
不同补光时段对夏黑葡萄生长及光合特性的影响
2023-08-14 06:34王壮伟吴伟民王博王西成闫莉春
江苏农业科学 2023年13期
王壮伟 吴伟民 王博 王西成 闫莉春
摘要:通过探究不同补光时段对设施栽培夏黑葡萄生长和光合特性的影响,明确利于设施葡萄生长和树势营养的最佳补光时段,为葡萄设施栽培提供理论支撑。以5年生夏黑为试验材料,在大棚避雨生产模式下,采用以红光为主波段的红蓝光LED灯作为光源,设T1(05:00—21:00)、T2(05:00—09:00)、T3(17:00—21:00)3个不同时段的补光处理,测定葡萄新梢生长、叶片营养、叶绿素含量、光合特性指标和果实品质。结果表明,设施栽培模式下,人工补光增加了夏黑葡萄新梢前期生长的粗度,缩短了节间长度,提高了叶片叶绿素含量,延缓了老叶叶绿素的降解,延长了叶片功能期,3个补光处理的嫩叶叶绿素含量与对照相差不大,而老叶的叶绿素含量都显著高于对照。补光处理增加了叶片中氮、钙、镁的含量,增加了叶片面积和干物质含量,T1处理果实单粒质量增加9.09%,果实品质与对照无显著差异。本研究中夏黑葡萄净光合速率日变化呈双峰型,高温是夏黑葡萄光合午休的主要原因;人工补光提高了葡萄光合作用的净光合速率、蒸腾速率、气孔导度、光饱和点,T1、T2处理的光合特征参数(除胞间CO2浓度外)均与对照呈显著性差异。综合比较叶片营养、新梢长势、果实品质、光合特性等各项指标,全天补光T1处理效果最好,早晨补光T2处理效果优于傍晚补光T3处理。
关键词:夏黑葡萄;补光;生长;光合特性
中图分类号:S663.104 文献标志码:A
文章编号:1002-1302(2023)13-0163-06
近年来葡萄设施栽培在我国发展迅速,成为我国目前葡萄生产的主要模式,葡萄设施栽培类型有促早栽培、延迟栽培、避雨栽培等多种形式,具有拉长果实供应期、扩大品种适应性、提高品质、减轻病害等显著效果。葡萄已成为我国设施栽培面积最大、技术最成熟的果树品种[1-2]。
葡萄为喜光作物,设施栽培通过薄膜覆盖达到保温、避雨的目的,然而由于受薄膜覆盖、薄膜老化、灰尘黏附等因素影响,也在一定程度上削弱了设施内环境的光照[3-4]。尤其是在早春和秋冬季节,光照度低、光照时间短、光照分布不均匀,蓝、紫和紫外等短波光线比例低,是设施园艺光环境的典型特点[5-6]。光照是影响设施园艺作物生长发育的主要环境因子之一,植物的形态建成、生理代谢、物质积累和基因调控等方面均与光环境有显著相关性。光照不足直接影响叶片的光合作用,影响植株生长和花芽分化,造成产量降低、品质变差、树势变弱等问题[7-9]。
