
大负荷运动后骨骼肌核心时钟基因Bmal1/Clock、Cry1/Per1的节律性振荡变化趋势
2023-08-07 01:11丁海丽傅泽铤夏雨
中国体育科技 2023年7期
丁海丽,傅泽铤,夏雨
生命的各种行为、生理和代谢功能都具有生物节律。生物节律是指生命活动按一定时间顺序、周而复始地发生变化的节律,主要由生物钟和外界环境信号共同调控(Dibner et al., 2010)。生物钟本质上是一组构成自主节律振荡的时钟基因,其周期性特异表达形成的“转录—翻译—反馈”环路(transcriptional-translational feedback loop,TTFL)是生物节律核心分子机制(Harfmann et al., 2015;Takahashi, 2017)。系列研究表明,哺乳动物时钟基因变异与肠道菌群稳态失衡、肾功能异常、呼吸道过敏性疾病、骨代谢疾病等密切相关(程冯丽 等,2021;刘程程 等,2021;熊勇红 等,2020;余明芳 等,2020;Thaiss et al., 2014;Voigt et al., 2016)。人体的生物节律尤其是昼夜节律能够影响运动能力及运动表现,运动也能对生物节律产生反作用。骨骼肌作为运动的动力结构和直接效应器官,可受到时钟基因调节参与各项生理功能与活动(Mayeuf-Louchart et al., 2015)。而运动作为一种授时因子(zeitgeber)能够影响骨骼肌节律变化,同时也受到骨骼肌固有时钟的调控。因此,明确运动训练后骨骼肌时钟基因的变化趋势、在观测竞技体育及运动促进健康的时间生物学效应、探讨择时训练的内在机制、改善生物钟失调导致的节律紊乱等方面具有重要意义。
在生物体内,生物钟通过时钟基因反馈回路自主产生昼夜节律。研究发现,大脑/肌肉芳香经受体核转位因子样蛋白1(brain and muscle ARNT-like1, Bmal1)、昼夜运动输出周期蛋白(circadian locomoter output cycles protein kaput, Clock)、Period(Per)及Cryptochrome(Cry)等基因对哺乳类动物的生物钟调控起着重要的作用(Keleher et al., 2014)。哺乳动物体内的生物时钟主要由2个转录翻译反馈环路组成,包括核心环路和稳定环路。核心环路的关键结构是转录因子Bmal1和Clock产物形成的异二聚体(Crnko et al., 2019),Bmal1/Clock在TTFL中起正向调控作用,能够与Per和Cry等其他时钟基因的启动子序列结合,激活其特异性表达并形成Per/Cry复合物,经蛋白质定向转运返回细胞核,可反抑制Bmal1/Clock活性,从而以负反馈方式调控自身基因的转录(Curtis et al.,2014)。
