宁夏枸杞铁镉响应基因的筛选及其功能验证
2023-08-15 08:30余慧王静梁昕昕辛亚平周军赵会君
生物技术通报 2023年7期
余慧 王静 梁昕昕 辛亚平 周军 赵会君
(北方民族大学生物科学与工程学院 国家民委黄河流域农牧交错区生态保护重点实验室,银川 750021)
宁夏枸杞(Lycium barbarumL.)为茄科枸杞属多年生落叶灌木,其果实枸杞子富含多糖、甜菜碱以及黄酮类化合物,具有滋补肝肾、润肺明目、抗氧化、抗衰老等功效[1]。宁夏枸杞是宁夏当地典型的经济树种,同时也是盐碱地栽植的先锋物种之一,在甘肃、青海、新疆、内蒙古和西藏等地也有大面积种植[2]。
铁元素(Fe)是植物生长发育过程中所必需的微量元素,广泛参与光合作用、呼吸作用、叶绿素生物合成、活性氧的形成与消除以及氮素的固定等关键生理代谢过程[3],虽然土壤中含有丰富的Fe,但其主要以三价铁形式存在,植物无法直接吸收利用[4],尤其西北地区土壤pH普遍偏高[5],在盐碱地种植的枸杞往往会出现缺铁失绿性黄叶病,尤以幼苗和幼树受害严重,这导致幼苗叶绿素合成受阻,含量降低,新叶变黄,生物量大幅度下降[6]。还有研究表明,Fe的缺乏会导致植物过量地吸收镉(Cd)[7],Cd是毒性很高的重金属元素,一般通过农艺措施进入环境,Cd通过Fe和锌(Zn)转运蛋白进入植物体内[8],降低叶绿素的含量以及影响植物体碳的固定从而对植物造成毒害,影响品质[9]。
近年来,FRO、ZIP和NRAMP家族中的一些成员参与了植物细胞内Fe和Cd的吸收和转运[10-11]。铁还原酶基因FRO能够将土壤中的三价铁转换成二价铁。迄今为止,在拟南芥中已克隆出8个铁还原酶基因AtFRO1-AtFRO8,其中,AtFRO2、AtFRO4及AtFRO5在根系中发挥作用[12-13],而AtFRO6、AtFRO7及AtFRO8主要在叶片、茎段、花以及果实中高效表达[14-16]。除了拟南芥,豌豆和番茄等植物根系中的重要三价铁还原酶基因也相继被克隆。豌豆Fe(Ⅲ)还原酶基因PsFRO1在根及叶中表达,该基因受缺铁信号诱导而在根表皮高效表达[17]。番茄Fe(Ⅲ)还原酶基因LeFRO1在根、叶、花、果中表达[18]。金属离子转运蛋白IRT1是ZIP基因家族中重要的成员,拟南芥的AtIRT1不仅能转运Fe,而且还能转运锰(Mn)、锌(Zn)以及Cd等金属元素,将AtIRT1敲除后的突变体在缺铁胁迫下叶片发黄,在苗期致死[19]。缺铁条件下,水稻OsIRT1和OsIRT2能增强对Cd的摄取和转运[7]。除了铁转运蛋白IRT1能运输二价铁离子之外,YSL、NRAMP以及OPT等基因也参与了Fe2+的转运过程[20]。金属转运家族Nramp家族不仅能吸收和转运Mn、Fe等微量元素,还参与了植物对Cd等重金属的吸收和转运[21],越来越多的研究证实,Fe和Cd共用离子通道[22]。
