
天目山针阔混交林与常绿阔叶林的空间结构比较*
2023-08-09 03:01汤孟平
林业科学 2023年5期
陈 睿 汤孟平
(1. 浙江农林大学省部共建亚热带森林培育国家重点实验室 临安 311300;2. 浙江农林大学环境与资源学院 临安 311300)
森林结构通常指不同种类和大小的植物的空间配置和分布状况,它决定着森林的功能和稳定健康程度。森林结构可分为空间结构和非空间结构两大类。森林空间结构是基于树木位置的结构,如树木的分布、树种相互隔离程度等;而非空间结构是指树种组成、直径分布、年龄结构等。森林经营的目标已由森林永续利用转变为森林可持续经营。Baskent 等(2005)指出,不考虑空间细节特征, 要维持环境和生态条件如生物多样性的稳定是不可能的。森林空间结构影响树木之间的竞争格局、森林群落的稳定性和森林经营措施发展方向(汤孟平等,2013;胡艳波等,2006)。因此,为实现森林可持续经营,获取精确的森林空间结构信息是十分重要的(汤孟平,2010;Szmytet al., 2018)。
目前,对森林空间结构的研究已成为了国际林业研究热点之一(赵中华等,2009;田国恒等,2010;邵芳丽等,2011;De Grooteet al., 2018)。常绿阔叶林和针阔混交林是亚热带的两种主要森林类型,其中常绿阔叶林是亚热带的顶极群落。常绿阔叶林的演替过程十分漫长,从次生裸地→灌丛→针叶林→针阔混交林→常绿阔叶林(宋永昌等,1995),整个演替需时约150 年以上(宋永昌,2013),每个阶段需20~30 年(余树全,2003)。在常绿阔叶林演替过程中,针阔混交林是演替为常绿阔叶林的关键阶段。从针阔混交林阶段开始,群落结构复杂性明显增加。近年来,学者们对亚热带常绿阔叶林和针阔混交林的空间结构开展了大量研究。吴大荣等(2003)在福建省三明市罗卜岩自然保护区内设置样地,应用相邻格子法进行每木调查,研究了常绿阔叶林的闽楠(Phoebe bournei)种群结构和空间分布格局;汤孟平等(2009)提出了基于Voronoi 图的混交度,并用于天目山常绿阔叶林混交度研究;汤孟平等(2012)还提出了全混交度,并研究比较了天目山常绿阔叶林的几种混交度,表明全混交度对不同混交结构均有较高分辨能力;赵洋毅等(2012)研究了中国西南亚热带地区典型常绿阔叶林的空间结构,发现2 种典型天然常绿阔叶林的主要建群树种均处于聚集状态,且轻度聚集状态的栲树(Castanopsis fargesii)林更稳定;曹小玉等(2020)在人工针阔混交林等典型森林群落研究了森林群落空间结构对土壤养分含量的影响,发现林分空间结构与土壤养分整体上相关极显著,混交度和林层指数对土壤养分的整体影响力起着决定作用。可见,以上学者侧重于这2 种森林类型中某一森林类型的空间结构特征分析。事实上,针阔混交林是向顶极群落常绿阔叶林演替的最后阶段的过渡森林类型,对这两种森林类型的空间结构进行比较,有助于从空间结构角度揭示该地区的森林演替规律。然而,这方面研究尚少见报道。