运动表现能力与时间生物学因素密切相关,时钟基因是导致运动表现差异的原因之一,Cry1/2作为过氧化物酶体增殖物激活受体δ(peroxisome proliferator-activator receptor,PPARδ)共抑制因子可调控运动能力(Jordan et al.,2017);Per1/2双敲除小鼠在择时运动中并未显示出运动持续时间和血糖水平的差异(Ezagouri et al., 2019);肌肉特异性Bmal1基因敲除可致小鼠肌纤维类型向I型转变(Vitale et al., 2019)。运动还可以调控骨骼肌分子时钟。研究发现计划性运动可影响骨骼肌自身节律表达,使时钟基因相位前移2~3个小时,而不影响中枢节律变化(Harfmann et al., 2015);去除啮齿类动物骨骼肌运动神经元会导致昼夜节律转录活性失调(Nakao et al., 2015);尽管这些研究初步验证了运动与骨骼肌生物节律的相互关系,但运动训练过程中骨骼肌时钟基因分子机制尚未被完全认识。基于此,本研究采用一次下坡跑建立大负荷运动大鼠模型,检测骨骼肌组织Bmal1、Clock 、Cry1、Per1表达,分析其节律振荡变化趋势,旨在探讨大负荷运动训练与时钟基因以及昼夜节律的关系,为运动表现、运动能力的外周生物节律机制提供实验依据。
1 研究对象与方法
1.1 研究对象
8周龄Sprague-Dawley(SD)大鼠208只,体质量(283.17±4.06)g,由成都达硕生物科技有限公司提供,在成都体育学院SPF级动物实验室分笼饲养,室内温度为22~25 ℃,湿度为60%~70%,自由饮食、饮水。实验室光照与黑暗时间12 h交替,光照起始时间(zeitgeber time, ZT)为8∶00,光照结束时间(ZT12)为20∶00。实验中动物处置符合国家科学技术部2006年颁布的《关于善待实验动物的指导性意见》相关规定,获得成都体育学院伦理委员会批准[(2021)34号]。
1.2 实验分组
大鼠实验前适应性喂养1周后随机分为空白对照(Control,C)组和运动(Exercise,E)组,根据运动72 h内不同取材时相(ZT0、ZT6、ZT12、ZT18、ZT24、ZT30、ZT36、ZT42、ZT48、ZT54、ZT60、ZT66、ZT72),C组分为C0、C6、C12、C18、C24、C30、C36、C42、C48、C54、C60、C66、C72共13个亚组;E组分为E0、E6、E12、E18、E24、E30、E36、E42、E48、E54、E60、E66、E72共13个亚组。