与其他植物相比,宁夏枸杞中Fe和Cd吸收转运相关的基因研究较少。本研究通过转录组测序技术及生物信息学分析技术,筛选与Fe和Cd吸收转运相关的关键基因,利用实时荧光定量PCR技术结合转基因酵母技术进行基因功能初步验证,深入探讨目的基因在Fe离子吸收及稳态维持中的作用以及在Cd吸收转运中的功能,为宁夏枸杞黄叶病的防治、培育高效Fe利用枸杞品种和低镉累积品种的培育提供理论基础及技术支持。
1 材料与方法
1.1 材料
以宁杞1号组培快繁苗为材料,取生长良好、高度基本一致的组培苗移入Hoagland营养液中进行水培,每7 d更换一次营养液,培养条件为28℃光照16 h/22℃黑暗8 h,光照强度4 000 lx,相对湿度60%-70%。待水培苗生长至10 cm左右时进行处理,用于转录组测序及RNA提取。
1.2 方法
1.2.1 转录组测序及与金属离子吸收转运相关基因的筛选 选取生长健康、大小一致的枸杞幼苗,以未经Cd处理的枸杞水培苗作为对照,用100 μmol/L的CdCl2处理12 h和48 h后,分别收集对照组和处理组的叶片和根系组织,每个样品重复取样3次,液氮速冻后送深圳华大公司进行转录组测序。利用GO(Gene Ontology)数据库和KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库进行注释[23],挖掘Cd转运相关功能基因,利用热图聚类分析筛选与Cd吸收转运相关的转运蛋白基因并进行后期验证。
1.2.2 枸杞组织RNA的提取及cDNA合成 选取生长健康、长势基本一致的枸杞幼苗,部分幼苗经200 μmol/L Fe-EDTA处理,于处理后第0、2、8、12、24、48和72 h收集枸杞的根系组织并液氮速冻;剩余部分经100 μmol/L CdCl2处理后第0、8、12、24、48和72小时收集枸杞的根系组织,液氮速冻,用于RNA的提取。取0.1 g枸杞根系组织于液氮中充分研磨成粉末后,利用RNAprep Pure Plant Kit试剂盒(北京,天根)提取叶片总RNA,1.0%琼脂糖凝胶电泳检测RNA的完整性。以总RNA为模板,用逆转录试剂盒逆转录合成cDNA(北京,全式金),用作实时荧光定量PCR的模板。