本研究采取空间代时间方法(刘保双等,2013),在地处亚热带的浙江天目山国家级自然保护区,选择针阔混交林和常绿阔叶林,分别设置大型固定标准地,进行每木精确定位,调查获取数据,采用聚集指数、竞争指数和混交度指数,对两类森林的空间结构特征进行比较,旨在从空间结构角度揭示群落演替规律,为常绿阔叶林与针阔混交林可持续经营提供依据。
1 研究区概况
浙江天目山国家级自然保护区地处临安区西北部(119°24′11″—119°28 ′21″ E, 30°18′30″—30°24′55″ N),海拔300~1 556 m,总面积4 284 hm2。具有典型的中亚热带森林生态系统和森林景观。该区土壤随海拔升高由红壤向棕黄壤过渡,近山脚局部地带还分布有石灰性土壤,海拔600 m 以下为红壤,海拔600~1 200 m为黄壤,海拔1 200 m 以上为棕黄壤。植被类型丰富,植被分布有明显垂直变化:常绿阔叶林为典型地带性植被,主要分布在海拔850 m 以下;常绿落叶阔叶混交林为主要植被类型,主要分布在海拔850~1 100 m;落叶阔叶林分布在海拔1 100~1 380 m;落叶矮林分布在海拔1 380 m 以上(周红敏等,2009)。其中,该地区常绿阔叶林乔木树种以壳斗科(Fagaceae)常绿树种为主,如青冈(Cyclobalanopsis glauca)、细叶青冈(C.gracilis)、短尾柯(Lithocarpus brevicaudatus)等,此外还常混生有常绿针叶和落叶阔叶树种,如杉木(Cunninghamia lanceolata)、榧树(Torreya grandis)、枫香(Liquidambar formosana)、榉树(Zelkova serrata)等(汤孟平等,2006)。
2 研究方法
2.1 样地调查
常绿阔叶林和针阔混交林是浙江天目山国家级自然保护区的两种典型森林类型。原有天然常绿阔叶林在1941 年受山火干扰,于1956 年建立浙江天目山国家级自然保护区后自然缓慢恢复。研究区的针阔混交林则起源于1953 年建立天目山林场时人工种植的杉木林,对原有森林植被自然恢复进程有一定影响,1988 年限制采伐后,由人工经营转为近自然经营。
为监测浙江天目山自然保护区内的常绿阔叶林和针阔混交林的结构动态变化,于2005、2006 年先后在常绿阔叶林和针阔混交林中分别建立了1 块1 hm2(100 m × 100 m)固定样地。常绿阔叶林固定样地的立地特征为坡向南坡,坡位中坡,坡度38°,海拔645 m。针阔混交林固定样地的立地特征为坡向南偏西36°,坡位中坡,坡度33°,海拔640 m。采用相邻格子调查法,把样地划分为100 个10 m × 10 m 的调查单元。在每个调查单元内,对胸径 ≥ 5 cm 的树木挂牌编号,记录树种名称,测量胸径、树高、冠幅等因子。采用激光对中全站仪(徕卡TCR702Xrange)测定每株树木基部的三维坐标 (x,y,z) (汤孟平等,2006)。分别于2015、2016 年进行复查。本研究采用2015、2016 年的复查数据。
2.2 空间结构指数
1)混交度 混交度计算采用基于Voronoi 图的全混交度指数(汤孟平等,2012),计算公式为:
式中:Mci为对象木i的全混交度;ni为对象木i的最近邻木株数;Mi为对象木i的简单混交度,计算公式为:是一个离散性的变量,当对象木i与第j株相邻木非同种时,Vij= 1,反之Vij= 0;ci为对象木的最近邻木中成对相邻木中非同种的个数;表示最近邻木的隔离程度;Di为空间结构单元的Simpso n指数,表示树种分布的均匀度,[0,1],当只有1 个树种时,Di= 0,当有无限多个树种且株数比例均等时,Di= 1,Pj是空间结构单元中第j树种的株数比例,Si为空间结构单元的树种数。
按(1)式计算的混交度是以对象木为中心的局部混交度,在对森林群落的研究中还要计算平均混交度:
关于(1)式中n的取值,本研究采用基于Voronoi图相邻多边形个数来确定相邻木个数。混交度的范围是[0, 1],将混交度划分为5 个等级,即0 为零度混交,(0, 0.25]为低度混交,(0.25, 0.5]为中度混交,(0.5,0.75]为强度混交,(0.75, 1]为极强混交(李际平等,2014)。
2)聚集指数 采用聚集指数分析林木空间分布格局。聚集指数是相邻最近单株距离的平均值与随机分布下期望的平均距离之比(Clarket al., 1954),计算公式为:
森林群落聚集指数的公式为:
式(3)和(4)中:Ri为对象木i的聚集指数;R为聚集指数;ri为第i株单株到它最近相邻木的距离;F为样地面积。
根据(4)式计算结果,如果R> 1、R< 1、R= 1,则该样地内林木有均匀分布、聚集分布、随机分布的趋势。在聚集分布(0 ≤R< 1)情况下,根据R的取值范围划分聚集程度,取值在[0, 0.25)、[0.25, 0.5)、[0.5,0.75)、[0.75, 1)区间内时,分别表示强度、中度、低度、弱度的聚集等级。取值在(1, 1.38)、[1.38, 1.76)、[1.76,2.14)、[2.14, ∞)区间内时,分别表示为弱均匀、低均匀、中均匀、强均匀4 个等级。
3)竞争指数 通常采用竞争指数描述树木之间对环境资源的竞争关系。竞争指数的选择除了要保证理论上的合理性,还要考虑到实际操作的可行性(汤孟平等,2013)。在多种竞争指数中,Hegyi 竞争指数的测定和计算均简单,所以最常用,而基于Voronoi图的Hegyi 竞争指数,改进了Hegyi 竞争指数在确定竞争单元时尺度不统一的不足(汤孟平等,2007)。因此,本研究中采用基于Voronoi 图的Hegyi 竞争指数(V_Hegyi 竞争指数),计算公式为(汤孟平等,2007):
式中:CIi为对象木i的竞争指数;Lij为对象木i与竞争木j之间的距离;di为对象木i的胸径;dj为竞争木j的胸径。
样地中所有对象木的竞争指数为:
式中:CI 为森林群落竞争指数;CIi为对象木i的竞争指数。
森林群落竞争指数为:
当对象木和竞争木为同一树种时,CI 表示种内竞争指数,否则为种间竞争指数。本研究根据森林CI实际分布,确定CI 取值在 [0, 12)、[12, 28)、[28, 44)、[44, ∞)区间时,分别表示弱度、低度、中度、强度4 个竞争等级。