1.3 运动方案
通过一次性下坡跑建立大鼠大负荷运动模型。E组经3天适应性喂养后,先采取2天适应性运动,具体运动方案:第1天:跑台坡度0°,运动速度16 m/min,运动时间5 min;第2天:跑台坡度0°,运动速度16 m/min,运动时间10 min;第3天不运动。同时C组正常喂养。适应性训练完成后第4天清晨于6∶30开始运动,坡度-16°,速度16 m/min,运动时间90 min,跑完即刻与ZT0(08∶00)时相相同。
1.4 取材与样本采集
按照不同时间进行分批取材。实验大鼠称重后腹腔注射10%水合氯醛(3.5 mL/kg)麻醉,腹主动脉取血。用有齿镊夹起大鼠腹股沟处皮肤,剥离多余结缔组织,暴露完整肌肉,并将皮肤分离至跟腱部位,用止血钳夹住大鼠足部,将跟腱翻折暴露,手术刀划断跟腱并用齿镊提起,用手术刀划开外层腓肠肌暴露出深层比目鱼肌,冰冷生理盐水漂洗,滤纸拭干,剔除结缔组织。所有时相取右侧组织置于液氮迅速冷冻后-80 ℃冰箱保存,待测RT-PCR实验;每24 h取左侧2 mm×3 mm组织块投入2.5%戊二醛固定液,制备电镜样品。
1.5 透射电镜观察肌纤维超微结构
采用透射电镜(型号JEM-1400PLUS,日本电子JEOL生产)观察比目鱼肌的肌原纤维特征,包括肌小节排列顺序、肌节明/暗带分布情况、线粒体改变情况。将骨骼肌切段后进行样品预固定(3%戊二醛),接着再固定(1%四氧化锇),经丙酮逐级脱水,按比例设置脱水剂的浓度(在100%浓度中更换3次),处理后的骨骼肌样品使用脱水剂,后通过环氧树脂包埋,制备约50 nm切片后,漂浮于刀槽液面上,再捞至铜网,分别通过醋酸铀和枸橼酸铅双染色10~15 min和1~2 min,透射电镜观察采图。
1.6 实时荧光定量PCR检测核心时钟基因mRNA表达
将100 mg组织在液氮中磨碎,加入1 mL已预冷的TRIzol,依次采用氯仿、异丙醇、75%乙醇经反复离心分离提纯样本总RNA,经超微量紫外分光光度计和琼脂糖凝胶电泳检测浓度和纯度后,将RNA逆转录合成cDNA。
采用PIKORed 96(ThermoFisher生产,美国)实时荧光定量PCR测定Bmal1、Clock、Cry1、Per1的mRNA表达水平,PCR反应体系与反应条件见表1、表2。所有引物均交由上海生工生物工程技术服务有限公司设计合成,并以ULTRAPAGE纯化,引物序列见表3。使用Thermo Scientific PikoReal软件分析PCR过程各检测样本的CT(Threshold cycle)值。通过2-△△CT计算X相对mRNA表达水平。