1.2.3 实时荧光定量PCR分析Fe、Cd胁迫下目的基因表达变化模式 为进一步研究与Fe离子吸收转运相关的基因在Fe、Cd胁迫下的表达变化情况,筛选了5个相关基因,并使用primer express 3.0软件设计实时荧光定量引物(表1),采用美国MX3000pTMqPCR实时荧光定量PCR仪,以宁夏枸杞的β-actin作为内参基因[24],SYBR Green Universal Master mix kit(Toyobo, Osaka, Japan)为荧光染料,模板为稀释15倍的cDNA,每个样品设3个重复,反应程序为95℃ 10 min;95℃ 30 s,60℃ 30 s,72℃ 30 s,45个循环。采用2-△△CT法计算基因相对表达量。

表1 基因克隆以及特异性表达分析引物表Table 1 Primers for gene cloning and specific expressions
1.2.4 枸杞中与Fe、Cd吸收转运相关基因克隆及生物信息学分析 利用常规PCR法,以cDNA为模板,克隆基因全长序列(表1),测序,blast比对并确认。利用在线工具(https://web.expasy.org/protparam/)预测目的基因分子量;利用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)预测蛋白质跨膜结构域;利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测蛋白质的二级结构;利用swiss-model(https:// www.swissmodel.expasy.org/)预测蛋白质的三级结构;使用MEGA 7.0软件的Neighbor-Joining(邻接)法构建系统发育树。
1.2.5 酵母表达载体的构建及转基因酵母功能分析 所用酵母为Fe敏感型酵母菌株(△CCC1)及Zn/Cd/Ni/Co敏感型酵母菌株(YK44),构建YES2-IRT2、YES2-FRO以及YES2-NRAMP3酵母表达载体并测序验证。
Fe的敏感型转化试验:将Fe敏感型菌株△CCC1接种于YPDGAL液体培养基中进行活化培养,采用醋酸锂法制备△CCC1酵母感受态细胞,将YES2-IRT2、YES2-FRO、YES2-NRAMP3以及YES2空质粒分别导入△CCC1感受态细胞,在CMUracil和SD+His的筛选培养基上筛选阳性重组酵母菌株并进行PCR验证。配制含0、400、600和800 μmol/L FeSO4·7H2O的YPD固体培养基。