2.3 q 值
异龄林株数按径级的分布可用负指数分布表示(于政中,1993;Garciaet al., 1999),公式为:
式中:D为胸径;a,k为常数(由曲线拟合后得到)。
根据(8)式,异龄林相邻径级株数之比为:
式中:q为2 个相邻径级中较小径级株数与较大径级株数之比;h为径阶距。
通常认为,q值在1.3~1.7 之间较为合理(Garciaet al.,1999)。
2.4 边缘矫正
当对象木处于样地边缘时,以其为中心所构建的林木空间结构单元会不完整,从而导致计算结果有误,为了消除这个边缘效应,需进行边缘矫正(周红敏等,2009)。一般采用8 邻域法进行边缘矫正,即仅以原样地林木作为对象木进行空间结构指数的计算(汤孟平等,2009)。但是,不同地形对空间结构特征有着不同程度的影响(龙俊松等,2021)。若采用传统的平移整体样地的8 邻域法,当样地跨度过大时,矫正邻域地形与实际邻域地形有显著差异。为此,本研究改进传统的8 邻域法,采用平移边缘单元格的方法进行边缘矫正。
2.5 优势树种确定
优势树种对整个森林群落和环境的形成与变化有显著控制作用(汤孟平等,2006)。本研究采用Ohsawa 优势度分析法确定群落优势树种(Ohsawa,1984),计算公式为:
式中:Y为总树种数;T为前位树种数;U为剩利树种数;Xi为第i个前位树种的胸高断面积百分比,X为优势树种理想胸高断面积百分比,Xj为第i个剩余树种(U)的胸高断面积百分比。假设群落只有1 个优势树种,则优势树种的理想百分比为100%;如果有2 个优势树种,则理想百分比为50%。以此类推,分别计算每种情况下的d值。当d值计算为最小值时,前位树种的个数即是群落优势种数。
2.6 统计分析
首先,应用Python 编程语言,结合ArcMap 10.7 软件的Voronoi 图生成功能及面邻域分析和近邻分析功能,编程计算空间结构指数。然后,采用IBM SPSS 20 中的Kolmogorov-Smirnov 检验法(以下简称K-S 检验),检验数据是否服从正态分布。若服从正态分布,则用t 检验分析森林群落的空间结构指数之间的差异;若不服从正态分布,则用Wilcoxon-Mann-Whitney 秩和检验法(以下简称W-M-W 检验)分析不同森林群落的空间结构指数之间的差异。
3 结果与分析
3.1 群落空间结构比较
3.1.1 2 种森林群落结构基本概况 2 种森林群落的株数密度、树种数、q值、空间结构指数统计结果见表1。空间结构指数计算只针对胸径 ≥ 5 cm 的树木。可以看出,常绿阔叶林与针阔混交林都是典型的异龄混交林,常绿阔叶林的株数密度比针阔混交林大54.72%,树种多23 种。2 种森林类型的株数与胸径的关系见图1,2 种森林株数随径级呈倒J 型分布,其中常绿阔叶林的株数分布与对象木径级相关性更高,R2=0.967 6,q值比针阔混交林小7.3%,株数按直径分布更趋平缓。常绿阔叶林的q值为1.608,位于1.3~1.7范围内;针阔混交林q值为1.735,大于1.7,因此常绿阔叶林株数按直径分布结构为合理状态,针阔混交林株数按直径分布结构则不合理。