
过表达ERF76 小黑杨的木材组分分析及影响
2023-09-05 01:47:16廖诗贤王宇婷姜廷波
山东林业科技 2023年4期
李 鑫,廖诗贤,王宇婷,姜廷波
(东北林业大学林木遗传育种国家重点实验室,黑龙江 哈尔滨 150040)
杨树(populus)属杨柳科(Salicaceae)杨属(Populus)植物,分布广泛、适应性强、在我国约有60 多种[1]。杨树所具有的速生特性使其成为林木产品生产的重要原料之一,主要用于生物质能源制造、民用建筑、板料及纸浆制造等[2]。其中小黑杨(Populus simonii×P.nigra)是通过小叶杨(Populus simonii Carr)和黑杨(Populus nigra)杂交而来的优良品种[3]。为培育品质更为优良的树种,利用基因工程将转录因子转化到植物体内来改变植株部分特性是改良树种的有效途径之一。AP2/ERF 家族是一个庞大的植物特异性转录因子群,在许多生物过程中发挥重要作用[4]。研究所涉及的ERF 类转录因子参与植物一系列的重要生命活动,包括激素信号诱导调控、代谢调控、细胞分化、植物生长发育等[5]。根据报道,ERF76 转基因杨树中植物激素ABA 和GA 生物合成和信号传导通路相关基因的相对表达水平显著高于野生型[2]。其中GA 参与调控纤维素的合成,并通过DELLA 参与调控[6]。这为ERF76 基因影响杨木生长发育进而影响木材化学组分提供了一定的理论基础。而化学组分原料主要为纤维素、半纤维素、木质素等,是有关木材产品生产最主要的原料[7-10]。对于这些组分的分离与应用工艺有许多,例如碱/尿素处理工艺[11]、稀酸处理[12]、水性助溶剂处理[13]、自水解[14,15]、碱处理[16]等处理方式。考虑到能源、机械等实际生产条件以及实验室条件,文章采用氢氧化钠对目标杨木进行碱法处理以及半纤维抽提[17]工艺条件进行试验。
文章研究的主要目的有二,首先是探究过表达ERF76 小黑杨在基因的影响下木材中的化学组分发生了哪些变化。其次是通过碱处理工艺处理探索由该转基因造成的化学组分变化在工业化应用处理过程中会造成哪些影响,意在对日后该转基因木材的合理应用提供一定的理论与数据支持。
1 实验材料与方法
1.1 实验材料
1.1.1 植物材料实验选用位于东经125°42’—130°10′,北纬44°04′—46°40′野生型小黑杨(WT)与ERF76 过表达转基因小黑杨(2 年生与6 年生)。取样时间为2022 年6 月
1.1.2 试剂氢氧化钠、72%浓硫酸、4%硫酸、苯、95%乙醇、70%乙醇、亚氯酸钠、去离子水、冰醋酸、丙酮、饱和氢氧化钡溶液、邻苯二甲酸氢钾
1.2 杨木化学组分测定方法
水分、有机物抽提、综纤维素、灰分分别根据国家标准GB/T462-2008、GB/T2677.6-1994、GB/T2677.10-1995、GB/T2677.3-1993 测定。酸溶木质素和酸不溶木质素含量按照美国国家可再生能源实验室NREL/TP-510-4261 “Determination of structural carbohydrates and lignin in biomass”检测,杨木纤维素与半纤维素的测定采用高效液相色谱测定水解液(水解液来自酸不溶木质素制备过程)中葡萄糖与木糖的含量,以葡萄糖和木糖的标样作为标准测定含量。分别对WT 与转基因杨木进行水分、综纤、苯醇-抽提、klason 木质素、酸溶木质素、半纤维素、纤维素、灰分等组分进行测定。筛选优良转基因株系进行进一步实验。