Cd的敏感型转化试验:将Zn/Cd/Ni/Co敏感型酵母菌株(YK44),接种于YPDGAL液体培养基中进行活化培养,采用醋酸锂法制备YK44酵母感受态细胞,将YES2-IRT2、YES2-FRO、YES2-NRAMP3以及YES2空质粒分别导入YK44感受态细胞,在CM-Uracil和SD+His的筛选培养基上筛选阳性重组酵母菌株并进行PCR验证,配制含0、40、60和100 μmol/L CdCl2的YPD固体培养基。
将重组酵母菌株接种到YPDGAL液体培养基中,待OD600达到0.5-0.6时停止培养,用灭菌水分别稀释10-1、10-2、10-3和10-4倍,各取2 μL依次接种于上述含有重金属的YPD固体培养基中,30℃倒置培养72 h,观察酵母生长状况,每个试验处理重复3次。
2 结果
2.1 转录组数据中与金属离子吸收转运相关基因的筛选及其注释
以无Cd胁迫的宁夏枸杞为对照组(CK),以100 μmol/L CdCl2胁迫下的宁夏枸杞为处理组进行转录组测序,通过GO和KEGG分析12和48 h的根系和叶片转录组数据,设置log2≥2为筛选条件,筛选到大量受Cd诱导而上调表达的基因(图1、表2),高效响应金属离子的跨膜转运蛋白主要包括:ZIP跨膜转运蛋白家族、NRAMP跨膜转运蛋白家族、YSL跨膜转运蛋白家族、OPT跨膜转运蛋白家族基因等,表明这些跨膜转运蛋白家族的基因能积极地响应Cd的胁迫,可能具有转运金属离子的作用。
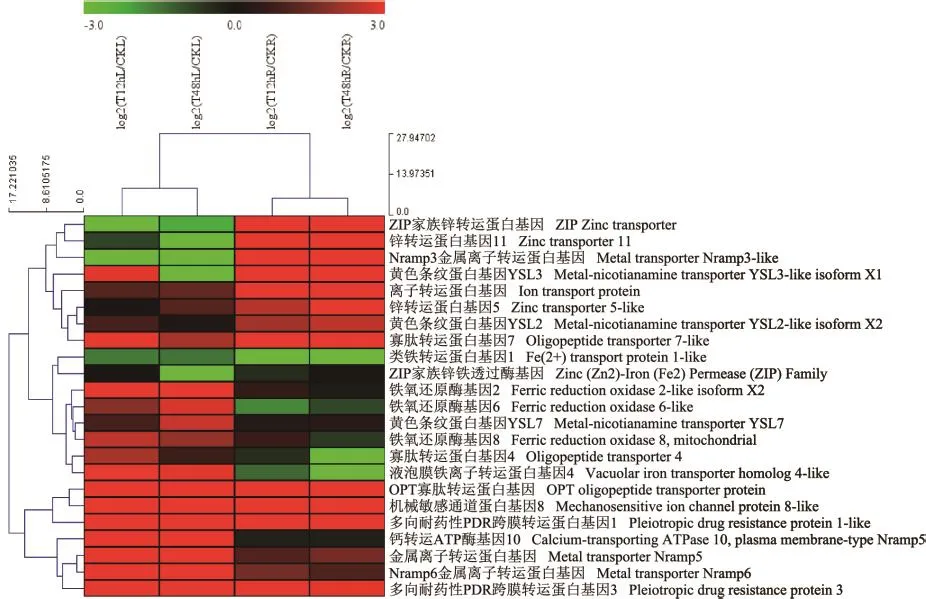
图1 宁夏枸杞根系及叶片中离子跨膜转运蛋白基因的筛选及热图聚类分析Fig.1 Screening and hot map of ion transporter genes in the leaf tissue and root tissue of L.barbarum
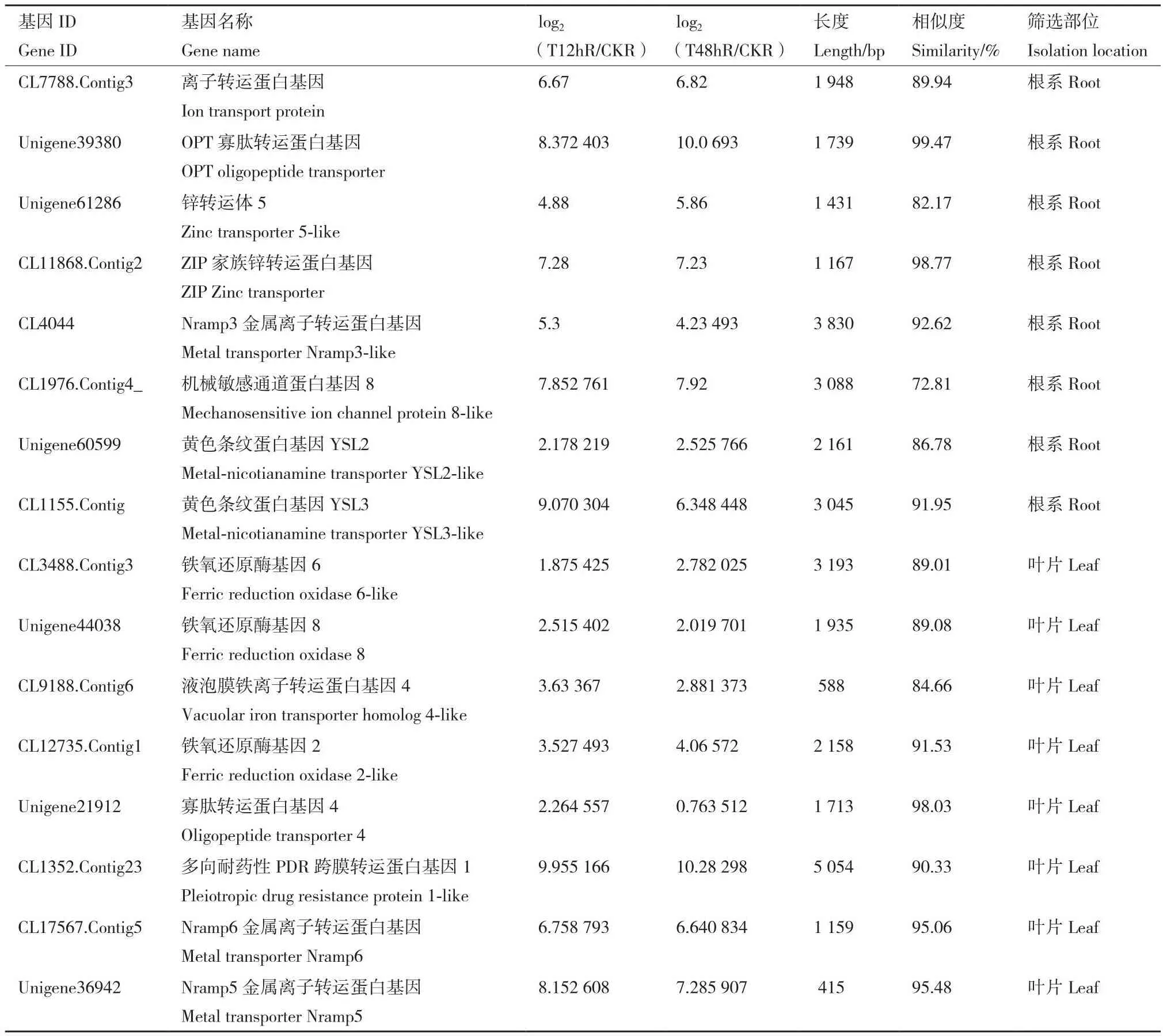
表2 根系及叶片中筛选的离子转运相关基因Table 2 Genes related to ion transporter selected from the root and leaf
2.2 Fe、Cd胁迫下目的基因表达模式分析
基于转录组数据分析,筛选5个与跨膜转运蛋白基因进行Fe、Cd胁迫下的表达模式分析,其在200 μmol/L Fe-EDTA胁迫下的表达倍数(图2)。IRT1在Fe胁迫8 h时开始上调表达,随着处理时间的延长而增加,在12 h表达量达到极显著水平(P<0.001),在24 h表达量达到最大值,是对照的4.37倍;IRT2在胁迫2 h开始显著上调表达,在48 h表达倍数达到极显著水平(P<0.001),是对照的4.13倍;FRO在Fe胁迫初期表达量略有下降,随着时间的延长逐渐增加,在72 h时,表达量达到极显著水平(P<0.001),为对照的3.28倍;NRAMP3和YSL在Fe胁迫后的72 h时却呈现极显著下降水平。结果表明,这些基因在高铁胁迫下呈现不同的表达变化模式,甚至同一基因在不同的时间点表达倍数有明显的差异,推测这些基因在枸杞中参与Fe元素的吸收和转运。