在空间结构方面,常绿阔叶林群落聚集指数为0.943,林木分布格局属弱度聚集;针阔混交林群落聚集指数为1.004,林木分布格局为弱均匀分布。2 种森林群落的全混交度均在[0.5,0.75)之间,混交程度为中度混交。森林群落V_Hegyi竞争指数在常绿阔叶林处于[28, 44)范围,属中度竞争;针阔混交林处于[12, 28)范围内,整体呈低度竞争。
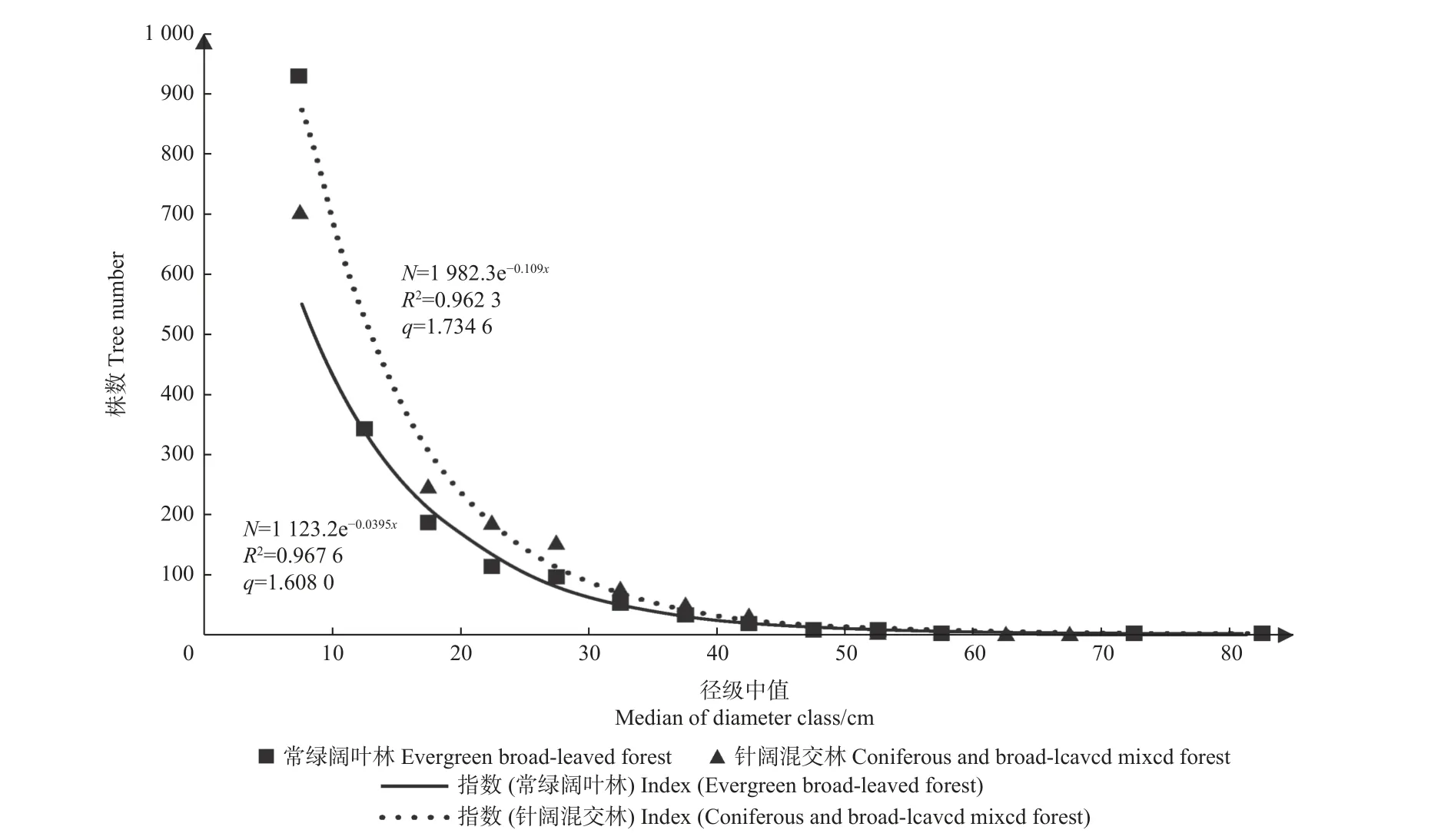
图1 两种森林群落株数与径级的关系Fig. 1 Relationship between tree number and diameter class for two forest communities
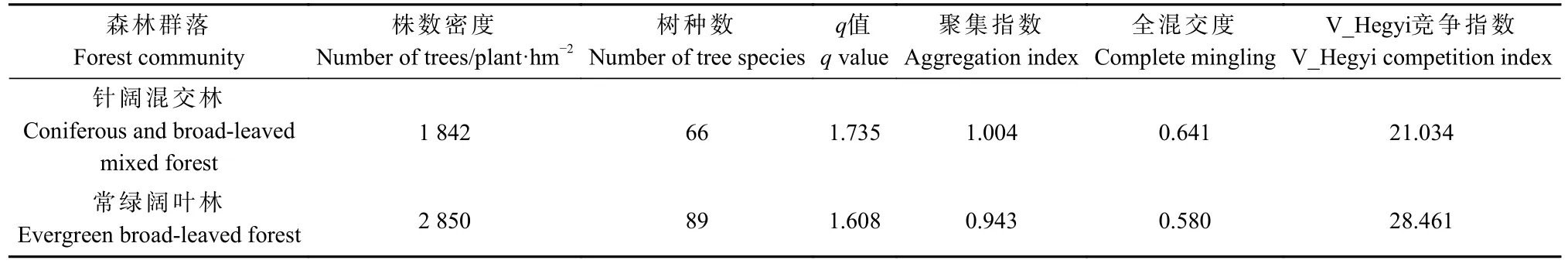
表1 群落因子统计结果Tab. 1 Statistical results of community factors
3.1.2 森林群落林木空间分布格局比较 从表1 可看出,常绿阔叶林群落的林木分布格局属弱度聚集,针阔混交林的林木分布格局属弱均匀分布,K-S 检验表明2 个群落的聚集指数均不符合正态分布(P< 0.05)。因此,采用W-M-W 检验,表明存在显著差异(P<0.01)。常绿阔叶林在演替周期上属针阔混交林演替的最高阶段,2 个森林群落演替阶段的不同表现在分布格局上存在显著差异。
对2 种森林群落的对象木胸径按5 cm 间距划分径级,并统计胸径 ≥ 5 cm 树木的株数密度,常绿阔叶林为1 795 株·hm-2,针阔混交林为1 822 株·hm-2(表2)。群落整体中,在常绿阔叶林和针阔混交林中,呈现聚集分布的林木(R< 1)分别占65.13%和60.65%,最小径级占比例分别高达51.75%、38.69%。在常绿阔叶林中,随着林木径级增长,聚集指数呈现较明显上升趋势,即林木分布趋于均匀;针阔混交林中则相对不明显,在大径级中存在随径级增长聚集指数下降的情况。
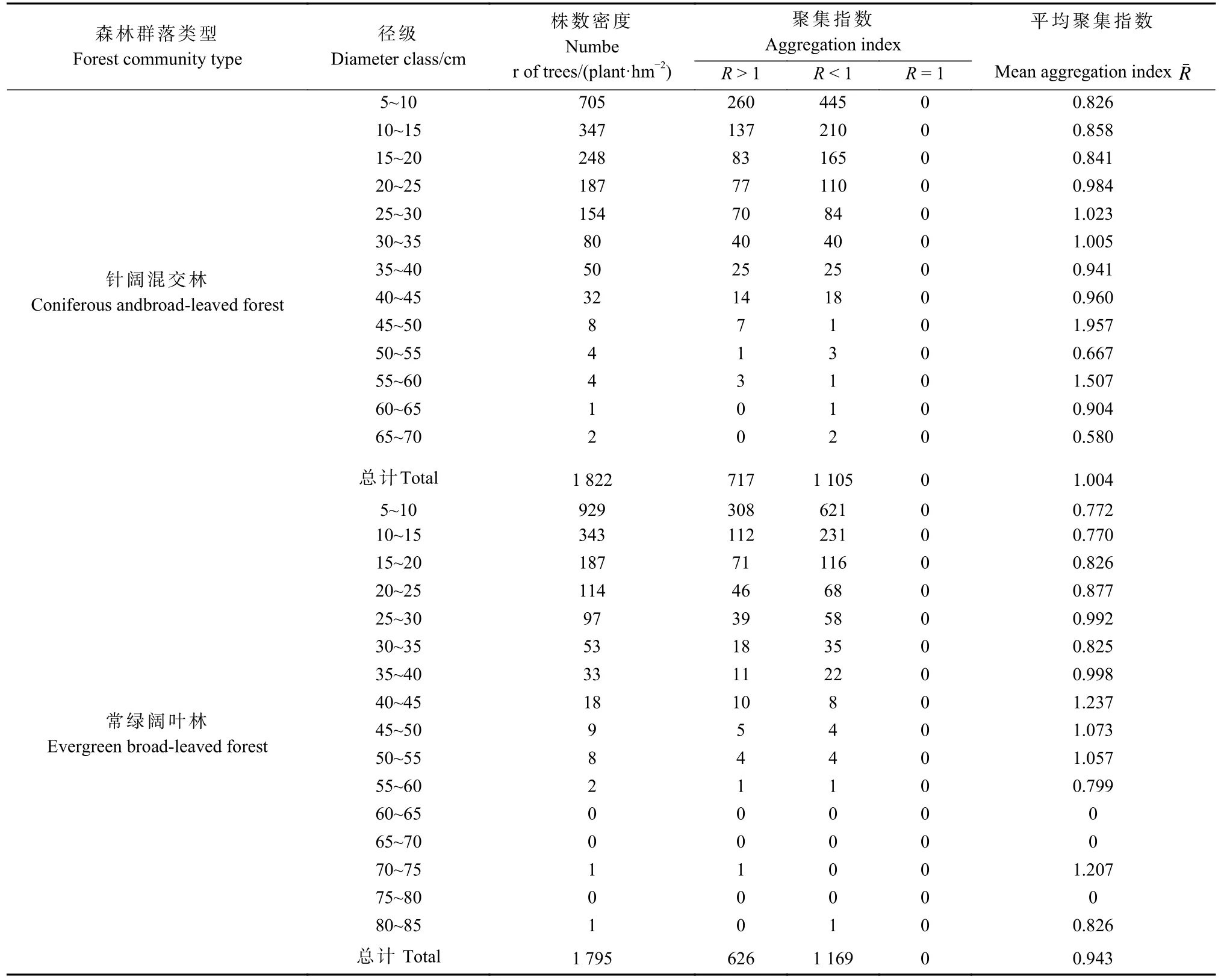
表2 对象木径级与聚集指数的关系Tab. 2 The relationship between Diameter class of subject trees and aggregation index
3.1.3 森林群落混交度比较 树种的混交程度是由树种的数量和相互隔离关系决定的。一般情况下,混交度越高,群落越稳定(汤孟平等,2004;邢晖,2014)。常绿阔叶林和针阔混交林的森林群落树种混交情况见图2,2 个群落中混交度等级占比最大的为强度混交,分别占了45.32%和47.81%,且混交度等级按单木数量从高到低排序均为强度混交、极强混交、中度混交、低度混交和零度混交。从图2 可看出,针阔混交林的单木强度混交和极强混交比例比常绿阔叶林高。K-S 检验结果显示,2 个群落混交度分布均不属于正态分布(P< 0.05)。因此,采用W-M-W 检验进行差异性分析,结果显示2 个群落的全混交度之间存在显著差异(P< 0.01)。