人工补光是改善设施内光环境的有效措施。目前补光在设施园艺作物上的应用研究已有很多报道,通过人工补光改善设施内光环境,增强光照度、延长光照时间、调节光谱组成等,能提高作物光合能力,促进植株生长发育,提高产量,改善品质[10-12]。前人研究表明,一年两熟设施栽培模式下,夜间补光能提高巨峰葡萄春果叶片SPAD值、淀粉含量及全氮含量,显著增加产量,提高果实可溶性固形物含量和果皮花色苷总量;能增加冬果葡萄新梢粗度,加快果实膨大速率,提高品质[5,13-14]。阳光玫瑰葡萄冬果膨大期内夜间补光,可明显提高果實中吲哚乙酸(IAA)和赤霉素(GA3)的含量,降低脱落酸(ABA)含量,提高产量,增糖降酸,有效改善葡萄冬果的品质[15]。不同光质处理在设施葡萄生产上的报道也较多,有研究表明,补充红光、蓝光均明显增加温室葡萄新梢基部粗度,补充蓝光、紫外光减小了单叶面积,补充红光增加了新梢总的干物质积累[16]。夜间紫外光和蓝光灯照能够对“瑞都香玉”起到增糖降酸的作用,并且促进萜烯类物质合成[6]。
在实际生产中,尽可能地以较低的补光能耗达到较佳的补光效果,是设施葡萄补光措施应用的主要原则。本研究以夏黑葡萄为试材,进行避雨设施栽培,以设施农业上普遍使用的红蓝光为光源,设置不同时间段的补光处理,研究不同时段补光对葡萄新梢叶片生长、营养积累及光合性能的影响,以期初步确定葡萄生长发育对不同时段补光的响应特性,为设施葡萄光环境调控技术提供理论依据,筛选葡萄设施生产较佳的补光时段,实现低碳高效生产。
1 材料与方法
1.1 试验材料
试验于2021年在江苏省农业科学院葡萄园进行。供试葡萄品种夏黑,为定植6年的植株。树冠和长势一致,“一”字形棚架整形,株行距为2 m×3 m,南北行向,设施避雨根域限制栽培,花期新梢进行摘心处理,果穗整形每穗留70个果粒。
1.2 试验区概况
葡萄园建在江苏南京。南京属亚热带季风气候,年平均温度15.4 ℃,年极端气温最高39.7 ℃,最低-13.1 ℃。雨量充沛,年降水1 200 mm,春有梅雨时节,夏季亦高温多雨,降水主要在5—9月。
1.3 试验设计
光源使用以红光为主波段的红蓝光LED灯(红光650 nm、蓝光450 nm,红蓝光比例为3 ∶1),功率26 W。悬挂于树体种植行上方30 cm,灯距1.5 m,补光从芽绒球期开始,至成熟期结束。试验设计4个补光时段处理,分别是处理T1全天补光(05:00—21:00)、处理T2早晨补光4 h(05:00—09:00) 、处理T3傍晚补光4 h(17:00—21:00)、CK不补光。每3株树为1个小区,小区面积为18 m2(6 m×3 m),随机排列,每个小区设补光灯4盏,每个处理3次重复。每个处理间用反光膜进行遮挡,避免处理间光照影响。
1.4 测定方法
1.4.1 新梢及叶片生长指标的测定
在葡萄盛花期,每个处理选取10个长势均匀的新梢(总长度>1 m),用卷纸和游标卡尺测量新梢总长度、总节数、第3节新梢长度和粗度、第4张叶片宽度。计算平均节间长度=新梢总长度/总节数;叶面积大小=0.806 8x1.931 1[17]。将上述测量的叶片采回,置于105 ℃烘箱中杀青 30 min,75 ℃烘至恒质量后测定干物质含量(干质量/鲜质量×100%)。