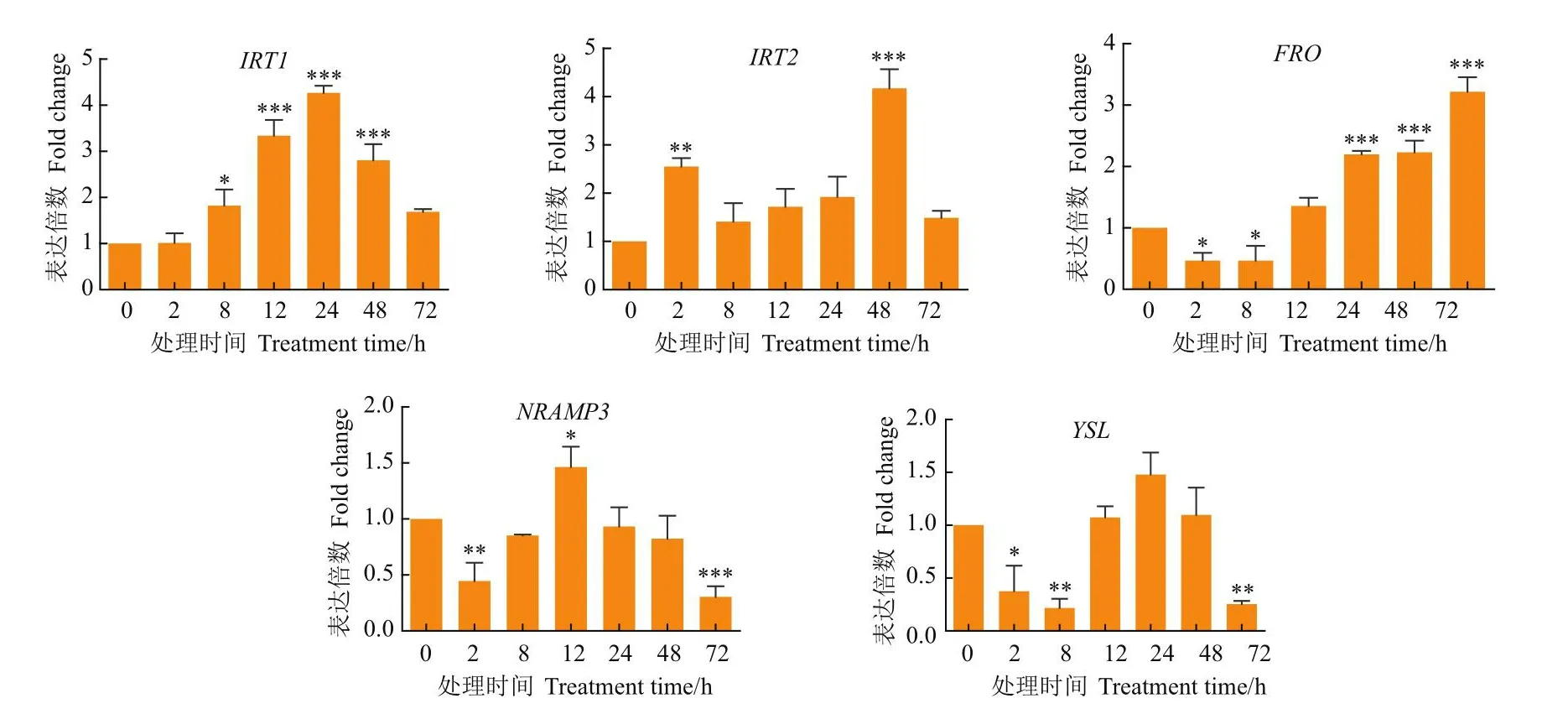
图2 目的基因在200 μmol/L的Fe-EDTA胁迫下的表达变化模式Fig.2 Expression pattern of target gene under 200 μmol/L Fe-EDTA stress
IRT1在Cd胁迫下8、48及72 h表达倍数达到显著水平(P<0.01);IRT2在Cd胁迫初期表达量并未上调反而有所下降,在第48小时显著上调表达(P<0.05),在72 h表达倍数达到极显著水平(P<0.001),是对照的5.04倍;FRO的表达量在8 h开始极显著被抑制(P<0.001),48 h恢复表达水平;NRAMP3在72 h表达量达到最大值,为对照的6.06倍,YSL为对照的3.26倍(图3)。结果显示,FRO受到Cd的负调控,其余的基因都受到Cd的诱导表达,表达倍数有明显的差异,推测这些基因可能参与了枸杞对Cd的耐受、转运或解毒。
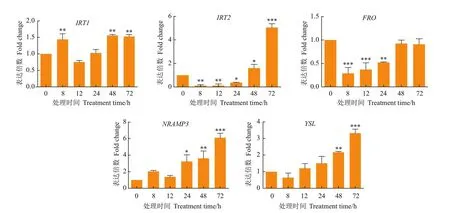
图3 目的基因在100 μmol/L的CdCl2胁迫下的表达变化模式Fig.3 Expression pattern of target gene under 100 μmol/L CdCl2 stress
2.3 LbIRT2、LbFRO以及LbNRAMP3的克隆及生物信息学分析
根据实时荧光定量PCR结果筛选了IRT2、FRO及NRAMP3为目的基因进行全长序列的克隆及后期基因功能验证,利用PCR技术克隆并进行生物信息学分析以及转基因酵母功能分析。以宁杞1号cDNA为模板,采用常规PCR法,克隆到IRT2、FRO及NRAMP3全长序列,分别为645、2 220和1 533 bp。
通过在线工具预测这3个基因的理化特征(表3),LbIRT2和LbFRO编码的蛋白理论等电点均大于7,属于碱性蛋白;LbNRAMP3编码的蛋白属于酸性蛋白。LbIRT2编码的蛋白不稳定指数大于40,属于不稳定蛋白;而LbFRO和LbNRAMP3编码的蛋白属于稳定蛋白。用SOPMA软件分析蛋白的二级结构,结果(表4)显示,LbIRT2和LbNRAMP3的α-螺旋含量约占55%,LbFRO蛋白约占37%;其次是无规则卷曲,占20%-30%,β-转角含量最少。LbFRO和LbNRAMP3三级结构主要由α-螺旋构成,与二级结构预测结果相符合(图4)。