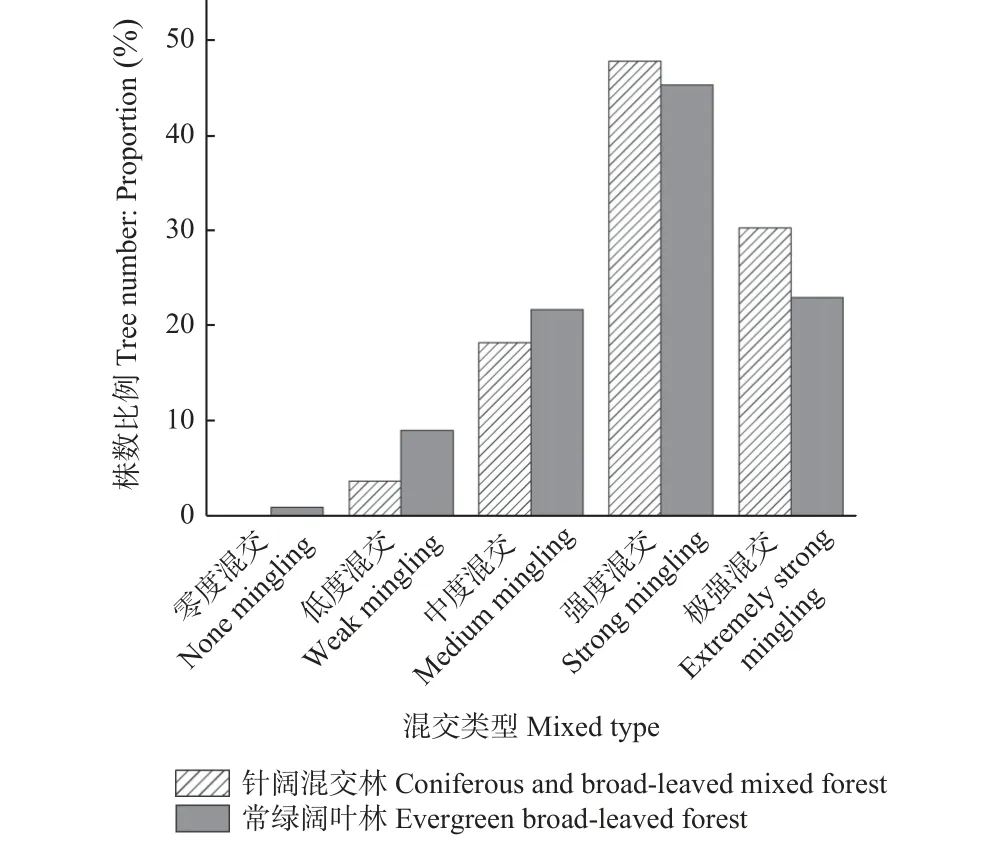
图2 两种森林群落全混交度分布Fig. 2 Frequency distribution histogram of mingling for two forest communities
3.1.4 森林群落竞争强度比较 树木竞争指数能反映树木与其相邻木的竞争关系。竞争指数越低表示受到竞争压力越小,即该树种竞争生长优势越大。常绿阔叶林和针阔混交林的群落竞争情况见图3,2 个森林群落中,均以弱度和低度竞争占主导地位,其中弱度竞争占最大比重,常绿阔叶林为50.67%,其余依次是低度竞争25.81%、强度竞争13.99%和中度竞争9.53%;在针阔混交林中,弱度竞争占比更高,为61.26%,其次为低度竞争23.59%、强度竞争8.50%和中度竞争6.66%。可以看出常绿阔叶林中弱度竞争林木数量比例小于针阔混交林。在常绿阔叶林中,除弱度竞争以外的竞争强度的林木数量占比均高于针阔混交林。对两个森林群落的V_Hegyi竞争指数进行KS 检验,结果显示均不符合正态分布(P< 0.05);因此进行了W-M-W 检验,结果显示2 个森林群落的V_Hegyi 竞争指数存在显著差异(P< 0.01)。
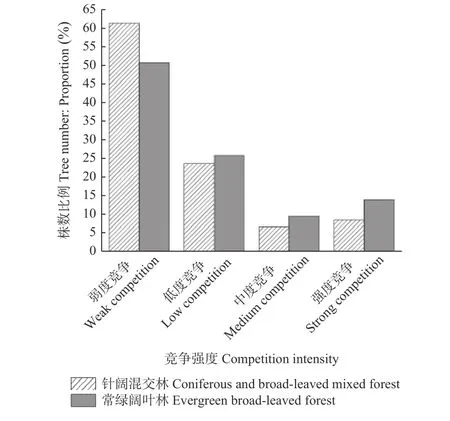
图3 2 种森林群落竞争强度分布Fig. 3 Frequency distribution histogram of V_Hegyi competition index for two forest communities
3.2 森林群落优势树种空间结构比较
3.2.1 2 种森林群落优势树种基本概况 常绿阔叶林和针阔混交林的优势树种分别为14 和4 种(表3),均包含常绿阔叶树种、常绿针叶树种和落叶阔叶树种。相对而言,常绿阔叶林各优势树种胸高断面积比例较针阔混交林均匀。针阔混交林中杉木占49.3%,处于明显优势地位,高于常绿阔叶林前5 个优势树种的胸高断面积比例之和(44.44%)。
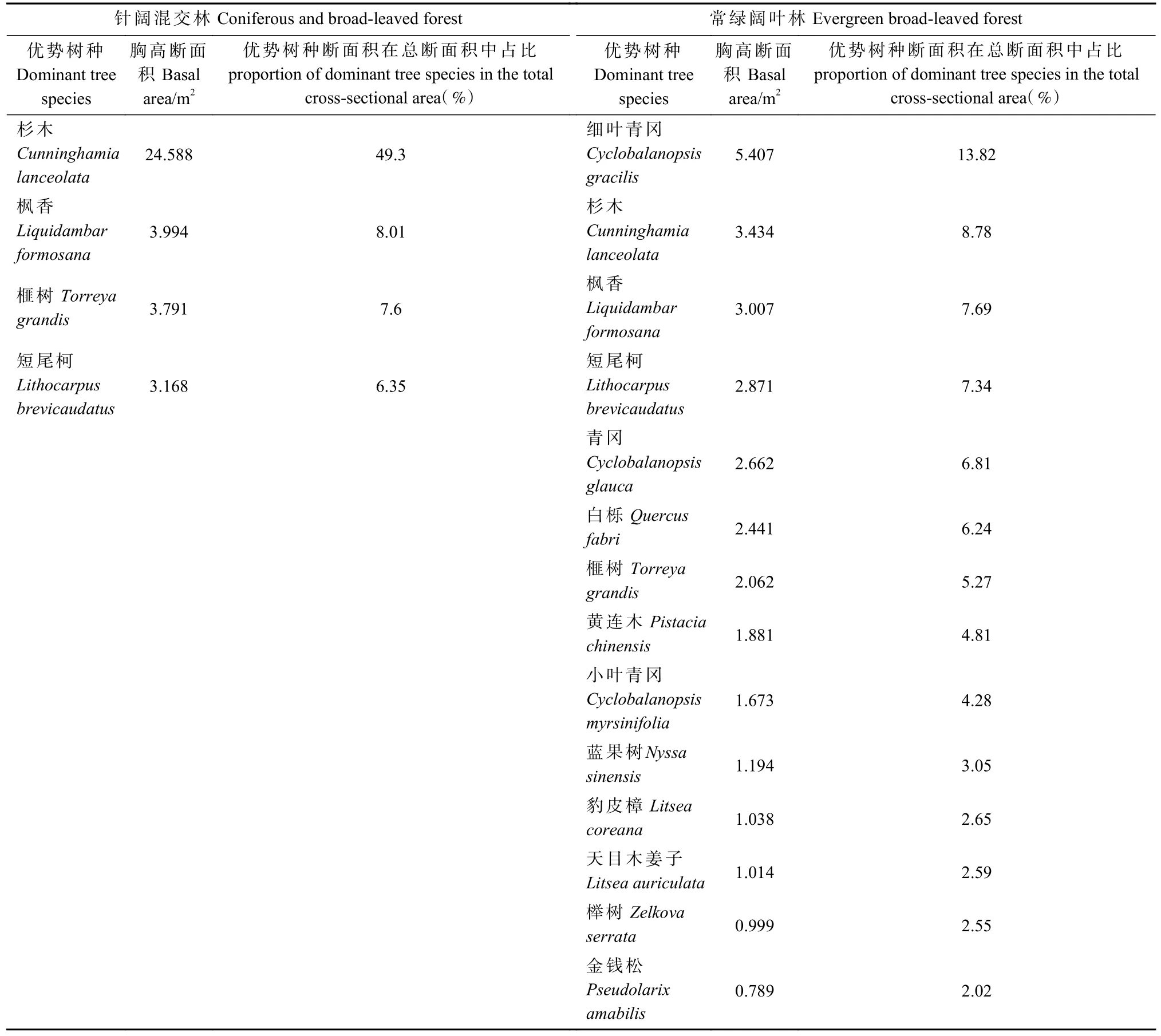
表3 各群落优势种组成情况Tab. 3 Composition of dominant tree species in community
3.2.2 2 种森林群落优势树种分布格局比较 2 个森林群落优势树种的聚集指数见图4。在常绿阔叶林中,常绿阔叶树种均处于弱度聚集状态,与森林群落整体分布格局相似,而榧树(常绿针叶树种)、金钱松(常绿针叶种)和蓝果树(落叶阔叶种)处于均匀分布状态。针阔混交林中,枫香和榧树处于均匀分布状态,杉木和短尾柯处于弱度聚集状态。与常绿阔叶林相比,针阔混交林优势树种的聚集指数变化波动较小,即在针阔混交林中各优势树种空间分布格局更为相似。
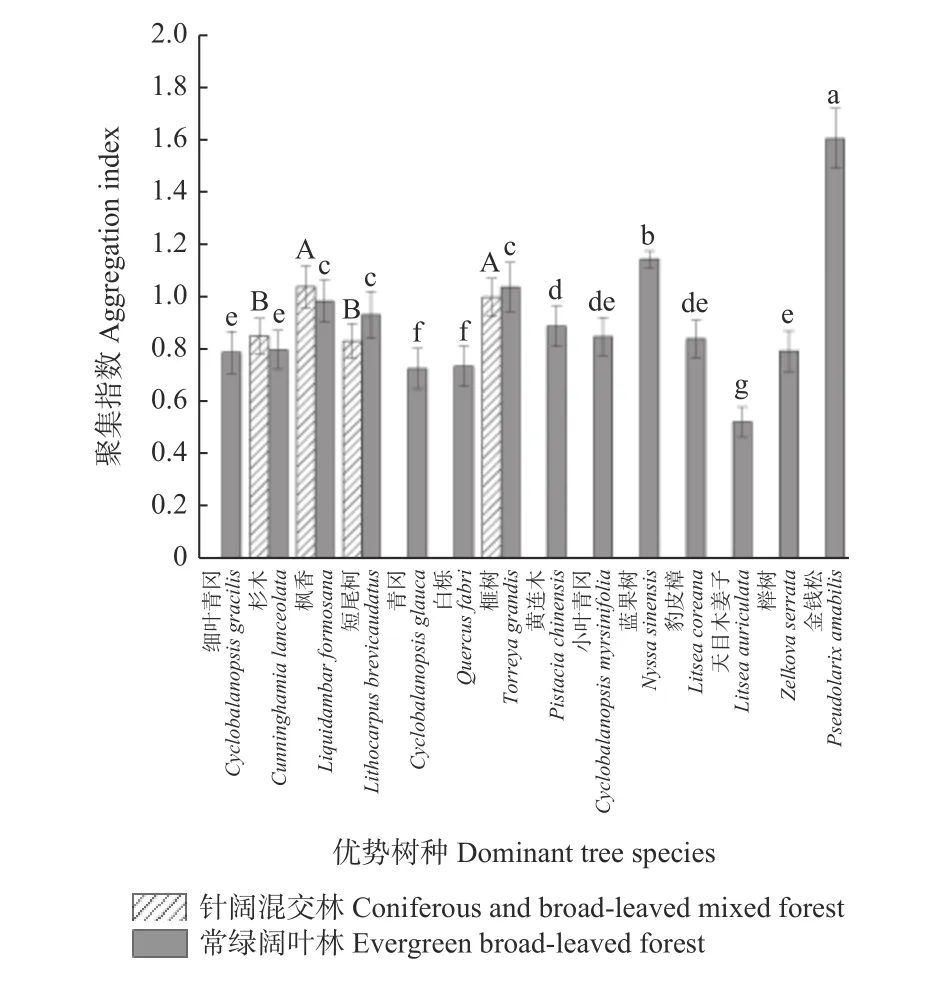