1.4.2 叶绿素含量的测定
参考彭运生等的丙酮乙醇(2 ∶1)混合液测定法[18]。在葡萄果实转色期,每个小区选取5个长势均匀的新梢,分别选取新梢顶端第2张展开叶(嫩叶)和基部第3张叶(老叶),剪成宽度小于1 mm细丝,混合均匀,称取0.2 g叶片,放入丙酮 ∶无水乙醇=2 ∶1的混合液40 mL中,室温避光浸泡提取至叶片完全发白,采用分光光度计分别测定663 nm和645 nm处的吸光度,计算总叶绿素含量。
1.4.3 光合指标及光响应曲线的测定
葡萄盛花期,在天气晴朗的07:00—09:00,各处理选择新梢中部成熟叶片,用LI-6400便携式光合仪测定蒸腾速率、气孔导度、胞间CO2浓度。
5月20日,天气晴朗,于08:00—10:00测定新梢中部成熟叶片(第4~6张叶)的净光合速率,测定过程中使用光合仪的LED光源,光强Q值设置为2 000、1 800、1 600、1 400、1 200、1 000、800、600、500、400、300、200、100、50、0 μmol/(m2·s),CO2浓度设置为400 μmol/(m2·s),自动记录光合参数。
1.4.4 净光合速率测定
5月22日,天气晴朗,针对本研究的补光时段选取一天中06:00、08:00、10:00、12:00、14:00、16:00、18:00、20:00 6个时间点,用光合仪测定不同处理的净光合速率,仪器光强按自然光强进行设定,选取长势均匀的新梢中部成熟叶片(第5~6张叶),测定位置为叶片中部并避开叶片的主脉。
1.4.5 叶片养分的测定
在果实转色期,每个小区选取10个长势均匀的新梢,采取新梢中部正常生长的叶片,测定叶片中氮、磷、钾的含量,测定方法参考NY/T 2017—2011《植物中氮、磷、钾的测定》和NY/T 2420—2013《植株全钾含量测定 火焰光度计法》,采用凯氏法测定植物全氮含量,钼锑抗比色法测定植物全磷含量,火焰光度计法测定植物钾含量[19-20]。选取新梢顶端第2张展开叶(嫩叶)和基部第3张叶(老叶),分别测定老叶和嫩叶钙、镁的含量,测定方法参考GB 5009.268—2016《食品安全国家标准 食品中多元素的测定》,采用电感耦合等离子体质谱法测定钙、镁元素含量[21]。
1.4.6 果实品质测定
在果实完全成熟时采收,每个小区随机选取10串大小色泽均一的果穗进行果实品质的测定。每穗随机选取5粒果实,采用电子天平进行称质量,计算果实单粒质量;从中随机选取10粒用FHH-1型果实硬度计测定果肉硬度;用ATAGO PAL-1数显折射计测定可溶性固形物含量;用酸碱滴定法测定果实可滴定酸含量。
1.5 数据处理与分析