表1 PCR反应体系Table 1 PCR Reaction System

表2 PCR反应条件Table 2 PCR Reaction Conditions
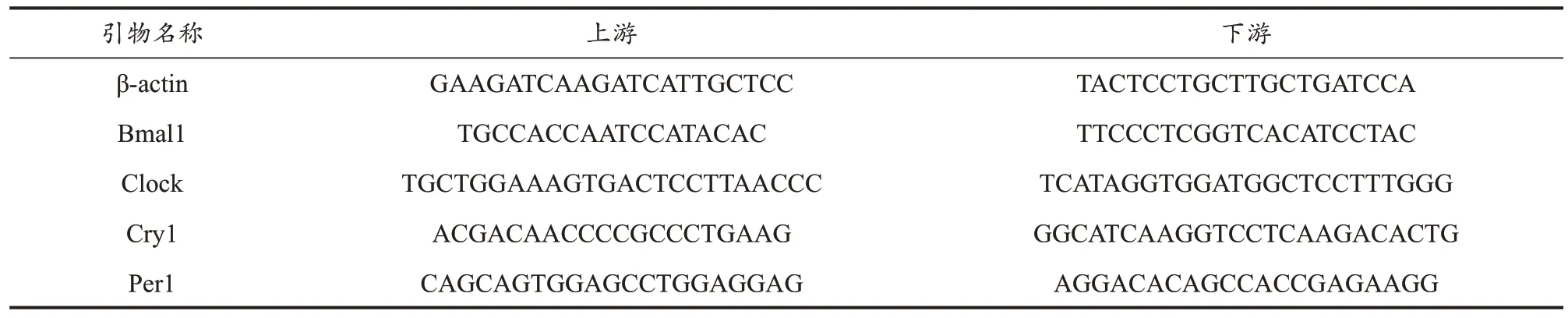
表3 PCR引物序列Table 3 PCR Primer Sequence
1.7 数据统计与分析
所得数据以平均数±标准差(M±SD)表示。使用GraphPad Prism 8.0统计软件进行数据处理。对所得数据进行正态性检验,比较符合正态分布的两组独立样本时采用Student’st检验,方差齐性时采用LSD检验,方差不齐时用Welch’s校正检验,P<0.05表示差异具有显著性。
应用余弦分析软件circacompare (R package) 获取拟合余弦曲线参数,其拟合的余弦函数方程为Y=Mesor+Amplitude*cos(time_Radians-φ),其中Mesor为基线/中值;Amplitude为节律震荡的振幅;time_Radians为时间所对应的弧度值;φ为峰值相位。
2 研究结果
2.1 各组骨骼肌核心时钟基因表达的节律分析
C组拟合余弦函数曲线显示,Bmal1 mRNA于ZT0.25处于峰值,随后下降到ZT12.25为谷值,然后回升至ZT24完成一个周期节律振荡;Clock mRNA于ZT5.43处于峰值,随后下降到ZT17.43为谷值,然后回升至ZT24完成一个周期节律振荡;Cry1 mRNA于ZT1.20处于谷值,随后上升到ZT13.20为峰值,然后回落至ZT24完成一个周期节律振荡;Per1 mRNA于ZT0.42处于谷值,随后上升到ZT12.42为峰值,然后回落至ZT24完成一个周期节律振荡。在之后2天中, Bmal1、Clock、Cry1、Per1 mRNA的变化趋势与0~24 h相似,72 h内共呈现3个完整近日节律(图1~4,表4)。
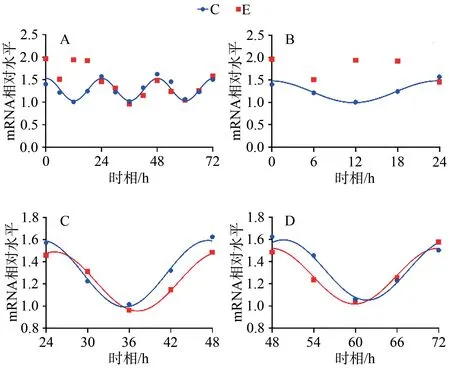
图1 Bmal1 mRNA 表达及拟合余弦函数节律性变化Figure 1. Bmal1 mRNA Expression and the Rhythmic Changes of the Fitted Cosine Function
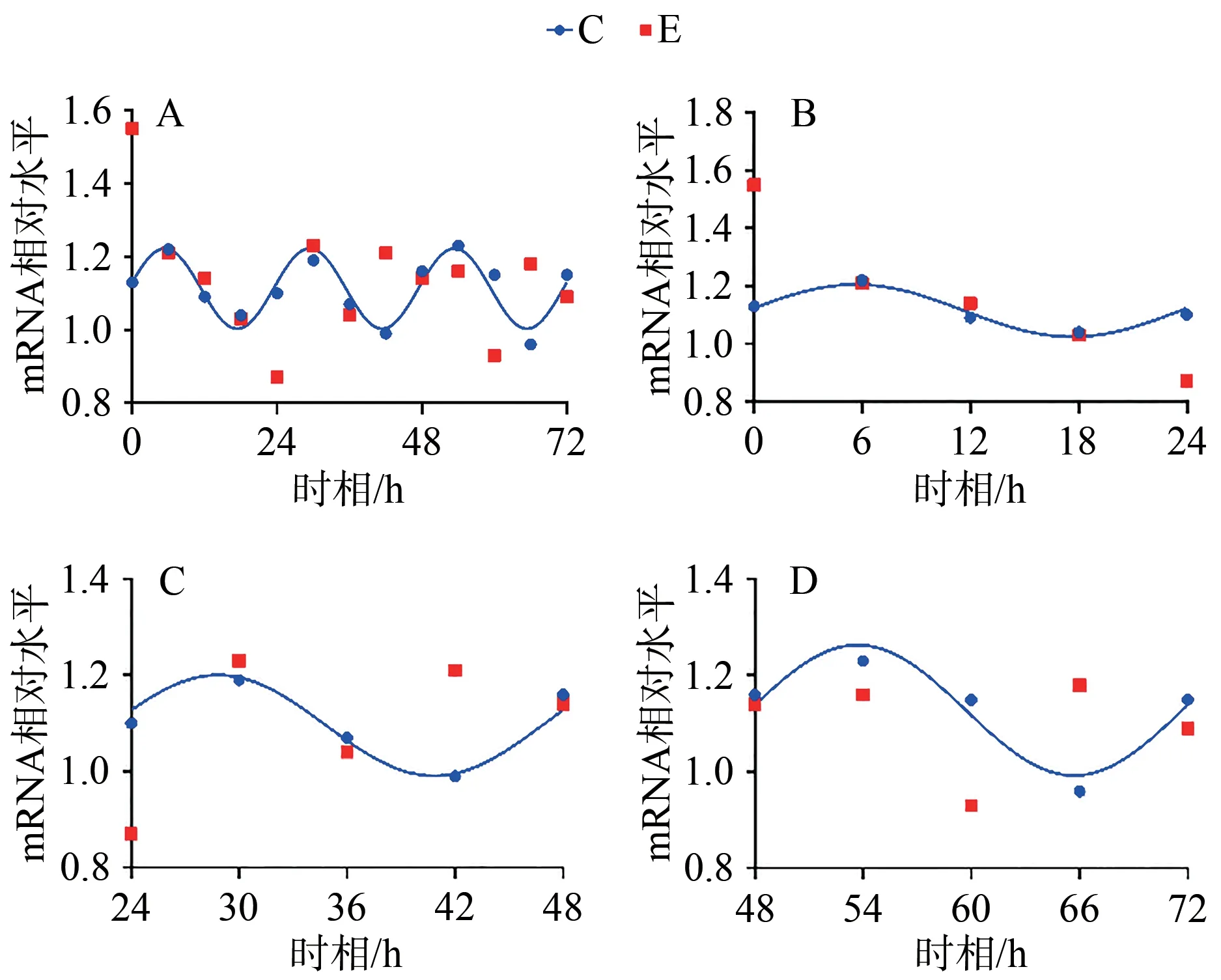
