
不同放牧强度下祁连山南麓高寒草甸植物群落及土壤水分特征
2023-09-06 10:01陈宇鹏杨永胜王军邦张秀娟
草业科学 2023年8期
陈宇鹏,何 琦,杨永胜,王军邦,张秀娟
(1.长江大学园艺园林学院, 湖北 荆州 434025;2.青海大学生态环境工程学院, 青海 西宁 810016;3.中国科学院西北高原生物研究所 /中国科学院高原生物适应与进化重点实验室, 青海 西宁 810001;4.中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室, 北京 100101)
放牧作为人类在草地生态系统上进行最持久、影响最深远的经济活动之一[1]。其通过采食和践踏等活动,不仅影响土壤容重、孔隙、持水能力及土壤养分等土壤理化特征,而且对植被群落结构、种子萌发、生长、定植及土壤水分循环等生态过程产生深刻的影响[2]。相关研究表明,适度放牧条件下牲畜对土壤的践踏作用使土壤颗粒间隙变小,能够保持和改善土壤含水量[3-4],进而有利于草地利用率的提高和物种多样性的维持。然而,持续高强度的放牧必然会使草地土壤水分急剧丧失[5].,继而引起植被盖度降低、物种减少及植被生产力降低等现象[6],最终导致草地退化,甚至沙化[7]。可以看出,开展不同放牧强度下草地植被群落及土壤属性的变化特征研究,对于草地生态系统的可持续利用和科学管理是非常必要的。
祁连山地区位于青藏高原东北缘,是我国西北地区多条内陆河的发源地[8],在维系区域经济发展、保持水源涵养功能、维持生物多样性及维护生态环境稳定性等方面起着至关重要的作用。高寒草甸是祁连山地区最主要的植被类型之一,由于高海拔、低温、低压及强紫外线等环境特点,造成高寒草甸生态系统极为敏感且脆弱。近几十年来,在诸多人为和自然因素的共同影响下,祁连山高寒草甸发生了严重退化,植物及土壤质量衰退,生产力、经济潜力及服务功能减弱,恢复功能降低甚至丧失[9-10]。就高寒草甸退化的原因而言,多数学者认为自然因素对草甸退化过程有影响作用,人类放牧活动对草甸退化过程起到决定性作用[11]。目前,有关放牧造成高寒草甸生态系统退化的研究多集中在地上部分植被生物量[12]、土壤化学性质[13]和土壤微生物活性[14]等方面,而涉及土壤水分特征变化这一影响草地退化的关键过程和因素[15]方面的研究相对较少,限制了进一步深刻理解过度放牧引起高寒草甸退化的内在过程机制。同时,相关研究多集中在内蒙古温性草原[16]、温带荒漠草原[17]等海拔较低、环境条件相对较好的地区,针对高海拔、自然条件恶劣的高寒草甸地区的研究相对较少。鉴于放牧对草地生态系统的影响具有过程上的复杂性和空间上的分异性,低海拔地区的研究结论并不能直接应用于特殊的高寒草甸地区。因此,十分有必要就放牧强度对祁连山南麓高寒草甸植物群落特征和土壤水分特征的影响进行深入研究。
本研究基于祁连山南麓的高寒草甸放牧管理试验,通过野外实地监测与室内分析相结合的方法,分析不同放牧强度下植物群落组成、生物量、土壤容重和土壤水分特征的变化情况。明确祁连山南麓高寒草甸植物群落结构特征和土壤水分特征对不同放牧强度的响应特征,以期为祁连山南麓高寒草甸生态系统的研究与恢复提供基础数据积累,也为该区域草地资源的合理和持续利用提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于青海海北高寒草甸生态系统国家野外科学观测研究站(海北站),地处青海海北藏族自治州门源县境内祁连山东段南麓,地理位置为37°29′~37°45′ N,101°12′~101°23′ E,海拔3 200~3 600 m。