采用“叶子飘模型”对净光合速率随光照度响应曲线进行拟合分析得到光饱和点和光补偿点[22]。利用Excel 2016和SPSS 18.0软件对试验数据进行分析,采用Duncan s新复极差法进行显著性检验。
2 结果与分析
2.1 不同补光处理对葡萄新梢生长的影响
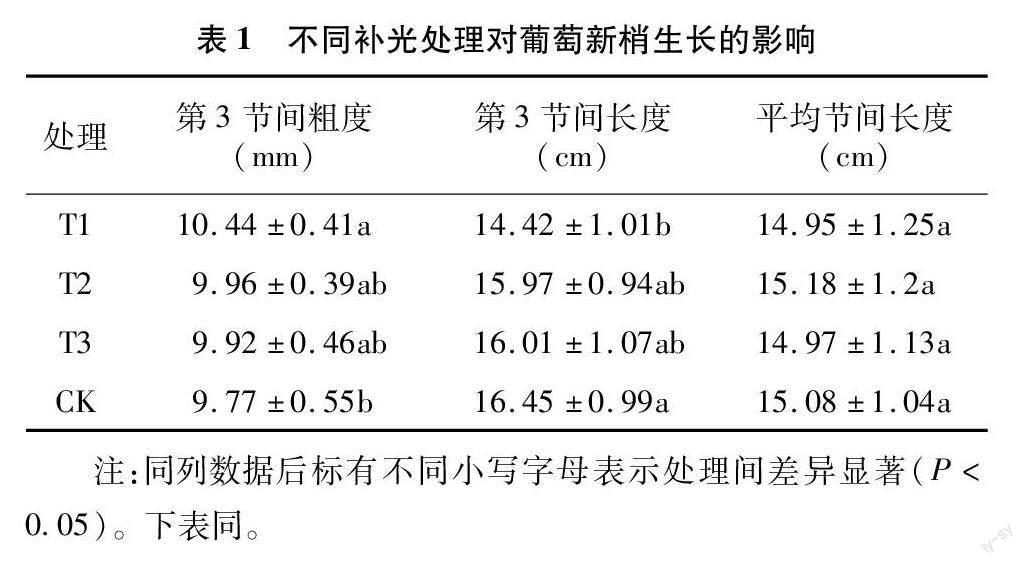
在初春葡萄设施栽培生长前期日照周期短,薄膜覆盖易造成棚内光照时长和光照度不足,新梢第3节间的长度和粗度反映了夏黑新梢初春季的生长发育状况。由表1可知,补光能增加新梢第3节间粗度,降低节间长度,但只有T1处理与对照差异显著。可见人工补光可以防止初春弱光环境下的新梢徒长,这与前人的研究结果[13,16]相一致。各补光处理新梢的平均节间长度与对照均无显著性差异,说明随着栽培后期温度上升,日照周期渐长,日照强度增强,已能满足葡萄生长的基本光强需求,补光不再影响新梢的节间长度。
2.2 不同补光处理对葡萄叶片生长的影响
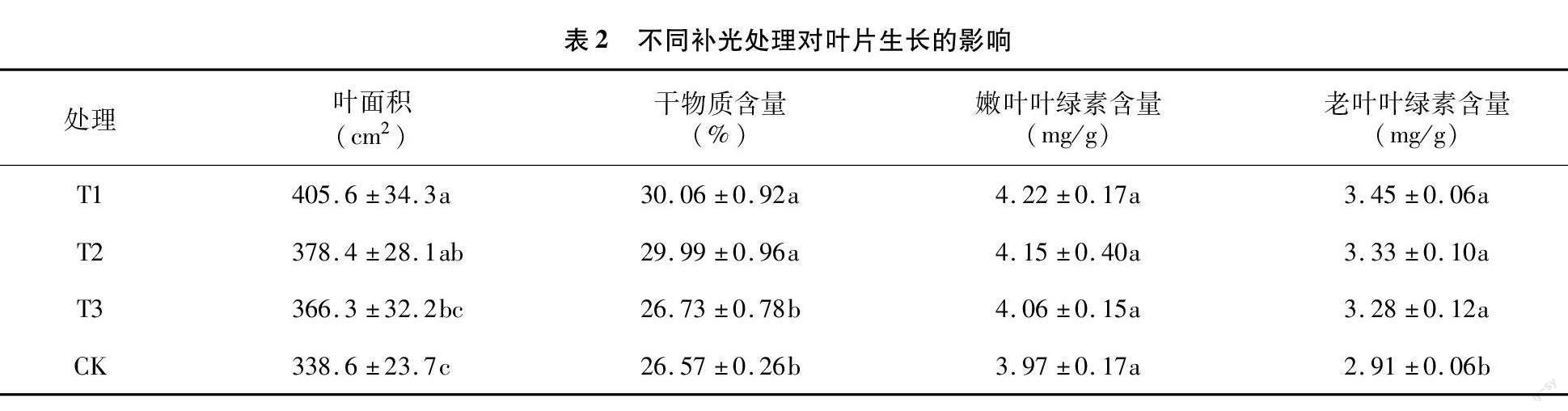
由表2可知,T1处理的叶面积、干物质含量和叶绿素含量最高,其次是T2和T3处理,对照最低。补光处理能增加叶片的面积和干物质含量,其中T1、T2处理与对照呈显著性差异,T3处理与对照差异不显著。补光处理嫩叶的叶绿素含量与对照相差不大,而补光处理的老叶的叶绿素含量都显著高于对照。与嫩叶相比,3个补光处理老叶的叶绿素含量分别比嫩叶低18.25%、19.76%、19.21%,而对照老叶叶绿素含量比嫩叶低达26.70%,可见补光延缓了老叶叶绿素的降解。
2.3 不同补光处理对叶片矿物质含量的影响
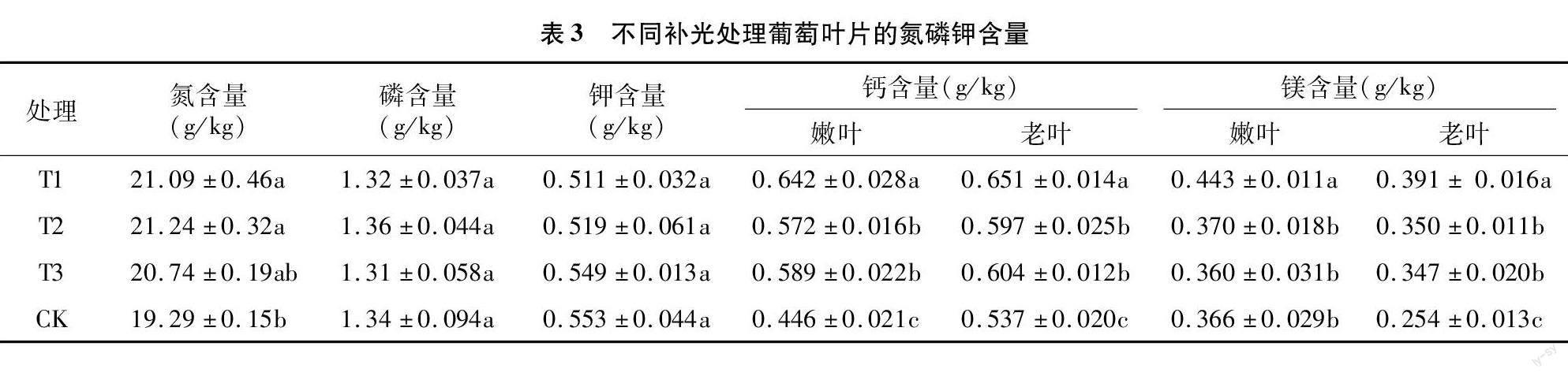
由表3可知,补光处理增加了葡萄叶片中氮的含量,T1、T2处理与对照呈显著性差异,分别增加了9.33%、10.11%。但补光对叶片中磷、钾含量无影响。
镁是植物体内可移动和重复利用的元素,缺乏时首先在老叶上体现;钙是不可移动再利用元素,缺乏时首先在嫩叶上体现。3个补光处理的嫩叶和老叶的钙含量都显著高于对照,其中T1处理的叶片钙含量最高,显著高于T2、T3处理。补光处理的嫩叶与老叶相比,钙含量相差不大,T1、T2、T3处理嫩叶的钙含量分别比老叶低1.38%、4.19%、2.48%,而对照嫩叶的钙含量比老叶低达16.95%。补光处理的老叶镁含量都显著高于对照,T1处理的嫩葉镁含量显著高于其他处理,T2、T3处理的嫩叶的镁含量与对照相差不大。3个补光处理老叶的镁含量比嫩叶的分别低11.74%、5.40%、3.61%,而对照老叶比嫩叶低达30.60%(表3)。
2.4 不同补光处理葡萄叶片的光合特征参数
光合作用是物质积累和能量转化的过程,气孔导度、胞间CO2浓度和蒸腾速率会间接影响叶片的净光合速率。由表4可知,胞间CO2浓度各处理间无显著性差异。补光处理T1、T2的最大净光合速率、光饱和点、气孔导度、蒸腾速率都显著高于CK,T3处理只有光饱和点显著高于对照。