图2 Clock mRNA 表达及拟合余弦函数节律性变化Figure 2. Clock mRNA Expression and the Rhythmic Changes of the Fitted Cosine Function
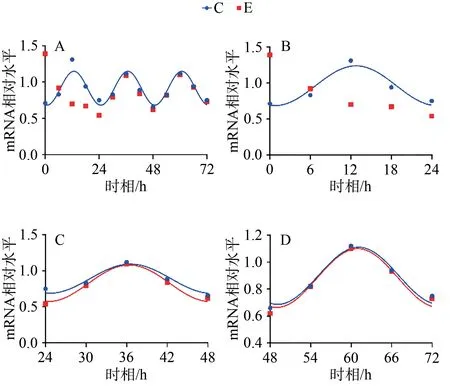
图3 Cry1 mRNA 表达及拟合余弦函数节律性变化Figure 3. Cry1 mRNA Expression and the Rrhythmic Changes of the Fitted Cosine Function
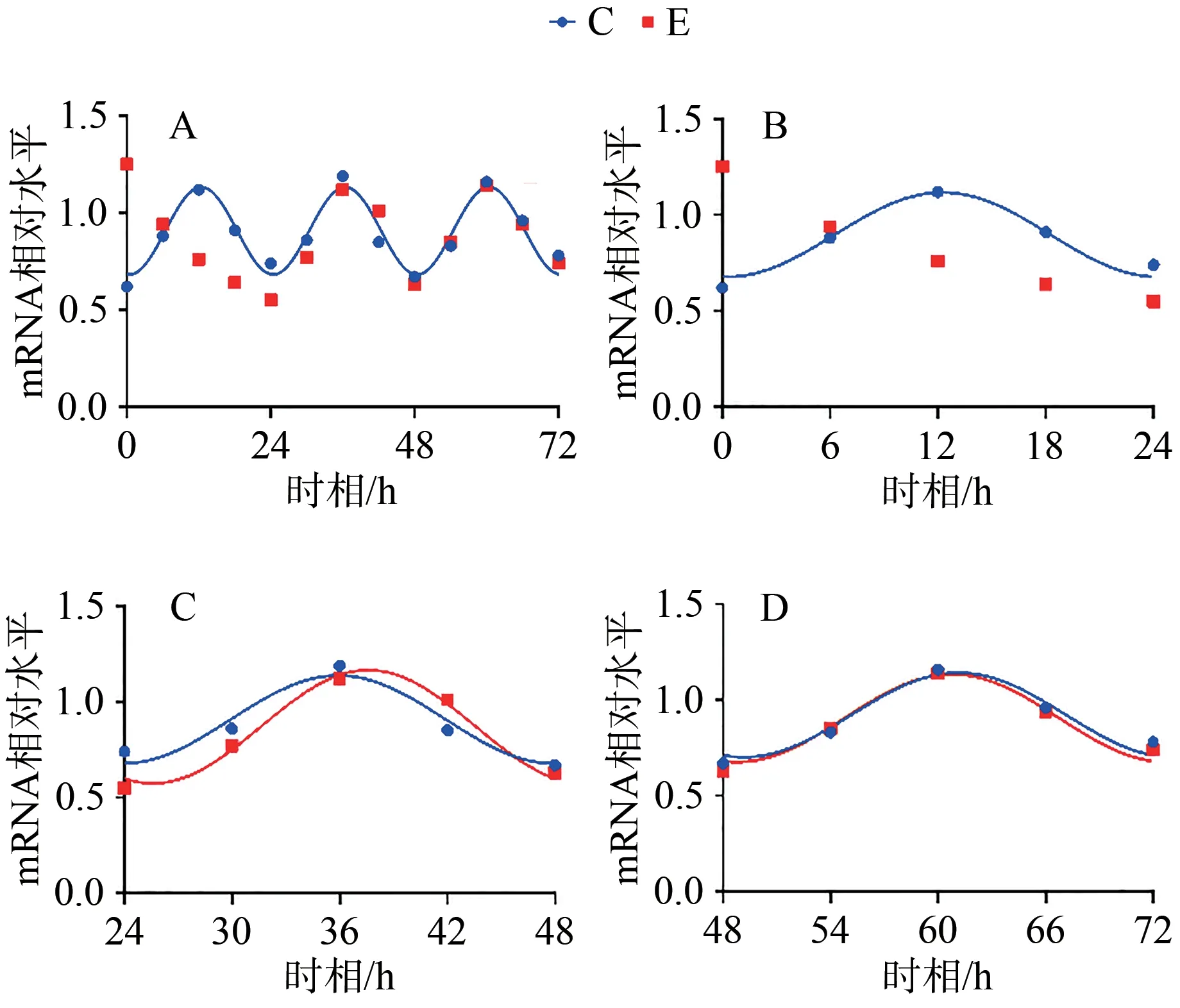
图4 Per1 mRNA 表达及拟合余弦函数节律性变化Figure 4. Per1 mRNA Expression and the Rhythmic Changes of the Fitted Cosine Function
E组拟合余弦函数曲线显示,Bmal1 mRNA在0~24 h近日节律消失,24~48 h与48~72 h恢复近日节律;Clock mRNA在0~72 h近日节律消失;Cry1 mRNA在0~24 h近日节律消失,24~48 h恢复近日节律;Per1 mRNA在0~24 h近日节律消失,24~48 h恢复近日节律(图1~4,表4)。
2.2 各组对应时相骨骼肌核心时钟基因表达的比较分析
统计结果显示,E组Bmal1 mRNA呈现先升高,后趋于正常的状态,在0、6、12和18 h显著高于C组对应时相(P<0.05);Clock mRNA变化趋势与Bmal1 mRNA相似,在0 h差异显著(P<0.05);Cry1 mRNA在0、12 h显著低于C组对应时相(P<0.05);Per1 mRNA变化趋势与Cry1 mRNA基本相同,在0、12 h显著低于C组对应时相(P<0.05;表4,图5)。
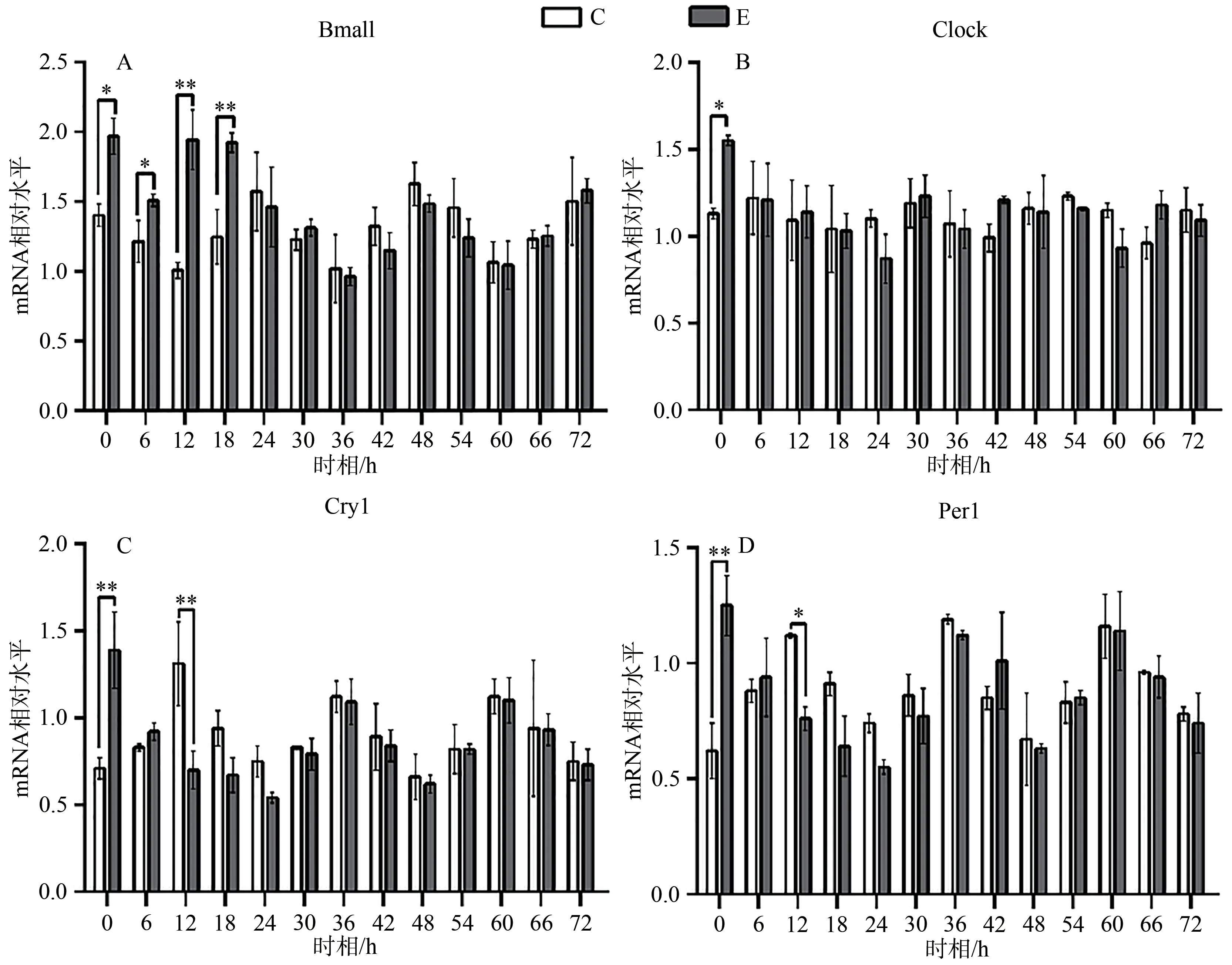
图5 E组与C组同一时相Bmal1、Clock、Cry1、Per1 mRNA的相对表达变化Figure 5. Relative Expression Changes of Bmal1, Clock, Cry1 and Per1 mRNA in Group E and Group C at the Same Timepoint
2.3 各组骨骼肌纤维超微结构变化