研究区具有明显的大陆性气候特征,无明显的四季之分,仅有冷暖二季之别,干湿季分明,暖季湿润多雨,冷季干燥少雨。年降水量约为580 mm,植物生长季(5 月-9 月)降水量占全年降水的近80%[18]。土壤为暗沃寒冻雏形土,植被类型为典型高寒禾草-嵩草草甸,以矮嵩草(Kobresia humilis)为建群种,以垂穗披碱草(Elymus nutans)、异针茅(Stipa aliena)、高山豆(Tibetia himalaica)、麻花艽(Gentiana straminea)等为主要优势种。
1.2 试验设计
放牧强度试验平台于2011 年8 月在海北站的高寒矮嵩草草甸建立,建立前为牧户冬季放牧草地。参照以往研究放牧强度的经验[19],设置4 种放牧强度,分别为轻度放牧(LG,4.5 羊·hm-2)、中度放牧(MG,7.5 羊·hm-2)、重度放牧(HG,15 羊·hm-2)和极重度放牧(SG,30 羊·hm-2),试验地用围栏围封,LG、MG、HG 及SG 样地面积分别为44 m × 100 m、53 m × 100 m、40 m × 100 m 和40 m × 100 m,每个样地均放牧2 只当地藏系绵羊,每只羊体重在25~30 kg,LG、MG 和HG 样地放牧时间为9 月20 日-翌年5 月31 日,SG 样地为全年放牧。
1.3 研究方法
1.3.1 样品采集
植物群落调查与土壤样品采集于2021 年8 月底进行。以每个样地的中心点为中央设计30 m × 30 m观测区。调查和采样时在观测区以“S”型设置3 个50 cm × 50 cm 观测样方,即每个样地3 个重复样方,样方与围栏距离至少2 m。
1.3.2 植物群落调查
通过观察植物的形态特征,对植物进行分种。在功能群分类中,依据不同科的植物对于家畜的营养价值不同,将禾本科(Gramineae)植物归为禾草类,莎草科(Cyperaceae)植物归为莎草类,其余物种归为杂草类。采用点样框法[20]测定样方内植物群落总盖度及各物种分盖度。使用卷尺测定样方内单种高度和植物群落的平均高度,单种高度至少随机测量8 个重复,若不足8 株则全部测量。测量完成后齐地刈割分种样方内植物地上部分,装入纸质信封中65 ℃烘干至恒重,得到植物群落单种地上生物量及总地上生物量。植物群落调查每个处理设置3 个重复。优势度(重要值)、物种多样性指数、均匀度、优势度指数和重要值等指标采用下列式计算[15]。
式中:RC为相对盖度,Ci为单种盖度;RB为相对生物量,Bi为单种高度;RH为相对高度,Hi为单种盖度;IV为优势度(重要值)。
计算得到群落Shannon-Wiener 多样性指数、Pielou 均匀度指数、Simpson 优势度指数、Patrick 丰富度,相应计算公式为:
Shannon-Wiener 多样性指数(H′):
式中:S为样方内物种数,Pi为第i种植物的相对重要值。
1.3.3 地下生物量及原状土壤样品采集
地下生物量:在群落样品采集样方内,用直径为7 cm 的土钻分层(0-10、10-20、20-30、30-40 cm)采集植物根系和土壤,每个土层采集3 钻样品合为一体,然后用2 mm 筛子将土壤和植物根系分离标记。根系经清洗后装入信封,在65 ℃的烘箱内烘至恒重,得到地下生物量。