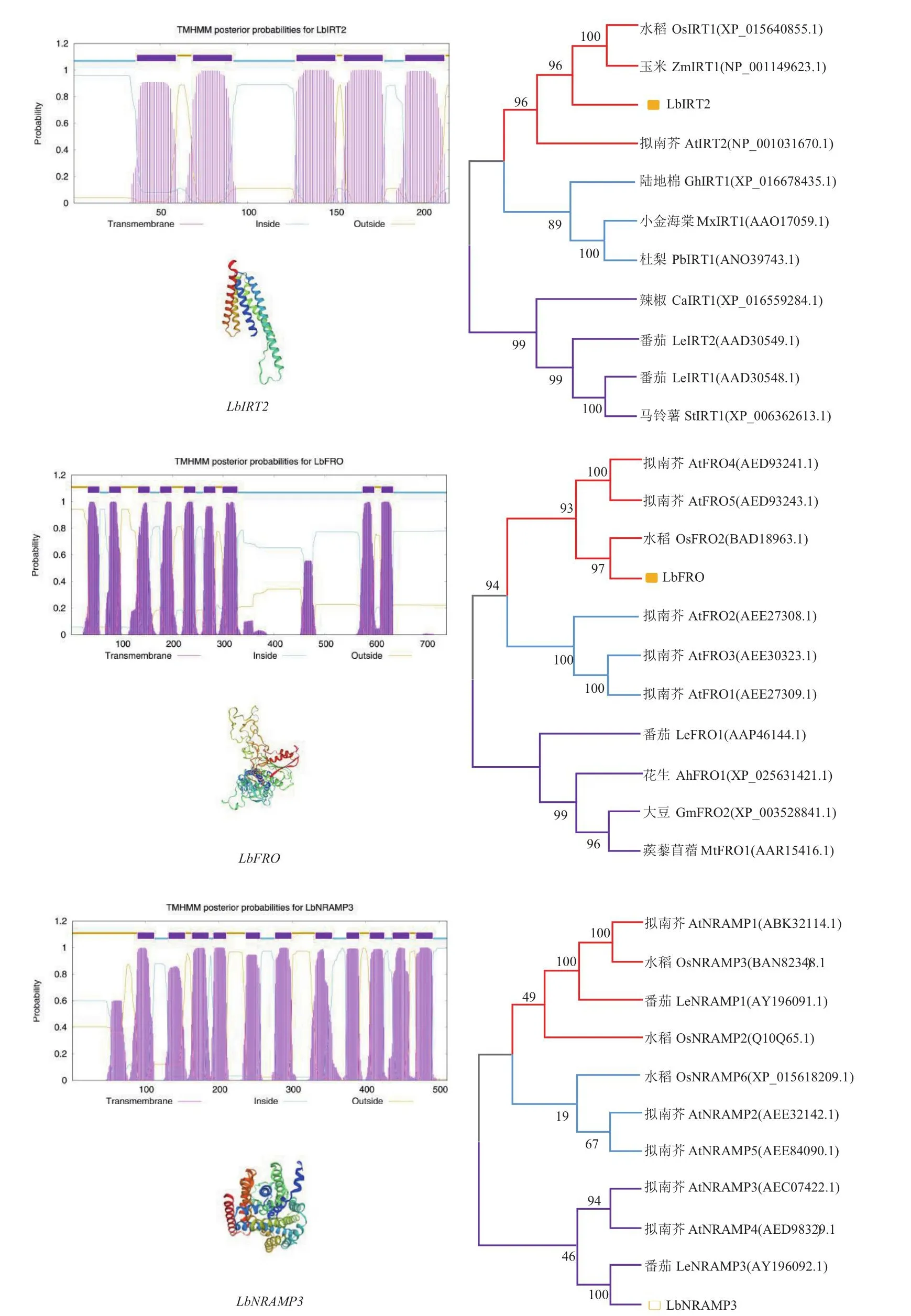
图4 LbIRT2、LbFRO及LbNRAMP3跨膜结构、蛋白三级结构以及进化树分析Fig.4 Transmembrane structure, tertiary structure and phylogenetic tree analysis of LbIRT2, LbFRO and LbNRAMP3 genes
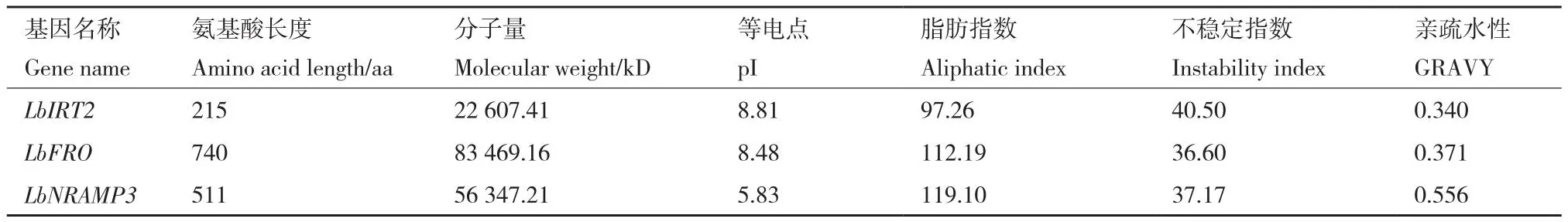
表3 宁夏枸杞中响应铁镉胁迫的功能基因理化性质分析Table 3 Physicochemical properties of functional genes responsible to iron and cadmium stress in L.barbarum
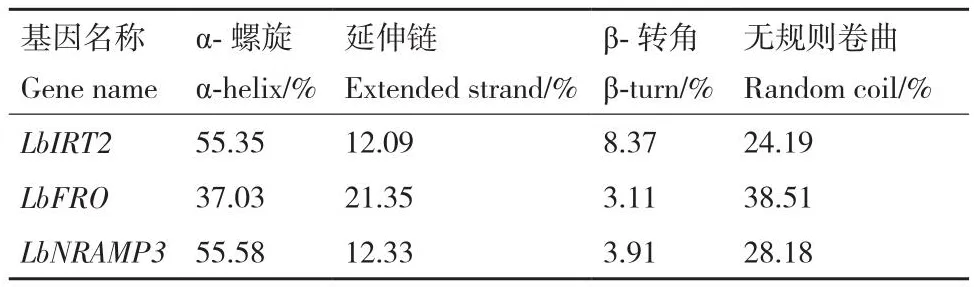
表4 宁夏枸杞中响应铁镉胁迫的功能基因二级结构预测Table 4 Prediction of secondary structure of functional genes in L.barbarum response to iron and cadmium stress
跨膜结构域预测显示,LbIRT2具有5个跨膜结构域;LbFRO具有9个跨膜结构域;LbNRAMP3具有11个跨膜结构域,符合NRAMP3家族成员的10-12个典型跨膜螺旋的结构特征。
聚类分析显示,LbIRT2和玉米以及水稻的IRT1聚为一类,和拟南芥的AtIRT2亲缘关系较近;LbFRO和水稻的OsFRO2同源性最高和拟南芥的AtFRO4和AtFRO5处在同一分支;LbNRAMP3与番茄的LeNRAMP3具有较高的相似性,和拟南芥的AtNRAMP3和AtNRAMP4处在同一分支。
2.4 转基因酵母功能分析
通过分析不同浓度Fe及Cd处理下LbIRT2、LbFRO以及LbNRAMP3的转基因酵母(图5)。随着FeSO4·7H2O浓度的增加,3个转基因酵母菌株生长状况要优于对照,在600和800 μmol/L处理下,能明显观察到转化YES2-IRT2、YES2-FRO以及YES2-NRAMP3的酵母菌生长状况比对照好;在40和60 μmol/L的CdCl2胁迫下,YES2-IRT2和YES2-NRAMP3转基因酵母的长势均弱于对照组,在100 μmol/L的CdCl2处理YES2-NRAMP3酵母菌株生长受到极显著抑制,而YES2-FRO转基因酵母在CdCl2处理下的生长状况优于对照组。