图4 2 种森林群落优势树种聚集指数比较Fig. 4 Aggregation index of dominant tree species in two forest communities
3.2.3 2 种森林群落优势树种混交度比较 2 种森林群落优势树种的全混交度见图5。常绿阔叶林中各优势树种的全混交度之间存在显著差异。细叶青冈虽然处于最高优势地位,但是树种全混交度最低。金钱松在优势树种中优势地位最低,全混交度最高。在针阔混交林中,4 个优势树种的全混交度则较为平均,无明显差异。
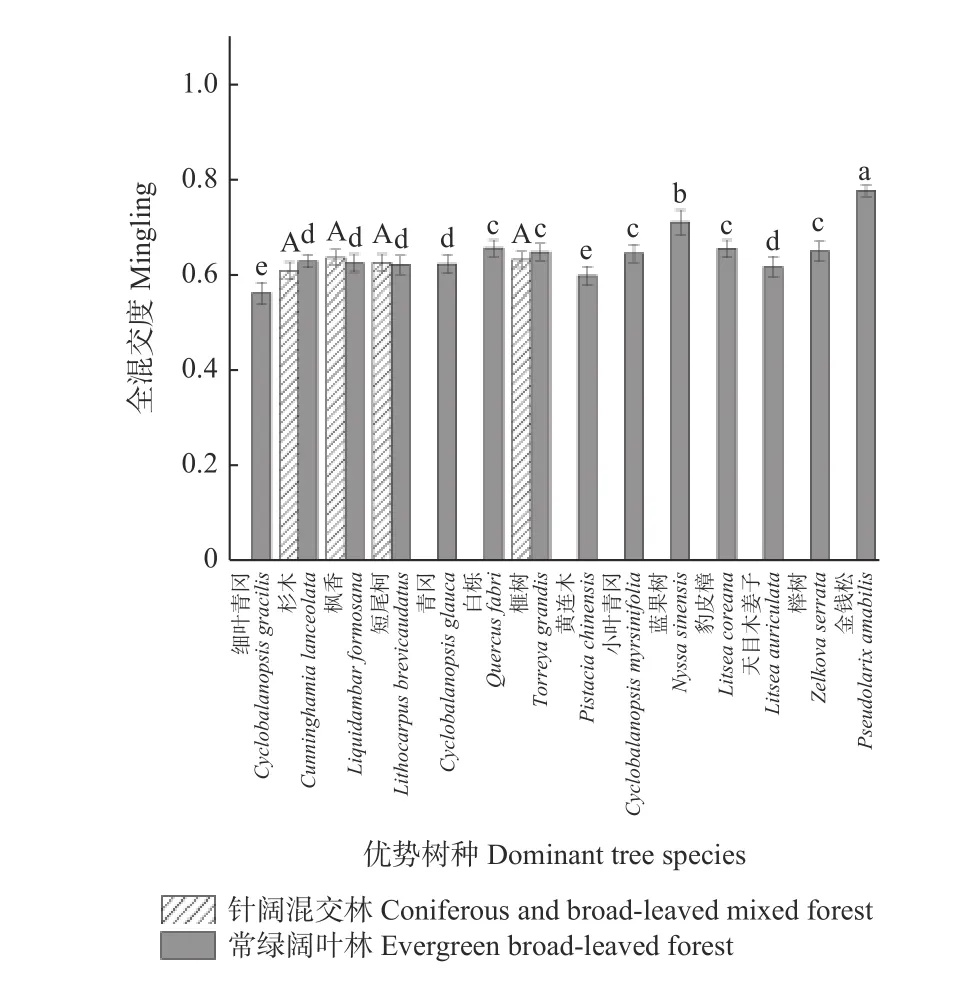
图5 2 种森林群落优势树种混交度比较Fig. 5 Mingling of dominant tree species in two forest communities
3.2.4 2 种森林群落优势树种竞争强度比较 2 个森林群落的优势树种V_Hegyi 竞争指数见图6。在常绿阔叶林14 个优势树种中,有9 种呈现中度竞争,3 种呈现强度竞争,2 种呈现低度竞争。针阔混交林优势树种的竞争指数较小,4 个优势树种分别有2 种强度竞争和2 种低度竞争。在常绿阔叶林中,常绿针叶树种金钱松的V_Hegyi 竞争指数显著大于其余优势树种。从针阔混交林发展到常绿阔叶林,优势树种的组成在原有树种的基础上增加了10 个树种,且原有优势树种的V_Hegyi 竞争指数也有明显增加。

图6 两森林群落优势树种竞争指数比较Fig. 6 Competition index of dominant tree species in two forest communities
4 讨论
通过比较分析天目山针阔混交林和常绿阔叶林的各空间结构指数,发现针阔混交林中的优势树种在常绿阔叶林中同样占有较高优势地位,但常绿阔叶林群落整体的竞争程度更强,分布更倾向随机分布,针阔混交林则有更高的混交度。吴鞠等(2018)在研究林分密度和混交度对长白山天然风景林树木形态的影响时,提出当森林群落混交度较低时,可通过适当提高混交度和密度来提升森林空间利用效率,使整体群落结构更加稳定。因此,在森林经营中,可通过每木调查,找出林地中混交程度较弱的部分,通过人为措施如补植不同树种,适当提升群落整体混交度。
树种组成更丰富的常绿阔叶林的混交度低于针阔混交林,其原因与树种繁殖方式相关。常绿阔叶林中,壳斗科植物总体优势地位较高,壳斗科植物是聚集繁殖的,导致混交度下降,且加剧竞争,也正是这种聚集繁殖方式确定了其在常绿阔叶林中的优势地位。汤孟平等(2009)在研究天目山常绿阔叶林的混交度时指出,细叶青冈等壳斗科优势树种是造成常绿阔叶林整体混交度低的原因之一。从研究的优势树种顺序可看出(表3),细叶青冈在常绿阔叶林中优势地位最高。可见,森林空间结构及其动态变化与树种繁殖方式密切相关。因此,在森林经营或人工林改造过程中,不可忽视树种的生物学特性,盲目追求高混交度,而应根据当前群落演替阶段及树种繁殖特性分阶段设置目标,逐渐促进森林进展演替。