1.3 转录组数据分析
将杨木试样送至生工生物工程 (上海) 股份有限公司进行cDNA 文库构建和转录组测序,从中找寻ERF76 基因(Potri.005G195000.1)并使用TBtools 软件分析。对比转录组数据野生型与ERF76 转基因杨木的差异基因。
1.4 半纤维素抽提
将实验所用6 年生小黑杨杨木自然风干一段时间后去除树皮,通过球磨机制备尺寸为40-60 目的杨木木粉,将合格木粉通过苯-醇抽提脱脂,并根据国标GB/T462-2008 测量木粉水分后备用。称取1g(绝干、精确0.0001)脱脂木粉放置于锥形瓶中,用20 mL 浓度为100 g/L 的氢氧化钠溶液与之充分混合,并处于室温下反应(0.5-3 h)反应结束后通过恒重的G2 玻沙漏斗分离固液相。
固相用去离子水洗涤至滤液为中性后放入(105±2℃)烘箱内烘至绝干,称重计算固相得率。液相使用冰醋酸调节液体pH 值为5.5 并加入3 倍体积(60 mL,浓度95%)的乙醇溶液。等待完全沉淀后再次固液分离,用70%的乙醇溶液洗涤并风干称量半纤维素质量并计算半纤维素提取率。
1.5 碱法组分分离
将实验杨木(6 年生)静置一段时间自然风干水分后去除树皮分别裁剪制备长2 mm 宽1 mm 高30 mm的木杆。将5g 杨木杆分别用用碱量为20、30(NaOH 计)的氢氧化钠,以液比为1∶6 的配比放在玻璃瓶中混合并充分搅拌。封盖后将盛有溶液的玻璃瓶转移到高压灭菌锅中,预设灭菌锅反应时间90 min 温度120℃。静待反应结束后采用布氏漏斗过滤,分离固液相。固相产物用去离子水清洗直至产物滤液变为中性为止。收集固体产物并放入(105±2℃)烘箱中烘干并记录得率数据,烘干原料经球磨机粉碎,并筛选40~60 目木粉用于检测残余木质素含量(参照1.2)每种处理包含3 次重复。
2 结果与分析
2.1 组分分析
由(表1)可知,转基因株系在纤维素、半纤维素、木质素等化学组分含量上发生了变化。由此可以判断过表达ERF76 基因对小黑杨木材组分造成了一定影响,且由于转基因株系的不同表现出了不同的结果。通过转录组数据分析得知ERF76 基因在不同小黑杨株系中的表达倍数,将其与木材各个化学组分定量结果相互比对发现,当FC 倍数相对较低时ERF76 过表达小黑杨的纤维素占比有所提高。当FC 倍数进一步提高后纤维素占比含量逐步下降直至出现负增长;半纤维素的含量占比与FC 变化的规律与纤维素相反,木质素含量占比的变化规律与半纤维素相同,虽然这一规律在木质素组分变化上有所体现但总体上对木质素占比含量影响并不是很大,变化的规律主要体现在纤维素与半纤维素的变化之上。

表1 两年生杨木各化学组分含量表Table1 Biennial poplar content table of each chemical component
经过比较P1、P2、P3 株系与WT 在纤维素的占比差的绝对值分别为1.25、0.37、0.06;半纤维素的占比差的绝对值分别为1.06、0.22、0.21;根据比较结果P1 的化学组分变化较其它株系更加明显,因此选定P1 株系进行进一步的研究。
由(表2)可知经历过4 年生长发育,由ERF76 基因的过表达木材对化学组分的影响在P1 株系体内不断累积并增大。与WT 纤维素含量的差值由四年前的1.25 变为3.05; 半纤维素的差值由1.06 变为3.16;四年生长发育过程中小黑杨木质化水平不断提高,但WT 与P1 的木质素水平差距不大。