原状土壤样品:利用铁锹在采集地上生物量的区域挖掘50 cm 深方形坑,利用内径为5 cm 的环刀,收集0-10、10-20、20-30 和30-40 cm 层面的原状土壤样品,每个样地6 个重复。环刀法采集完土壤样品立即装盖密封,分别标号,装入塑料袋带回实验室,放置冰箱保存备用。
1.3.4 土壤容重及水分性特征的测定
土壤容重及土壤持水能力:将装有湿土的环刀揭去上下底盖,仅留垫有滤纸带孔底盖,放入平底盆内注入并保持盆中水层高度至环刀上沿为止,使其吸水达12 h,此时环刀土壤中所有孔隙都充满了水,盖上上、下底盖,水平取出,用干毛巾擦掉环刀外沾的水,立即称重(M2)。然后,将在吸水12 h 后的环刀仅留垫滤纸的带网眼的底盖,放置在铺有干砂的平底盘中2 h,此时环刀中土壤的非毛管水分已全部流出,但毛细管中仍充满水分,盖上底盖后立即称量质量(M3)。再继续将环刀放置在铺有干沙的平底盘中48 h,盖上底盖称量(M4)。按照下列公式分别计算土壤饱和持水量、毛管持水量及田间持水量:
土壤饱和持水量(W1,mm):
式中:M代表环刀质量(g);H 代表环刀高度,为5 cm;M1代表滤纸重量(g);M2代表吸水12 h 后土壤及环刀质量(g);M3代表放在平盘上2 h 后土壤及环刀质量(g);M4代表放在平盘上48 h 后土壤及环刀质量(g);ρ代表水的密度,为1 g·cm-3;R 为环刀半径,为2.5 cm;10 为cm 与mm 之间的换算系数;h 为土层高度,为10 cm。
将采集的环刀放置在温度为105 ℃的烘箱内,烘干至恒重,称重。按照下列公式计算土壤容重:
式中:D代表土壤容重(g·cm-3);M5代表烘干土壤和环刀总重量(g);V代表环刀体积(cm3)。
土壤饱和导水率:采用定水头法测定土壤饱和导水率。将原状环刀土样有孔盖一端朝下,覆上滤纸,盖上环刀孔盖,浸入水面与环刀上口接近平齐的水中,浸泡8 h 之后将环刀取出,揭去环刀盖,用胶带将其与另外一个空环刀连接,并放置在已架设好铁架台的三角漏斗上,马氏瓶出水口距环刀内土壤表面的高度为3 cm。安装完成后,马氏瓶开始供水,使土壤表层形成3 cm 的稳定水层,持续供水4 h后测定,每隔5 min 记录渗出水量,同时记录水温。当最终测定的渗出水量基本不变或者循环出现时,停止测定。取最终稳定值计算饱和导水率(Ks)。
式中:Q为一段时间内的流量(cm3);L为土柱高度(cm);A为土柱截面积(cm2);t为时间(s);H为水头高度(cm)。
为方便对比,将不同水温下饱和导水率统一为t = 10 ℃的值,计算公式如下:
式中:K10为温度等于10 ℃时的饱和导水率(mm·min-1);Ks为T时的饱和导水率(mm·s-1);T为测量时的水温(℃)。
1.4 数据分析
运用Excel 2019 对数据进行整理,数据表达为平均值 ± 标准误差。处理间的差异用SPSS 25.0 中的One-way ANOVA 模块实现。利用Origin 2018 绘图。
2 结果与分析
2.1 植物群落结构特征
2.1.1 植物种群组成