2.5 不同补光处理的净光合速率日变化
棚内自然条件下的温度和光照度日变化如图1所示,光照度06:00开始逐渐升高,12:00—13:00达到最高点,然后逐步下降,19:00后棚内光照度基本为零。棚内温度06:00为16.9 ℃,后逐渐升高,13:00达到棚内最高温度33.5 ℃,11:00—15:00期间棚内温度都在30 ℃以上,下午棚内温度下降缓慢,17:00仍有28.3 ℃。
由图2可知,一天中净光合速率的日变化呈双峰型,上午逐渐升高,中午降低,出现“午休”现象,下午回升后又很快下降。早晨补光时段(05:00—09:00,T1、T2补光),各处理光合速率总体比较结果为:T1≈T2>T3>CK。在06:00时,3个补光处理的净光合速率相差不多,T1、T2显著高于对照。08:00 时,T1、T2、T3处理仍相差不大,但都显著高于对照。09:00—17:00时段(只有T1补光),各处理光合速率总体比较结果为:T1>T2>T3>CK。在 10:00 时,T1、T2处理的净光合速率相近,3个补光处理都显著高于对照。12:00—14:00温度和光照处于一天中的高峰,各处理都出现午休,T1、T2处理净光合速率相近,显著高于T3、CK。16:00,各处理光合速率回升,T1处理净光合速率显著高于其他处理,T1、T2处理显著高于对照。傍晚补光时段(17:00—21:00,T1、T3补光),各处理光合速率总体比较结果为:T1≈T3>T2>CK;18:00时,3个补光处理都显著高于对照。20:00时,自然光照度接近为0,各处理的净光合速率几乎为0,T3处理人工补光对光合作用也无效果。通过各处理净光合速率日变化比较,由于下午受亚高温影响,下午补光效果明显低于上午,T1处理的补光效果最好,光合速率全天都高于其他处理,早晨补光T2处理的效果优于傍晚补光T3处理,T2处理只有18:00的光合速率低于T3,其他时间光合速率比T3高或相近。
2.6 不同补光处理对葡萄果实品质的影响
由表5可知,补光处理促进了果实膨大,增加了果实单粒质量,T1、T2处理的单粒质量显著高于CK,T1处理的单粒质量比对照高9.09%。但在可溶性固形物含量、果肉硬度、可滴定酸含量方面,补光处理与对照无显著性差异。可见,补光处理在增加夏黑葡萄产量的同时保证了果实品质,并没有因为产量增加而降低果实品质。
3 讨论与结论