表2 6 年生杨木各化学组分含量表Table2 Six-year-old poplar content table of various chemical components
由转录组数据通过TBtools 软件分析并结合(表3)可知ERF76 过表达转基因杨树与野生型杨树在549个基因表达上有所差异。其中上调基因195 个,下调基因354 个,其中影响转基因杨树碳水化合物合成的差异基因共27 个上调基因7 个(ID:Potri.002G006200.2;Potri.003G122000.1;Potri.004G071600.1;Potri.005G195000.1;Potri.007G015500.2;Potri.008G132500.1;Potri.010G201500.2),下调基因 20 个(ID:Potri.001G192200.2;Potri.002G060500.1;Potri.002G089800.1;Potri.002G089800.4;Potri.002G145300.1;Potri.005G201200.2;Potri.006G140300.3;Potri.008G055900.2;Potri.008G120000.2;Potri.008G123500.1;Potri.008G192600.1;Potri.008G219200.1;Potri.009G060800.4;Potri.010G010500.1;Potri.010G063100.1;Potri.014G094800.1;Potri.014G116800.1;Potri.016G025500.2;Potri.016G054600.2;Potri.016G075700.4;Potri.017G073200.3)由此为转基因杨木纤维素、半纤维素组分含量发生变化提供了分子层面的的依据。

表3 ERF76 杨木转录组差异基因统计表Table3 Transcriptome differential gene statistics table
2.2 半纤维抽提&碱法处理
针对P1 株系木材化学组分的特点,文章对该木材的工业化应用做了一些探究,对P1 转基因小黑杨进行了碱法工艺和半纤维素提取工艺[18]的进一步研究,研究相同工艺下WT 与ERF76 转基因杨树(P1 株系)的不同。实验结果如下:
根据报道,室温(25℃)下100 g/L 的氢氧化钠只能溶解聚合度较低的半纤维素[19]。结合(表4)和(图1)所示结果可以得出以下结论:从氢氧化钠抽提结果上看,在相同的处理条件下P1 脱除半纤维素的效率明显高于WT。由于该工艺条件针对于聚合度低的半纤维素进行抽提,且P1 的半纤维素脱除率较高,因此P1 所含低聚合度的半纤维素在总半纤维素中占比相对较高。由此侧面证明了P1 植株的半纤维素合成途经受到了ERF76 基因过表达的影响。同时通过实验平衡时间与半纤维素脱除效率,找寻到了ERF76 过表达小黑杨木材在室温 (25℃)氢氧化钠提取液浓度为100 g/L 提取半纤维素的最适时间为90 min。

图1 半纤维素脱除率Figure1 Hemicellulose removal rate
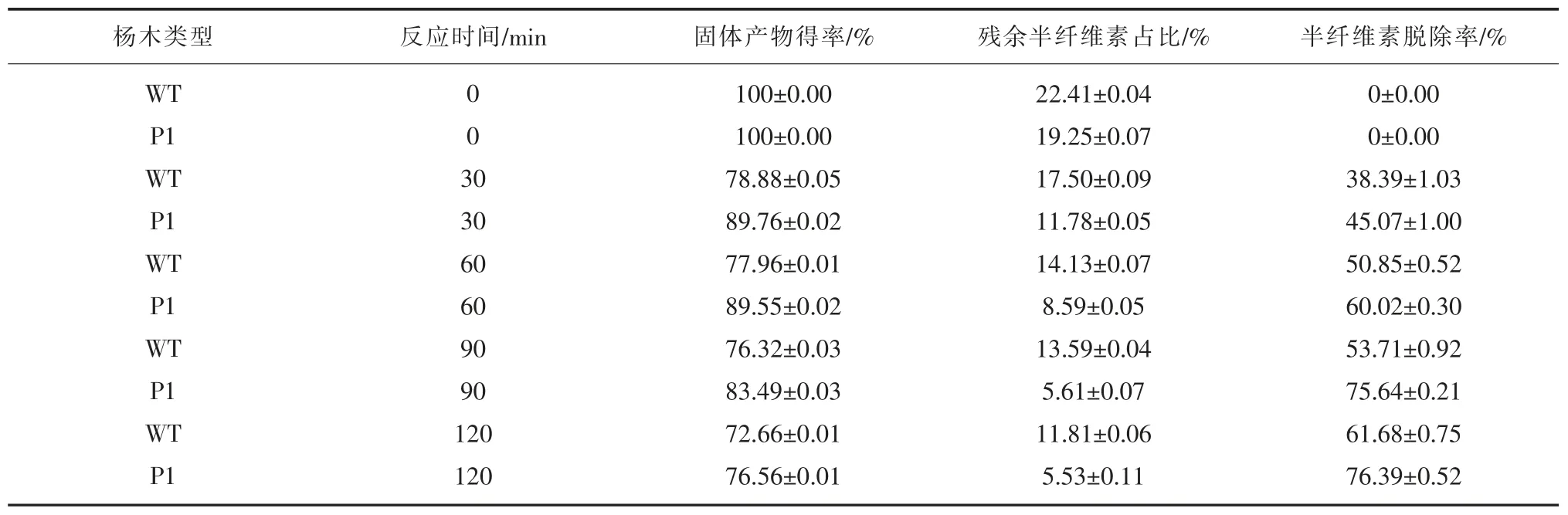
表4 半纤维素抽提结果表Table4 Hemicellulose extraction results table