随着放牧强度的增加,祁连山南麓高寒草甸植物群落组成发生明显变化(表1)。就物种数而言,轻度、中度、重度及极重度放牧样地中统计到的总物种数分别为14 科38 种、12 科29 种、13 科39 种、11 科15 种,总体呈下降趋势。从优势种角度而言,轻度、中度和重度放牧样地中重要值最大的均为山地早熟禾(Poa orinosa),分别为19.19、17.31 和21.75;在极重度放牧样地中,鹅绒委陵菜(Potentilla anserina)重要值最大(43.03)。在轻度放牧样地中,异针茅、羊茅(Festuca ovina)、钝 苞 雪 莲(Saussurea nigrescens)和鳞茎堇菜(Viola bulbosa)在群落中具有较明显优势。在中度放牧样地中,具有较明显优势的物种有羊茅和异针茅,均为禾草类植物;在重度放牧样地中,仅有异针茅具有较明显优势。极重度放牧样地中,莎草类植物矮生嵩草、暗褐薹草(Carex atrofusca)和杂草类植物二裂委陵菜(Potentilla bifurca)、海 乳 草(Glaux maritima)、西 伯 利 亚 蓼(Polygonum sibiricum)具有较明显优势。

表1 祁连山南麓不同放牧强度下各物种的重要值Table 1 Importance value indices of species under different grazing intensities on the southern slope of Qilian Mountains
2.1.2 功能群
随着放牧强度的增加,祁连山南麓高寒草甸植物功能群呈先稳定后显著变化的趋势,自轻度放牧至重度放牧,禾草类、莎草类以及杂草类植物重要值占比分别稳定在50.52%、5.05%和44.43%上下(图1)。至极重度放牧,禾草类植物的重要值占比较重度放牧显著下降83.01% (P< 0.05),莎草类和杂草类植物重要值占比较重度放牧分别显著提高365.59%、70.08% (P< 0.05)。可以看出随着放牧强度的增加,禾草类植物生长逐渐受到抑制,莎草类植物得到生长空间,毒杂草成为优势种,高寒草甸生产功能和饲用功能逐渐减弱。
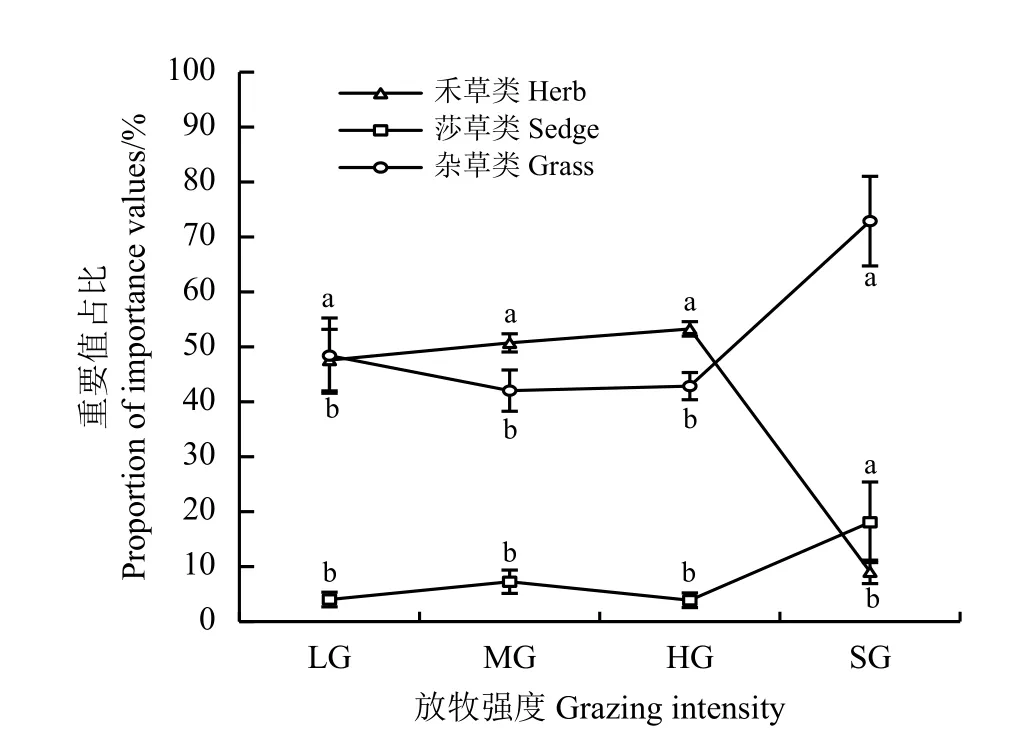
图1 祁连山南麓不同放牧强度下各功能群的重要值占比Figure 1 The proportion of importance values of each functional group under different grazing intensities on the southern slope of Qilian Mountains
2.1.3 高度与盖度
祁连山南麓高寒草甸植物群落总盖度和平均高度呈现先稳定后下降的趋势(图2)。从轻度放牧至重度放牧,植被盖度和高度分别稳定在92.8%和21.1 cm 上下,至极重度放牧时,二者分别显著下降25.0%和94.8% (P< 0.05)。
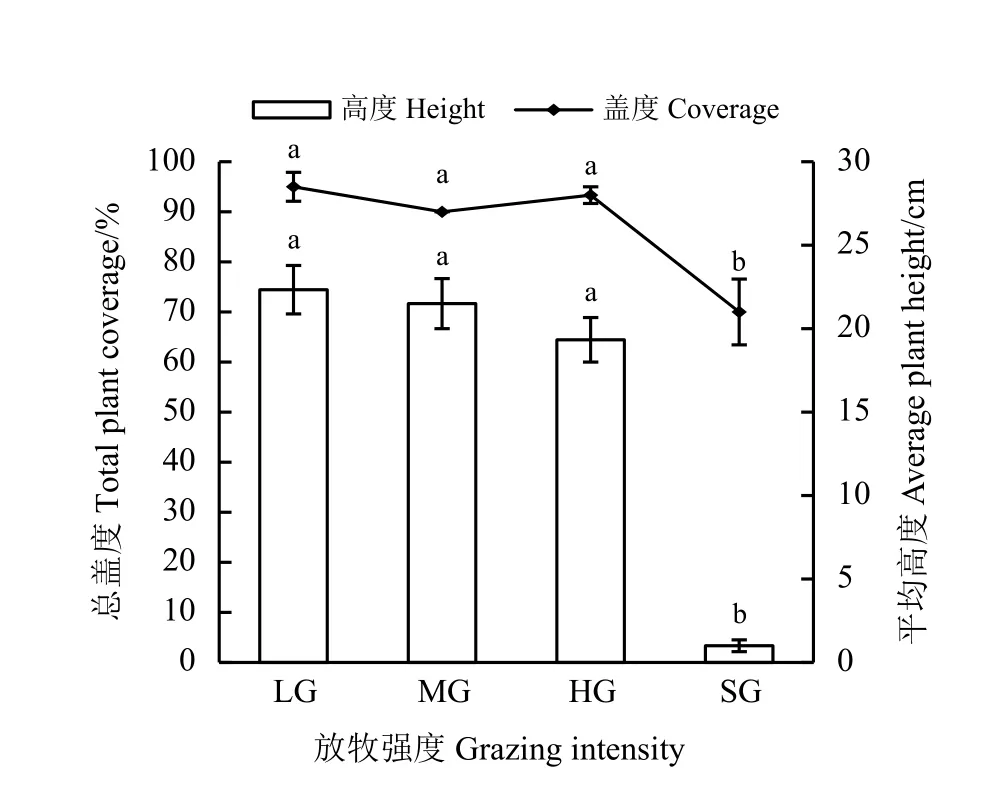
图2 祁连山南麓不同放牧强度下植被平均高度及总盖度Figure 2 Average vegetation height and total coverage under different grazing intensities on the southern slope of Qilian Mountains
2.1.4 生物量