透射电镜下观察肌纤维超微结构(图6)发现,C组在0、24、48和72 h各时相肌原纤维分布均匀,A带与I带分界清楚、排列规则,Z线清晰,线粒体排列整齐、形态完整。E组0、24和48 h出现不同程度的Z线加宽、断裂,A带与I带分界不清,线粒体肿胀并聚集在肌膜下,72 h虽有所缓解,但仍未完全恢复。

图6 各组大鼠骨骼肌纤维超微结构Figure 6. Ultrastructure of Skeletal Muscle Fibers in Each Group of Rats
3 分析讨论
昼夜时间(circadian time,CT)0时,Bmal1和Clock形成异二聚体,作用于Cry、Per等靶基因的 E 反应元件(Ebox,CACGTG)并启动转录过程,Cry基因的表达产物和Per基因的表达产物可形成复合物,在细胞核中不断积蓄并反馈抑制Bmall/Clock异二聚体驱动的E-box表达。CT12—CT24期间,Per和Cry的mRNA水平下降,现有Per/Cry复合物降解,以上过程进行约24 h周期循环,形成了生物钟调控网络的TTFL(黄增浩 等,2020)。本研究对未施加干预的对照组SD大鼠骨骼肌Bmal1和Clock、Per和Cry的mRNA进行检测,并获取拟合余弦曲线参数分析其变化趋势,发现72 h内上述基因均呈现3个完整近日节律周期,即昼夜节律振荡。进一步分析其振荡曲线的峰值与谷值,发现Bmal1在CT0—CT12期间由波峰降至波谷,在CT12—CT24期间回升至波峰,Clock的表达趋势与Bmal1基本一致,而Per1和Cry1的拟合曲线则与Bmal1和Clock呈现反向振荡,以上4个基因周期性特异表达形成的振荡规律与生物节律核心分子机制——TTFL相吻合。经转录组学研究发现,骨骼肌中3.4%~16.0%的基因是以节律性方式表达,其表达的Baml1、Rev-erbα、Clock等时钟基因不仅可维持昼夜节律稳定调节,而且通过激活相关信号转导通路或其他作用途径,参与调控骨骼肌机能(Perrin et al., 2018)。本研究基于骨骼肌时钟基因特异性节律表达,主要检测和分析了Bmal1/Clock、Cry1/Per1 mRNA昼夜变化趋势,实验结果进一步佐证了骨骼肌组织作为外周时钟的关键作用,表明骨骼肌表达的核心时钟基因参与维持昼夜节律的稳定输出。