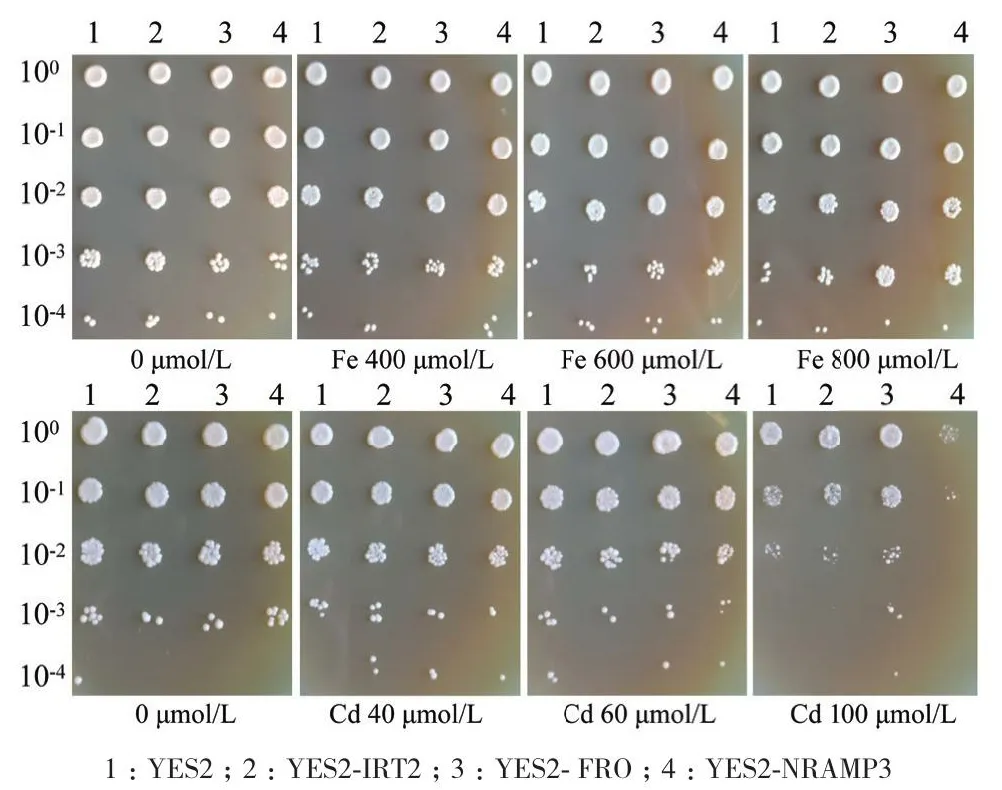
图5 转基因酵母的表型分析Fig.5 Phenotypic analysis of transgenic yeast
3 讨论
大量文献报道,FRO、ZIP和NRAMP家族中的一些成员参与了植物体内Fe和Cd的吸收转运过程。铁还原酶基因(FRO)在植物对Fe元素的吸收上起着重要的作用,能够将Fe3+还原成Fe2+[25],使植物能更加容易地吸收铁元素。Satbhai等[26]利用转基因技术将AtFRO2导入其他植物体,能够明显减轻植物缺铁黄化症状,番茄LeFRO1在叶片中的稳定表达,维持了植物对Fe的吸收利用[27],杜梨幼苗中与Fe吸收相关基因FRO2和IRT1在缺铁胁迫下显著上调表达,增加了对Fe的吸收利用[20],苹果属小金海棠MxFRO4和MxFRO6定位于细胞质膜上,在根、茎、叶中均有表达,受到缺铁和高铁胁迫的诱导表达[28],水稻OsFRO2表达可由铁过量或持续性缺铁诱导[29],这显示FRO基因受缺铁或高铁诱导表达,参与了植物对铁的吸收利用。本研究所构建的LbFRO系统进化树显示,LbFRO和水稻OsFRO2同源性最高,受到高铁诱导表达,这与Ishimaru等[30]研究结果十分相似,充分证实了FRO基因参与了枸杞中铁离子的吸收和转运。同时,该基因的过表达能增强YES2-FRO转基因酵母对Fe和Cd的耐受性,其机理有待于进一步的研究。
ZIP转运蛋白家族的基因不仅能够转运植物生长所必需的微量元素Zn、Fe、Mn和Cu等,同时还能转运Cd、镍(Ni)、钴(Co)等有毒元素[31]。AtIRT2是最早被发现并克隆到的ZIP家族成员[32],AtIRT2与AtIRT1有很高的功能相似性,AtIRT2已经被证实定位在细胞内的囊泡上,在根尖受低铁诱导表达;AtIRT1突变体在环境中无法正常生长,叶片严重黄化,幼苗期因缺铁致死。水稻中的OsIRT1在根、柄和叶中均有表达,过表达OsIRT1基因会提高水稻全株的Fe、Zn含量,并且对高浓度的Cd元素非常敏感,这表明OsIRT1还参与了Zn、Cd的吸收及转运。本研究转基因酵母对Fe、Cd的耐性分析表明,在高铁胁迫下LbIRT2的转基因酵母长势比空载体好,由于LbIRT2与定位在囊泡上的AtIRT2同源性较高,推测该基因可以将Fe快速螯合到囊泡中来减少毒害,这种现象被称为主动适应环境变化[33];Nakanishi等[7]发现转化水稻OsIRT1和OsIRT2的酵母菌较表达空载体的酵母对Cd的敏感性更强,通过检测发现转OsIRT2酵母细胞中Cd含量显著高于对照组,大约是对照的1.8倍,表明转OsIRT1和OsIRT2的酵母菌株累积了过量的Cd而导致对Cd胁迫更加敏感。