祁连山南麓高寒草甸生物量主要集中在地下部分,不论放牧强度高或低,地下生物量占总生物量的比值均在83.0%以上。随着放牧强度的增加,地下生物量占总生物量的比值呈先稳定后增加的趋势。轻度至重度放牧,地下生物量占总生物量的比值基本稳定在84.3%上下,至极重度放牧,地下生物量占总生物量比值达到88.4% (图3A)。深入分析不同深度地下生物量,可知祁连山南麓高寒草甸地下生物量主要集中在0-10 cm,随着深度的增加,地下生物量逐渐降低(图3B)。同时,随着放牧强度的增加,0-10 cm 地下生物量占总地下生物量的比例呈先稳定后增加的趋势,从轻度至重度放牧,0-10 cm 地下生物量占总地下生物量的比例基本维持在70.8%上下,至极重度放牧,0-10 cm 地下生物量占总地下总生物量比值达到80.8% (图3B)。说明随着放牧强度的增加,高寒草甸植物物质和能量向地下部分,特别是0-10 cm 层面的分配比例在增加。
祁连山南麓高寒草甸地上生物量随放牧强度增加呈先缓慢增加后显著降低的趋势。从轻度放牧至重度放牧地上生物量呈缓慢增加趋势,但未达到显著水平(P> 0.05),重度放牧样地地上生物量较轻度放牧增加15.1% (图3A)。至极重度放牧,其地上生物量较重度放牧显著降低71.9% (P< 0.05)。地下生物量及总生物量随着放牧强度的增加呈先稳定后显著降低的趋势,从轻度放牧至重度放牧,二者分别 稳 定 在2 206.08、2 617.03 g·m-2上 下(图3A)。至极重度放牧,二者较重度放牧分别显著降低55.96%、58.7% (P< 0.05) (图3A)。
随着放牧强度的增加,地下生物量均呈先稳定后降低的变化趋势(图3A),从轻度放牧至重度放牧0-10、10-20、20-30 和30-40 cm 地下生物量分别 稳 定 在1 560.5、344.8、192.7 和108.1 g·m-2上 下(图3B)。至极重度放牧,0-10、10-20、20-30 和30-40 cm 地下生物量分别较重度度放牧显著(P<0.05)降低52.2%、70.4%、71.7%和42.7% (图3B)。
由图4 可知,随着放牧强度的增加,地上与地下生物量比值呈先上升后下降的趋势,且未达到显著差异水平(P> 0.05)。重度放牧相较于轻度放牧增加了11.9%,极重度放牧相较于重度放牧降低了32.2%。
2.1.5 植物种群特征性指数
随着放牧强度的增加,Shannon-Wiener 多样性指数、Simpson 均匀度指数和Patrick 丰富度指数都呈现出先稳定后降低的趋势(表2)。相较于重度放牧,极重度放牧强度下Shannon-Wiener 指数、Simpson指数和Patrick 指数分别显著 降低41.8%、20.9%、68.5%(P< 0.05)。Pielou 优势度指数随放牧强度的增加呈下降趋势,各处理间差异未达到显著水平(P> 0.05)。
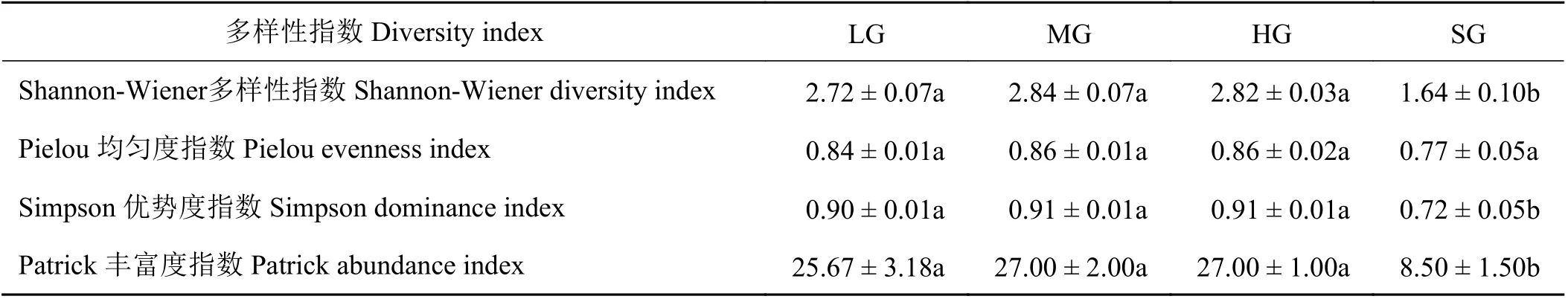
表2 祁连山南麓不同放牧强度样地的多样性指标Table 2 Diversity indices resulting from different grazing intensities in southern foothills of Qilian Mountains
2.2 土壤容重及水分特征