骨骼肌不仅作为外周组织表达时钟基因维持生物节律,还受到时钟基因调控参与各项生理活动。运动表现、运动能力与时间密切相关(黄华生 等, 2016; Faull et al.,2015),有研究发现跨时区产生的时差变化可影响运动表现能力,与向西跨经度旅行相比,向东跨经度旅行可使人体在间歇性短跑运动中表现下降、疲劳感更强(Fowler et al.,2017)。时钟基因是运动表现差异的原因之一,人体实验研究发现,以股四头肌为测试对象,受试者在进行45 min抗阻训练后6 h和18 h,Bmal1、Cry1和Per2的mRNA表达显著升高(Zambon et al., 2003);70 min、70% V.O2max强度的自行车运动后4 h和8 h,股外侧肌Bmal1 mRNA表达显著升高,而Per1 mRNA表达显著降低(Popov et al., 2018);15 min 80% V.O2max强度的自行车运动后60 min,股外侧肌中Per1和Per2的表达增加1.5倍,而Bmal1和Cry1没有变化(Small et al., 2020)。可见,不同形式、不同强度以及不同时间的一次运动后,人体骨骼肌中核心生物钟基因的表达变化并不一致(严露 等,2021)。深入了解运动与骨骼肌时钟基因节律振荡的关系,有利于深入探究肌肉运动能力的时间调节机制、择时运动如何增强运动效果以及过度训练如何影响机能状态。本研究采用可诱发骨骼肌微损伤的一次离心运动经典模型,试图明确大负荷运动能否影响骨骼肌时钟基因固有节律表达。实验结果发现,Bmal1、Cry1、Per1在运动后第1天(0~24 h)均出现近日节律消失,第2天(24~48 h)和第3天(48~72 h)基本恢复,相较而言,Clock最为敏感,72 h内的3个近日节律均受到破坏,表明一次大负荷离心运动可诱发骨骼肌核心时钟基因节律振荡紊乱,但因具体分子钟的不同,其周期性变化呈现特有的振幅和相位表达。
昼夜节律时钟基因大都存在亚型而显示功能冗余,如Cry与Per家族中Cry1、Cry2、Per1、Per2,若敲除单个基因并不会出现明显表型变化,而敲除Cry1/2、Per1/Per2双重基因才能破坏相应基因功能(Aoyama et al., 2017)。Bmal1作为一种非冗余时钟基因,敲除后可导致生物钟完全紊乱(Schiaffino et al., 2016)。Bmal1/Clock异二聚体、Cry/Per复合物是TTEL核心环路的关键结构,因此,需要在整体分析其作用效应的同时,进一步分析各分子的具体变化。本研究中,一次大负荷运动后Bmal1在0~24 h近日节律消失,且无论是光照还是运动刺激后,均在CT0左右即出现显著变化,而Clock在0~72 h内3个近日节律全部消失,推测Bmal1可能作为始动因子,首先影响Bmal1/Clock异二聚体结构,进而调控Cry/Per复合物,形成核心环路4个分子钟的周期性表达变异,在此过程中,Clock可能具有对运动更为敏感的特异性。然而,大负荷运动后时钟基因出现上述节律振荡紊乱的具体作用机制及其带来的后续效应,仍需要进一步探究。