正如(表5)所示结果发现,在相同的碱处理工艺下P1 与WT 木材在脱除木质素方面的差别不大,但P1 可以保持较高的制浆得率。提高反应工艺的用碱量后转基因木材仍旧维持着上述结果。造成这一结果的原因主要应有两点,第一转基因株系的和WT 纤维素组分含量、半纤维素组分含量存在差异(详见表2)。由于反应温度并未超过150℃,因此反应釜内的主要化学反应应为剥皮反应[20]。根据上述半纤维素抽提所得结论,P1 含有大量聚合度较低的半纤维素。因剥皮反应的特性是从纤维素、半纤维素链条的末端依次反应降解,且半纤维素含有大量支链更易降解。因此WT 与P1 在反应之初半纤维素快速溶出并降解,而纤维素反应较慢,此时木材纤维素组分上的不同成为了得率差异的主要原因。第二从反应接触面积来看,半纤维素的降解会为反应液的侵入提供更多的接触孔隙。由于P1 与WT 在半纤维素组分含量上的差异(详见表2)P1 较少的半纤维素含量,致使反应过程中WT 更易与反应液接触促进碳水化合物的不断降解。

表5 固体产物的得率、木质素分析表Table5 Table of yield and lignin analysis of solid products
3 结论
通过研究多年生过表达ERF76 小黑杨的化学组分变化为人们进一步了解ERF76 基因对木材组分方面的影响提供了相关依据。通过对两年生WT 与不同ERF76 过表达转基因株系木材化学组分的分析和转录组数据分析,揭示了ERF76 基因过表达倍率不同对木材各组分的影响。当FC 增高时纤维素组分占比曾高,半纤维素含量占比较低;当FC 进一步提高时半纤维素含量占比得到逐步提高,纤维素占比下降。根据实验结果选出化学组分来变化较为明显的ERF76 转基因株系(P1),通过进一步检验六年生WT 与P1 植株的木材化学组分数据,观察到由ERF76 基因过表达造成的纤维素、半纤维素组分影响在植物生长发育过程中不断累积。
针对ERF76 基因对纤维素、半纤维素组分影响较大的特点,对转基因小黑杨木材中相关纤维素、半纤维素材料方面进行了低聚合度半纤维素的提取和碱法处理工艺处理的进一步的探究。研究发现ERF76过表达的小黑杨木材中低聚合度的半纤维素占据其半纤维素总量的大部分。这为探究ERF76 基因影响植株低聚合度半纤维素合成路径提供了一定的思路。在碱处理实验过程中通过对反应温度的限制,利用剥皮反应为主要反应探究化学组分变化在比较温和的碱性反应过程中带来的影响。具体表现为转基因株系(P1)相较WT 存在高纤维素低半纤维素的特点。在剥皮反应中半纤维素含量低反应快、纤维素含量高反应慢,最终导致了P1 高得率结果。半纤维素提取结果上找到了转基因木材在室温下氢氧化钠浓度为100 g/L 时最适提取时间为90 min。
猜你喜欢
关东学刊(2022年3期)2022-10-14 11:48:52
亚热带植物科学(2022年1期)2022-05-17 12:39:32
上海包装(2019年8期)2019-11-11 12:16:14
四川农业科技(2019年5期)2019-07-01 09:46:46
江河文学(2018年4期)2018-11-12 21:01:51
湖南林业科技(2017年1期)2017-02-06 05:29:05
天津造纸(2016年1期)2017-01-15 14:03:29
浙江柑橘(2016年1期)2016-03-11 20:12:31
中国造纸学报(2015年1期)2015-12-16 19:35:25
中国造纸(2015年7期)2015-12-16 12:40:46