放牧强度对祁连山南麓高寒草甸土壤容重和水分特征的影响随深度的增加逐渐减弱,其作用主要集中在0-10 cm 层面(表3)。随着放牧强度的增加,0-10 cm 土壤容重、饱和导水率和饱和持水量呈现先稳定后显著变化的趋势,从轻度放牧到重度放牧,三者分别稳定在0.92 g·cm-3、0.24 mm·min-1、61.16 mm 上下。至极重度放牧,0-10 cm 土壤容重较重度放牧显著增加12.6% (P< 0.05),0-10 cm 饱和导水率和饱和持水量较重度放牧分别降低50.0%和5.9% (P< 0.05)。放牧强度对0-10 cm 毛管持水量、田间持水量及10-20、20-30 cm 土壤容重、饱和导水率、饱和持水量、毛管持水量、田间持水量均无显著影响(P> 0.05)。
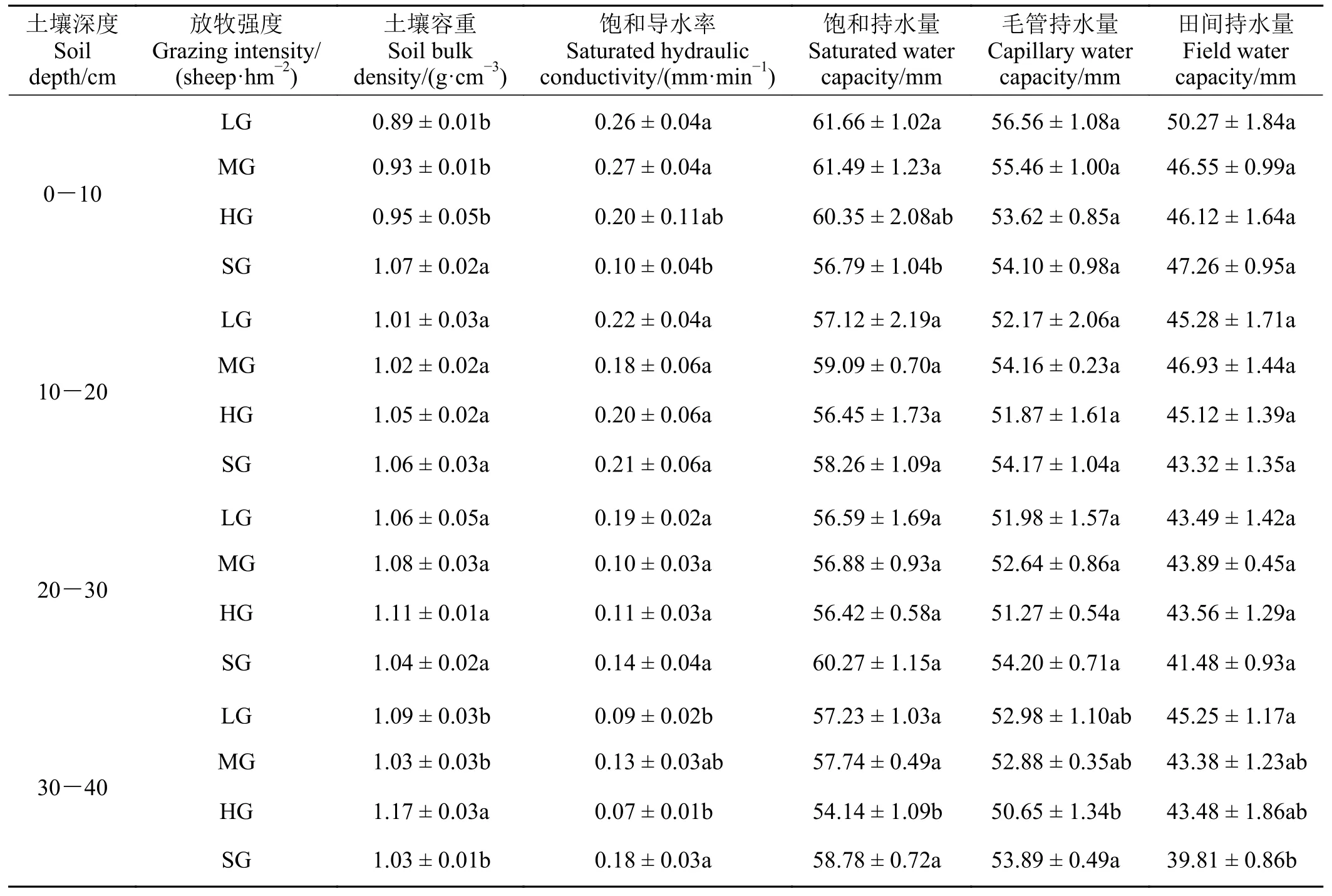
表3 祁连山南麓不同放牧强度样地土壤容重及水分特征Table 3 Soil bulk density and water characteristics of different grazing intensities on the southern slope of Qilian Mountains
值得注意的是,重度放牧样地30-40 cm 土壤容重、饱和导水率、饱和持水量、毛管持水量与其他放牧强度样地均呈现不同程度的显著性(P< 0.05)差异。其中,与重度放牧样地相比,极重度放牧样地土壤容重显著降低12.0% (P< 0.05),饱和导水率、饱和持水量和毛管持水量分别显著增加157.1%、8.6%、6.4% (P< 0.05)。
3 讨论
3.1 放牧强度对植物种群特征的影响
放牧对草地生态系统的影响首先会表现在植物群落特征和组成上。在放牧过程中,家畜通过对植被的踩踏和啃食能直接影响其盖度、高度等植被数量特征[21]。本研究表明,随着放牧强度增加,植物群落总盖度、平均高度、Shannon-Wiener 指数、Simpson 指数和Patrick 指数均先稳定后显著下降,植被群落结构发生明显改变,禾草类植物生长逐渐受到抑制,莎草类植物得到生长空间,毒杂草成为优势种(图1),高寒草甸生产功能和饲用功能逐渐减弱。这与前人的研究结果基本一致[22-23],出现这种现象的原因主要是,在一定的放牧强度阈值范围内(轻度放牧至重度放牧),植物群落及土壤对放牧活动造成的干扰具备一定的自我调节能力,植物可通过补偿生长等方式维持群落稳态。然而,一旦放牧干扰强度超过高寒草甸生态系统能够恢复稳态的阈值(极重度放牧),家畜喜食且高度相对较高的禾草类植物被过度采食,造成其优势度明显下降(图1、图2)。且由于家畜的频繁踩踏,必然会导致植被群落盖度和高度的快速下降。正是由于相对较高的禾草类植物优势度下降,造成禾草类植物对降水的截留作用和对阳光的遮蔽作用均明显降低,使相对低矮的莎草类和毒杂草植物能接收更多的光照和水资源[24],进而促使莎草类和毒杂草植物更好的生长。此外,由于莎草科植物具有株型低矮、分蘖能力强、根系发达且耐牧性较强的特点,家畜不喜食毒杂草等习性[3],促使莎草类和毒杂草植物能够很好的抵御极重度放牧带来的干扰。最终导致在极重度放牧条件下莎草类和毒杂草植物优势度明显提高,Shannon-Wiener指数、Simpson 指数和Patrick 指数显著下降。
3.2 放牧强度对生物量的影响