叶绿素是植物进行光合作用的主要色素,植物叶绿素含量与其净光合速率、连年丰产系数呈显著正相关关系,可作为评价植物生长状况、环境适应性等方面的基础数据[23-26]。随着叶片衰老,叶绿素会逐渐降解,叶绿素含量的下降是叶片衰老初期最明显的特征[27]。氮和镁是叶绿素分子的重要组成元素,本试验测定了不同处理叶片中叶绿素和矿物质元素含量,结果表明补光处理能提高叶片中叶绿素、氮、镁的含量,且随着叶片衰老及营养消耗,补光处理的老叶中叶绿素和镁降低量大大低于对照。可见,人工补光延缓了葡萄叶片的衰老,延长了叶片的功能期。王跃进等研究表明,叶片中的叶绿素含量与光照呈明显的正相关关系[28],也有很多研究证实补光有利于提高植物叶片的叶绿素含量[11,29-32]。王海波等研究表明,补光减缓了叶绿体超微结构的解体,有效延缓叶片衰老进程,延长了叶片的生理功能期[33]。本研究结果与以往研究报道相一致。
葡萄生产对钙的需求较高。目前,国内外关于植物钙吸收机制研究认为,钙在植物体内吸收和运输有被动吸收和主动吸收2种形式,主动吸收与根系活性有关,被动吸收主要受植物蒸腾作用的影响,钙由根系吸收后主要通过蒸腾液流由木质部运输到旺盛生长的枝梢、幼叶、花果等组织[34-35]。本试验中,补光处理显著增加了葡萄叶片的蒸腾速率,从而促进了钙的吸收和运输,使葡萄叶片中的钙含量显著高于对照。
本试验中夏黑葡萄光合速率日变化呈双峰形,具有明显光合“午休”现象。第1个高峰出现在 10:00 左右,第2个高峰出现在16:00左右,2个峰值远低于第1个峰值。这与陈冲等的测定结果[36-37]相一致。葡萄是对高温和强光胁迫比较敏感的果树,葡萄叶片光合作用最适温度分布在25~30 ℃[38],本试验设施大棚内最高光照度[<800 μmol/(m2·s)]远低于夏黑光合作用的光饱和点[>1 400 μmol/(m2·s)],而棚内温度 11:00—15:00间在30 ℃以上,下午至17:00一直处于28 ℃以上的亚高温区,可见高温是本试验中夏黑葡萄光合午休的主要原因。前人在大豆上的研究发现一定强度的光照能缓解高温胁迫对光合机构的伤害[39]。还有研究表明,钙能提高烟草、番茄、葡萄亚高温下叶片的净光合速率,维持气孔正常开闭,使光系统Ⅱ(PSⅡ)维持较高的活性[40-42]。本研究证实了这点,补光提高了夏黑葡萄叶片中钙的含量,缓解了高温对光合作用的影响,提高了夏黑葡萄中午时段的光合速率。
综上所述,人工补光措施改善了葡萄设施栽培棚内光照弱的问题,提高了葡萄叶片的叶绿素含量、净光合速率;提高了光合作用的气孔导度、蒸腾速率、光饱和点;增加了设施栽培葡萄新梢前期生长的粗度,缩短了节间长度,增大了叶片面积、干物质含量及矿物质元素含量,从而提高了树体营养积累,增强了树势。综合比较不同补光时段处理下葡萄生长发育、果实品质和光合特性指标,各处理的补光效果从高到低为:T1>T2>T3>CK。在实际生产环节中可综合考虑经济成本与补光效果,根据生产季节气候变化,合理安排补光时间段,以达到良好的经济效益与生态效益。
参考文献:
[1]段长青,刘崇怀,刘凤之,等. 新中国果树科学研究70年——葡萄[J]. 果樹学报,2019,36(10):1292-1301.