林木空间分布格局是影响森林发展的重要因素之一,它是由森林中多种环境因子综合决定的(操国兴等,2003)。关于森林群落的林木空间分布格局随演替阶段的变化,学者们有不同观点。一般情况下,天然林随森林群落演替过程的进行,林木空间分布格局存在向均匀分布发展的趋势(胡艳波等,2003)。但张家城等(1999)在研究群落优势树种在演替过程中的变化时指出,处于演替顶级阶段的群落在发育过程中,各优势树种的种群分布表现为由集群分布减幅波动地扩散为随机分布,最终在顶级群落中呈现随机分布格局。经多位生态学者研究证实,森林当中物种分布的优势格局多数为聚集分布格局,如广州鼎湖山(叶万辉等,2008)、浙江古田山(李立等,2010)、加蓬热带森林(Nestoret al., 2019)。陈婷婷等(2018)在分析武夷山常绿阔叶林空间结构特征时发现,森林群落整体呈强度混交的轻度聚集分布,有较高的树种异质性,并推断群落演替已达中后期。武夷山与天目山均处于亚热带地区,且优势树种组成类似。武夷山地区常绿阔叶林的优势树种主要由壳斗科和山茶科组成,天目山地区常绿阔叶林优势树种则主要有壳斗科组成。两个森林群落在小径级(5~10 cm)聚集程度均较高,这与张健等(2007)和潘磊磊等(2019)研究发现小径级林木呈聚集分布的结论一致。从天目山地区群落整体看,针阔混交林聚集指数为1.004,常绿阔叶林聚集指数为0.943(表2)。因此,可以认为针阔混交林仍处于演替的前期阶段,其林木呈随机分布;常绿阔叶林处于演替的中后期,其林木呈聚集分布;随着针阔混交林向常绿阔叶林演替,其林木分布格局将向聚集分布发展。
5 结论
通过比较分析浙江天目山国家级自然保护区针阔混交林和常绿阔叶林的空间结构,得出以下结论。
1)常绿阔叶林的株数和树种数均显著高于针阔混交林,林木个体的径级结构呈现倒“J”型分布。
2)常绿阔叶林呈现弱度聚集状态,针阔混交林呈弱均匀状态。2 个森林群落的全混交度均为中度混交,常绿阔叶林的树种数量多于针阔混交林,但全混交度却低于针阔混交林。常绿阔叶林的整体竞争强度大于针阔混交林。
3)优势树种的空间结构指数在针阔混交林均无显著差异,而在常绿阔叶林有较大差异。
4)常绿阔叶林处于演替中后期阶段,针阔混交林处于演替前期阶段。
5)在森林经营中,对常绿阔叶林有必要加强对有聚集繁殖特性的优势树种的抚育,增加群落混交度,降低群落竞争强度和聚集程度,以提高群落稳定性;对针阔混交林则可采取抚育结合促进更新的措施,增加群落密度和树种多样性,提高群落混交度和林木空间分布均匀度,以促进向常绿阔叶林的演替。
猜你喜欢
阿来研究(2020年1期)2020-10-28
临床检验杂志(电子版)(2020年1期)2020-04-03
绿色中国(2019年18期)2020-01-04
新农民(2019年9期)2019-02-19
农家科技下旬刊(2017年8期)2017-11-13
新世纪水泥导报(2016年1期)2016-07-01
学苑创造·B版(2015年12期)2016-06-23
中央社会主义学院学报(2016年2期)2016-05-04
福建农业科技(2015年3期)2015-02-27
华东师范大学学报(自然科学版)(2014年1期)2014-04-16