生物量在一定程度上代表草地的净初级生产力,能直接反映出植被对资源的竞争能力[25]。本研究结果显示,祁连山南麓高寒草甸的生物量主要集中在地下部分,且地下生物量的70%以上均集中在0-10 cm。这与前人研究结果[26-27]一致。这是因为高寒草甸区域植物以根系发达的禾草类和莎草类植物为优势种[28],植物将更多的生物量分配到地下来获取养分和水分资源。随着土层深度的增加,土壤水分、土壤温度、土壤养分和孔隙度降低,土壤紧实度增加,抑制了根系的生长,直接降低了根系生物量[29]。从轻度放牧到重度放牧阶段,地上生物量和地上生物量与地下生物量比值均呈缓慢增加趋势,说明在一定放牧强度条件下,放牧干扰可以提高祁连山南麓高寒草甸地上植被生产力。原因如下:适度的放牧条件下,牲畜的采食可以去除衰老组织,使存留叶光合作用增强、水分养分供给提高、新生叶光合速率增加[30],刺激植物向地上分配更多的物质和能量以增加植物的竞争优势。家畜的活动与排泄可以促进植物种子的传播,同时,家畜排泄物中的养分促进了植物的生长[31]。然而,从重度放牧至极重度放牧阶段,地上生物量、地下生物量及地上地下总生物量均显著低,且地下生物量占总生物量的比例及0-10 cm 地下生物量占总地下生物量的比例均呈增加趋势。说明一旦放牧强度超过草地生态系统所能承受的阈值,随着放牧强度的增加,植物生产力必然降低,且植物的物质和能量分配必然会向地下部分,特别是0-10 cm 层面增加,出现根系生物量向土壤表层积聚现象[32-33]。这主要是因为,高强度的放牧导致土壤容重增大,进而造成土壤渗透性、通气性与持水能力减弱,阻碍养分的溶解与移动,减缓植物根系对水分的吸收,从而降低草地的植物群落生产力,阻碍牧草的生长发育[34]。由于牲畜的反复啃食、践踏,处于生长期的植物发育受到抑制,植物不能弥补强放牧干扰造成的损失导致总生物量下降[35]。极重度放牧由于种间竞争减弱,总生产力下降,植物倾向于将物质与能量向下分配以获得更多的水分与养分以适应强放牧干扰[36]。极重度放牧条件下莎草类植物明显增加,其特殊的生物学特性(高地下/地上比)[37]必然促使生物量逐步向地下转移,由于莎草类植物根系多为须根系,主要分布在浅层土壤,所以0-10 cm 地下生物量占比明显增加。
3.3 放牧强度对土壤水分特征的影响
土壤容重可以反映土壤的松紧程度和土壤结构状况,土壤饱和导水率、饱和持水量、田间持水量及毛管持水量是衡量土壤导水和持水能力的重要参数,可以衡量土壤的通透性和贮水能力[38],都常用于评价土壤的水源涵养功能。本研究结果表明,放牧强度对连山南麓高寒草甸土壤容重及水分特征的影响主要集中0-10 cm 层面。从轻度放牧至重度放牧,土壤容重、饱和持水量、饱和导水率、土壤饱和持水量、田间持水量及毛管持水量基本稳定。这主要是因为:高寒草甸表层土壤中分布着由密集根系组成的草毡层[39-40],其致密且富有弹性,极耐牲畜践踏,能在一定程度上缓冲家畜对土壤的践踏作用,降低了家畜践踏对深层土壤水分特征指标的影响。本研究涉及的草地为冬季放牧,冷季漫长而寒冷,土壤冻结后家畜对于土壤的践踏作用有限,促使土壤水分特征指标对放牧强度的敏感性低于植被群落特征和生产力对放牧强度的敏感性,土壤水分特征指标的变化需要更长时间积累。至极重度放牧时,0-10 cm 土壤容重显著增加,饱和导水率、饱和持水量、田间持水量和毛管持水量均呈下降趋势,这与祝景彬等[41]的研究结果一致。