一次大负荷运动模型可诱发运动性骨骼肌损伤,有研究认为骨骼肌质量与肌纤维结构、功能受到时钟基因调控。Clock-/-小鼠可致骨骼肌肌丝结构破坏,单个肌纤维肌力水平下降(Andrews et al., 2010);Bmal1-/-小鼠骨骼肌粗细肌丝超微结构破坏,线粒体肿胀、体积减小(Dyar et al.,2016),但具体途径尚不明确,是否和生物节律中断或时钟基因振荡紊乱相关,还是中枢或其他外周组织的继发效应所致有待进一步探索。本研究在每个近日节律周期始末观察肌纤维超微结构,同时分析比较E组与C组对应时相基因表达差异,结果发现,在近日节律表达均受到破坏的0~24 h,E组相较C组出现显著变化,肌纤维超微结构损伤在0、24 h也较为明显,24~48 h时Clock节律未恢复且Bmal1节律发生偏移,而48 h时相的肌纤维超微结构仍有损伤现象,在48~72 h,随着大部分核心时钟基因近日节律的恢复,肌纤维超微结构损伤也明显减轻,表明时钟基因以及骨骼肌生物节律可参与运动诱发的肌纤维结构改变,机体生物钟与外界生物钟失去同步性有可能加速运动性骨骼肌损伤的发展。综上,本研究通过检测Bmal1/Clock、Cry1/Per1的mRNA表达,分析时相变化及其与肌纤维结构损伤的关系,初步阐释了大负荷运动后骨骼肌核心时钟基因的节律性振荡趋势,为运动员对抗或减轻大运动量训练后时钟基因变化对机体的影响,科学合理安排训练计划并预防运动损伤,以及应对跨时区、洲际参赛引起的昼夜节律紊乱等问题提供了实验依据与参考。
4 结论
大鼠骨骼肌核心时钟基因Bmal1/Clock、Cry1/Per1呈现昼夜节律振荡趋势,一次大负荷运动可诱发上述时钟基因节律紊乱,其时相变化与骨骼肌纤维超微结构损伤相一致。
猜你喜欢
中国当代儿科杂志(2023年2期)2023-02-17
中国医学影像学杂志(2021年6期)2021-08-13
中学数学杂志(高中版)(2016年6期)2017-03-01
现代检验医学杂志(2016年3期)2016-11-15
现代电生理学杂志(2016年4期)2016-07-10
北京信息科技大学学报(自然科学版)(2016年5期)2016-02-27
中国卫生标准管理(2015年5期)2016-01-14
法医学杂志(2015年4期)2016-01-06
职业技术(2015年8期)2016-01-05
中国医疗美容(2015年4期)2015-04-27