本研究进化树分析显示LbIRT2与OsIRT1同源性较高,同时转化LbIRT2的酵母生长受到了抑制,这与Nakanishi等[7]的研究结果一致,推测LbIRT2可能参与了枸杞对Fe和Cd的转运,有待于后期深入研究。
Nramp家族是一类具有转运金属离子功能的膜蛋白家族,不仅能转运Fe和Mn等营养元素,还参与了植物对Cd的吸收和转运。进化树分析显示,LbNRAMP3与番茄的LeNRAMP3基因以及拟南芥的AtNRAMP3亲缘关系较近。LeNRAMP3已经被证实定位在囊泡中,在根系中高效表达,在Fe的获取和再分配中发挥作用[34];AtNRAMP3定位在液泡膜上,在叶和根中高效表达,在酵母细胞中表达AtNRAMP3和AtNRAMP4能够提高酵母对Cd的敏感性,通过测定培养24 h后的转基因酵母中Cd的含量发现,转AtNRAMP3和AtNRAMP4的酵母中Cd含量显著高于对照,分别大约是对照的1.8倍和1.7倍[35-38]。本研究进化树分析显示,LbNRAMP3与AtNRAMP3和AtNRAMP4序列高度相似,酵母转化试验结果显示,转化LbNRAMP3的酵母在含40和60 μmol/L CdCl2的培养基上的长势相对于对照组较弱,在100 μmol/L CdCl2处理的培养基上无法生长,表明转化LbNRAMP3提高了酵母对Cd的敏感性。同时实时荧光定量PCR发现LbNRAMP3在Fe、Cd胁迫下明显上调表达,以上研究结果证明了LbNRAMP3参与了Fe和Cd的转运。
4 结论
从宁夏枸杞中克隆得到LbIRT2、LbFRO以及LbNRAMP3,其受Fe和Cd胁迫后显著上调表达,酵母中表达LbIRT2、LbFRO以及LbNRAMP3能够增强酵母对Fe和Cd的敏感性,这3个基因可能参与了枸杞对Fe和Cd的吸收转运。
猜你喜欢
学与玩(2022年2期)2022-05-03
军事文摘·科学少年(2021年1期)2021-02-04
医药前沿(2020年28期)2020-12-02
绿色中国(2019年13期)2019-11-26
阅读(低年级)(2019年9期)2019-11-15
心肺血管病杂志(2019年1期)2019-04-22
基层中医药(2018年10期)2018-12-06
教学考试(高考生物)(2017年4期)2017-12-13
阅读与作文(小学低年级版)(2015年8期)2015-05-30
食品工业科技(2014年9期)2014-03-11