这主要是因为极重度放牧条件下,植被群落盖度、高度、生物量均明显下降,莎草科和杂草类植被优势度明显增加,高寒草甸草毡层根土比和厚度逐渐增加并出现老化死亡,在冻融交替作用下,老化死亡的草毡层在脆弱处被撕裂,最终局部形成草毡层彻底消失的黑土滩[42],其在家畜高强度的踩踏及大量粪尿排泄情况下土壤表面压实板结[43],土壤容重显著增加,导致土壤孔隙度降低,进而促使土壤饱和持水量及导水率明显下降。可以看出,当高寒草甸草毡层存在时,放牧强度变化对土壤水分特征指标的影响较弱,一旦草毡层消失,高强度的放牧干扰必然导致土壤水分特征指标的持续恶化。因此,保持适当厚度的草毡层是维持祁连山南麓高寒草甸水源涵养功能的关键,在祁连山南麓高寒草甸的利用过程中一定要重视草毡层的保护。
本研究还发现,重度放牧样地30-40 cm 土壤容重、饱和导水率、饱和持水量、毛管持水量、田间持水量与其他放牧强度样地均呈现不同程度的显著性差异。深入分析其原因,发现主要是由各处理在30-40 cm 深度砾石比不同而导致的(表4),说明砾石比对土壤持水能力及导水能力有着较大影响[44]。目前,对于土壤砾石比空间格局在高寒草甸土壤水文过程和养分循环的影响仍研究较少,十分有必要进行针对性的深入研究。
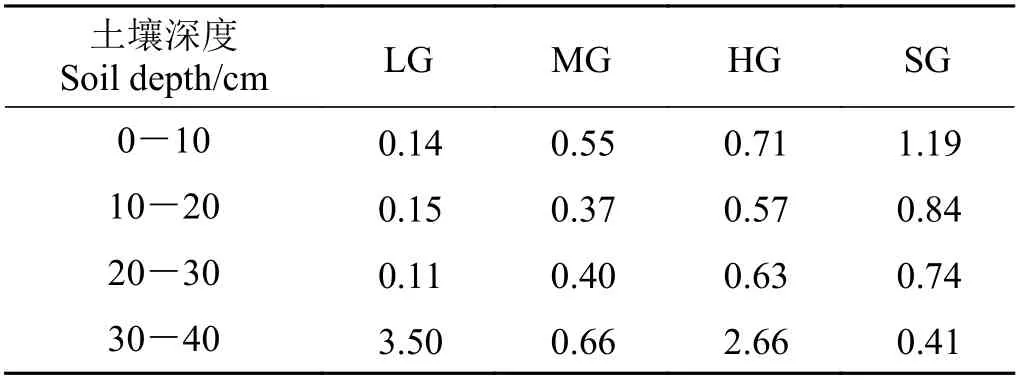
表4 祁连山南麓不同放牧强度样地砾石比Table 4 Gravel ratio for different grazing intensities on the southern slope of Qilian Mountains%
4 结论
放牧干扰明显改变了祁连山南麓高寒草甸植物群落结构。随着放牧强度增加,植物群落总盖度、平均高度、Shannon-Wiener 指数、Simpson 指数和Patrick 指数均先稳定后显著下降,植被群落结构发生明显改变,禾草类植物生长逐渐受到抑制,莎草类植物得到生长空间,毒杂草成为优势种。
从轻度放牧到重度放牧,放牧干扰可以提高祁连山南麓高寒草甸地上植被生产力,地上生物量和地上生物量与地下生物量比值均呈缓慢增加趋势。至极重度放牧阶段,高寒草甸地上生物量、地下生物量及地上地下总生物量均显著降低,地下生物量占总生物量的比例及0-10 cm 地下生物量占总地下生物量的比例均呈增加趋势。
放牧强度对连山南麓高寒草甸土壤容重及水分特征的影响主要集中0-10 cm 层面。从轻度放牧至重度放牧,土壤容重、饱和持水量、饱和导水率、土壤饱和持水量、田间持水量及毛管持水量基本稳定。至极重度放牧时,0-10 cm 土壤容重显著增加,饱和导水率、饱和持水量、田间持水量和毛管持水量则呈下降趋势。高寒草甸草毡层在维持土壤涵养功能方面起着至关重要的作用。
猜你喜欢
疯狂英语·新读写(2023年4期)2023-05-10
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
读者·原创版(2021年2期)2021-02-26
活力(2019年21期)2019-04-01
华人时刊(2016年17期)2016-04-05
黄河之声(2016年20期)2016-02-21
黄河黄土黄种人·水与中国(2014年11期)2014-12-16
河南水利与南水北调(2014年24期)2014-11-26
自然资源遥感(2014年3期)2014-02-27