[2]王世平,李 勃. 中国设施葡萄发展概况[J]. 落叶果树,2019,51(1):1-5.
[3]Meng J F,Ning P F,Xu T F,et al. Effect of rain-shelter cultivation of Vitis vinifera cv.Cabernet Gernischet on the phenolic profile of berry skins and the incidence of grape diseases[J]. Molecules,2012,18(1):381-397.
[4]曹 锰. 避雨栽培对葡萄生长小气候及果实品质的影响[D]. 北京:中国农业科学院,2015:1.
[5]李洪艳,文仁德,张 兰,等. 补光处理对设施栽培巨峰葡萄冬季果生长的影响[J]. 中国南方果树,2016,45(5):93-94,100.
[6]张克坤,刘凤之,王孝娣,等. 不同光质补光对促早栽培‘瑞都香玉葡萄果实品质的影响[J]. 应用生态学报,2017,28(1):115-126.
[7]王佳淇,韦晓桐,何莹钰,等. LED补光系统对设施园艺作物的影响[J]. 浙江农业科学,2020,61(5):950-954.
[8]王海波,王孝娣,赵君全,等. 设施促早栽培下耐弱光能力不同的葡萄品种冬芽的花芽分化[J]. 园艺学报,2016,43(4):633-642.
[9]赵光强. 弱光对葡萄生长发育影响机理的研究[J]. 潍坊学院学报,2006,6(6):99-101.
[10]王佳淇. 不同光质LED补光对大棚蓝莓生长发育的影响研究[D]. 金华:浙江师范大学,2020:2-5.
[11]季亚萍. 不同光质LED组合光源对石榴幼树生长和果实品质的影响[D]. 郑州:河南农业大学,2017.
[12]段青青,张禄祺,张自坤,等. 补光时间及光质对温室甜椒生长及产量品质的影响[J]. 农业工程学报,2019,35(24):213-222.
[13]黄秋凤,谢蜀豫,曹慕明,等. 夜间补光对巨峰葡萄春果叶片营养及果实品质的影响[J]. 南方农业学报,2019,50(4):781-787.
[14]成 果,陈国品,郭荣荣,等. 补光处理对巨峰葡萄春果花色苷组分的影响[J]. 南方园艺,2017,28(5):1-8.
[15]時晓芳,曹雄军,林 玲,等. 阳光玫瑰葡萄冬果膨大期补光对内源激素和果实品质的影响[J]. 中国南方果树,2021,50(2):122-127.
[16]孔 云,王绍辉,沈红香,等. 不同光质补光对温室葡萄新梢生长的影响[J]. 北京农学院学报,2006,21(3):23-25.
[17]项殿芳,吴学仁,赵德生. 葡萄叶面积测定方法的改进[J]. 河北职业技术师范学院学报,2002,16(4):1-5,11.
[18]彭运生,刘 恩. 关于提取叶绿素方法的比较研究[J]. 北京农业大学学报,1992(3):247-250.
[19]中华人民共和国农业部. 植物中氮、磷、钾的测定:NY/T 2017—2011[S]. 北京:中国农业出版社,2011.
[20]中华人民共和国农业部. 植株全钾含量测定 火焰光度计法:NY/T 2420—2013[S]. 北京:中国农业出版社,2014.
