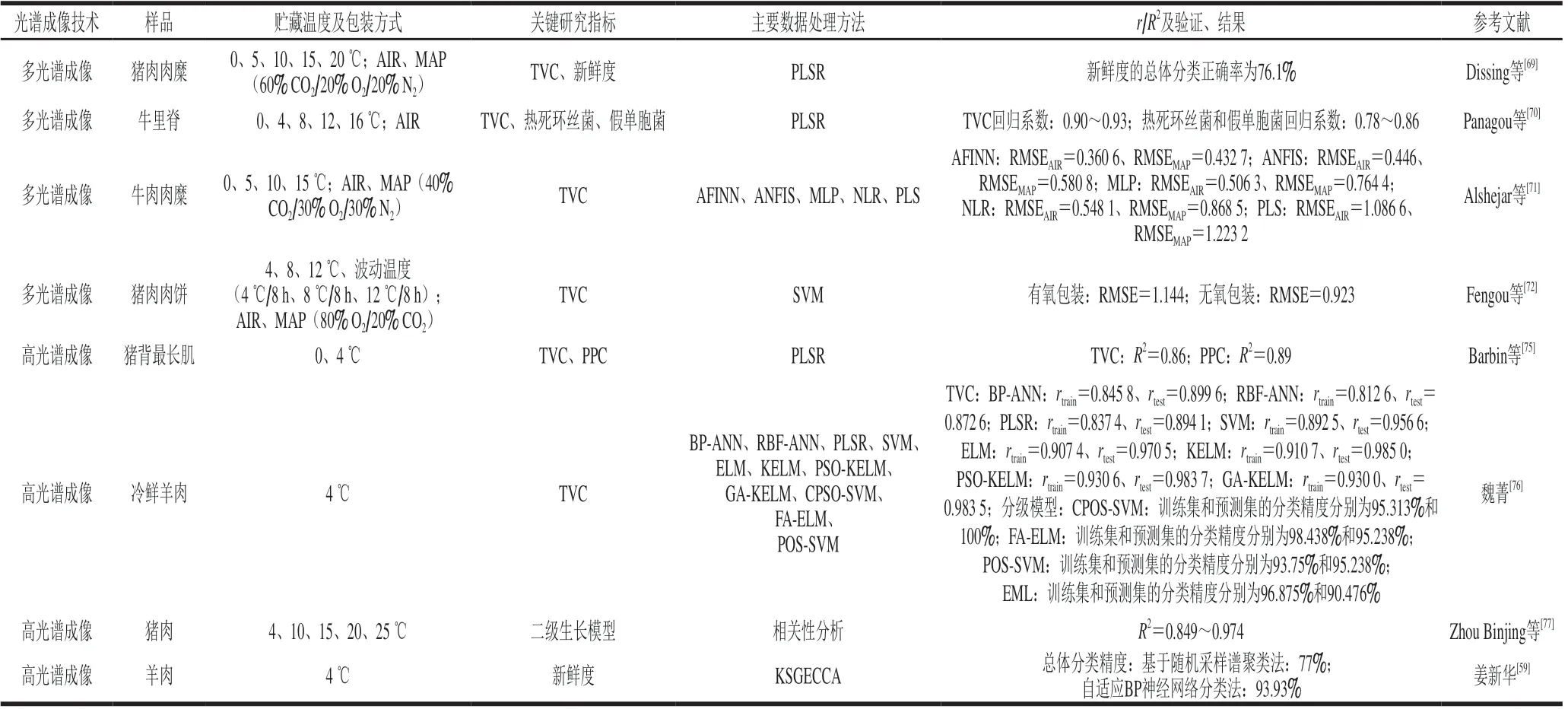
表6 光谱成像在肉类腐败检测中的应用Table 6 Application of spectral imaging in meat spoilage detection
3 结 语
肉在腐败过程中会发生很多变化,如酰胺含量的增加、C=O双键含量的减少、微生物数量的增加并伴随着代谢物质的产生等,可通过拉曼光谱、红外光谱和高光谱成像技术反映此类变化从而实现对肉类腐败的快速检测。3 种技术涉及了肉类新鲜度及其表面优势腐败菌的定性、定量检测,并能够涵盖不同贮藏条件下的猪、牛、羊、禽、鱼等多个物种的分割肉或肉制品。在数据处理上,光谱的预处理方法和机器学习的不断优化,使得腐败预测模型具有良好的分类和预测效果。ELM、KSGECCA法等也被应用于模型的建立中,并能够较好地捕捉光谱中与肉的腐败相关的信息。
虽然拉曼光谱、红外光谱和高光谱成像技术在肉类腐败检测中取到了一定的发展,但依然存在较多的不足。在定量分析中,3 种技术预测肉中TVC的稳定性和精确度仍不能满足肉类工业的实际需求;在定性分析中,3 种技术难以直接对肉中的腐败菌属、种进行区分。不断改进数据处理方法和获取大量的实验样本是光谱学技术解决以上不足的关键,并且可以以不同光谱技术的数据融合作为研究重点。随着计算机技术和编程方法的不断发展,相信未来的光谱学技术会融入更加先进、便捷的数据处理方法,结合光谱仪器在高准确度、低成本等方面的不断升级,并通过系统性地建立不同场景的数据库,最终实现应用于家庭和工业生产实时监测肉品表面微生物数量的目标。另外,通过进一步研究并确定每种腐败微生物的光谱特征峰,光谱学技术手段有望成为快速预测微生物种群结构的实用工具。
猜你喜欢
乳业科学与技术(2021年4期)2021-08-10
乳业科学与技术(2021年3期)2021-08-09
红领巾·探索(2018年11期)2018-12-10
中成药(2017年9期)2017-12-19
中华老年口腔医学杂志(2016年2期)2017-01-15
中国酿造(2015年9期)2015-06-20
现代检验医学杂志(2015年1期)2015-02-06
食品工业科技(2014年23期)2014-03-11
郑州大学学报(理学版)(2013年3期)2013-03-11
化学分析计量(2013年1期)2013-03-11