[21]国家卫生和计划生育委员会,国家食品药品监督管理总局. 食品安全国家标准 食品中多元素的测定:GB 5009.268—2016[S]. 北京:中国标准出版社,2017.
[22]Ye Z P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica,2007,45(4):637-640.
[23]王玉莲. 葡萄早期新生叶片叶绿素含量变化检测[J]. 河北林业科技,2006(3):4-5,8.
[24]徐成东,张 薇. 五个葡萄品种叶色素含量与光合特性分析[J]. 楚雄师范学院学报,2020,35(3):47-53.
[25]谢计蒙,王海波,王孝娣,等. 设施葡萄品种连年丰产能力与光合生理特性关系研究[J]. 果树学报,2012,29(5):843-851.
[26]王海波,王孝娣,韩 晓,等. 不同葡萄品种设施环境适应性评价方法研究[J]. 华北农学报,2019,34(增刊1):92-98.
[27]Zimmermann P,Zentgraf U. The correlation between oxidative stress and leaf senescence during plant development[J]. Cellular & Molecular Biology Letters,2005,10(3):515-534.
[28]王跃进,傅耕夫,杨晓盆. 光对苹果叶片及其叶绿体结构的影响[J]. 果树科学,1988,5(4):145-149.
[29]闻 靖,杨其长,魏灵玲,等. 不同红蓝LED组合光源对叶用莴苣光合特性和品质的影响及节能评价[J]. 园艺学报,2011,38(4):761-769.
[30]Liu N,Ji F,Xu L J,et al. Effects of LED light quality on the growth of pepper seedling in plant factory[J]. International Journal of Agricultural and Biological Engineering,2019,12(5):44-50.
[31]黄秋凤,陈 立,李 敏,等. 夜间延时补光调控对巨峰葡萄春果生长发育及光合特性的影响[J]. 南方农业学报,2021,52(8):2227-2233.
[32]孟力力,宋江峰,柏宗春,等. 远红光对生菜光合作用及叶绿素荧光特性的影响[J]. 江苏农业学报,2022,38(1):181-189.
[33]王海波,王 帅,王孝娣,等. 葡萄叶片衰老过程中不同光质对其光合和叶绿体超微结构的影响[J]. 园艺学报,2019,46(2):205-214.
[34]罗 媛. 菹草根/叶对钙的吸收/释放机制研究[D]. 武汉:华中农业大学,2013:6-7.
[35]马建军. 野生欧李钙素分配特性的研究[D]. 北京:中国农业大学,2005:1-2.
[36]陈 冲. 避雨栽培葡萄光合特性研究[D]. 长沙:湖南农业大学,2011.
[37]娄玉穗,尚泓泉,吕中伟,等. 基于光合特性的8个葡萄品种耐弱光、抗高温特点比较[J]. 果树学报,2021,38(9):1491-1502.
[38]唐桓伟. 一年两收栽培阳光玫瑰葡萄夏果和冬果生长期叶片光合特性研究[D]. 南宁:广西大学,2019.
[39]Havaux M,Greppin H,Strasser R J. Functioning of photosystems Ⅰ and Ⅱ in pea leaves exposed to heat stress in the presence or absence of light[J]. Planta,1991,186(1):88-98.
[40]谭 伟,李庆亮,罗 音,等. 外源CaCl2预处理对高温胁迫烟草叶片光合作用的影响[J]. 中国农业科学,2009,42(11):3871-3879.
[41]齐明芳,刘玉凤,周龙发,等. 钙对亚高温下番茄幼苗叶片光合作用的调控作用[J]. 中国农业科学,2011,44(3):531-537.
[42]郑秋玲,谭 伟,马 宁,等. 钙对高温下巨峰葡萄叶片光合作用和叶绿素荧光的影响[J]. 中国农业科学,2010,43(9):1963-1968.
猜你喜欢
小读者(2021年2期)2021-03-29
电子制作(2019年19期)2019-11-23
华人时刊(2019年13期)2019-11-17
今日农业(2019年16期)2019-01-03
中国交通信息化(2017年8期)2017-06-06
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
现代农业(2016年6期)2016-02-28
