
湖南栎类天然次生林林分空间结构对林下幼树物种多样性的影响
2023-07-26 06:15:24邱子阳何海梅朱光玉
中南林业科技大学学报 2023年6期
邱子阳,何海梅,黄 朗,朱光玉
(中南林业科技大学 林学院,湖南 长沙 410004)
森林更新是森林生态系统资源再生产的一个自然生物学过程。在此过程中,森林生物种群中幼树的生长和发育对未来森林群落的结构及其生物多样性具有深远的影响[1-3]。物种多样性是物种水平上的生物多样性,是用一定空间范围物种数量和分布特征来衡量的,较高的物种多样性能增加群落的稳定性,促进森林生态系统功能的发挥[4-7]。常用的物种多样性指标主要有物种丰富度指数、物种均匀度指数和物种多样性指数等。其中,物种丰富度指数通常用一个群落或生境中物种的数量来衡量,Margalef 指数或Gleason 指数经常被用来表示群落中物种的丰富程度,但由于Gleason指数考虑了样地面积,所以选择Margalef 指数可以避免空间信息的干扰[8];物种均匀度表示各物种的个体数量在群落中的分配状况,反映物种组成的均匀程度,主要的均匀度指数有Pielou 均匀度指数、Alatalo 均匀度指数和Hurlbert 均匀度指数等,Pielou 均匀度指数运用较为广泛。李其斌等[9]和周钰淮等[10]均采用Pielou 均匀度指数来代表物种的均匀程度;物种多样性指数是物种的丰富度和均匀度的函数综合表达,Whittaker[11]和Pielou[12]提出了包含重要值、生物量或相对高度等信息的Whittaker 指数,Auclair & Goff指数在此基础上还加入了相对密度[13],包含的信息更加全面。森林更新主要通过幼苗和幼树的更新来实现[14],林下幼树物种多样性的高低反映林分更新能力的强弱[15]。
林下植被物种多样与林分空间结构因子有着紧密联系,现阶段的大量研究发现,林下植被物种多样性林分空间结构对林下植被的物种多样性有显著影响[16-17]。朱光玉等[18]和朱军等[19]研究发现混交度是影响林下灌木和草本的重要因素;陶观护等[20]和刘丽等[21]分别运用相关分析和灰色关联分析法探究了林分空间结构对林下灌、草物种多样性的影响。许多学者运用了多种分析方法探究林分空间结构对林下灌、草物种多样性的影响规律,而关于林分空间结构如何影响林下幼树物种多样性工作的相关报道较少。林下幼树物种多样性对于林分更新具有重要意义,因此,探究林分空间结构对林下幼树物种多样性的影响规律具有非常重要的研究价值。
根据第九次全国森林资源清查结果,栎类林的面积占全国森林的11.95%,栎类林的蓄积占全国森林的9.98%,全国排名均为第一且以天然林为主[22]。栎类也是湖南的主要森林群落之一,具有涵养水源、防治水土流失等生态价值和供制造车船、提供家畜饲料等经济价值[23]。为探讨湖南栎类天然次生林幼树物种多样性的主要林分空间结构驱动因子是什么的科学问题,本研究以湖南栎类次生林为研究对象,采用3 种统计方法,通过对比分析探索乔木层空间结构对幼树物种多样性的影响规律,并量化林分空间结构对幼树物种多样性的影响,以期为通过林分空间结构调整提高栎类天然次生林林下更新幼树物种多样性提供理论依据。
1 研究区概况
湖南省地处中国中南部、长江中游及洞庭湖以南(108°47'~114°15'E,24°38'~30°08'N)。全省土地面积约为21.18 万km2,占我国国土面积的2.2%。湘东地区以山丘为主、湘西地区以山地为主、湘南地区以丘山为主、湘北地区以平原为主、湘中地区以丘陵为主,整个湖南地貌呈向东北开口的马蹄形。境内主要为红壤和黄壤。属于大陆性亚热带季风湿润气候,夏季高温多雨,冬季寒冷干燥。年平均气温16~19℃,降水充沛,年均降水量为1 200~1 700 mm。全省林地面积约1 300 万hm2,占土地面积的61.40%,属于中亚热带常绿阔叶林地带,森林覆盖率为59.57%,生物资源丰富,是中国珍贵的生物基因库之一。研究区域主要乔木树种包括青冈Cyclobalanopsisglauca、鹅耳枥Carpinusturczaninowii、樟树Cimamomum cayphora、枫香Liquidamabarformosana银杏、GinkgobilobaL、马尾松PinusmassonianaLamb等;主要的灌木树种包括山茶Camelliajaponica、箬竹Indocalamustessellatus、厚皮香Ternstroemia gymnanthera等。
2 研究方法
2.1 样地设置和调查
2015 年12 月—2016 年8 月,采用典型取样法,分别在宁乡县青羊湖林场、郴州市五盖山林场、平江县芦头林场、益阳市龙虎山林场以及桑植县八大公山自然保护区等地共设置49 块固定样地(20 m×30 m),在每块样地内建立直角坐标系。利用GPS 测定样地的经纬度坐标和海拔高度,同时记录坡向、坡位、坡度、腐殖质厚度、枯落物厚度、土壤厚度、郁闭度及乔木株数密度等基本情况。对样地内胸径大于5 cm(小于或者等于5 cm 的作为幼树幼苗进行调查记录)的活立木进行每木检尺并编号,记录树种名称、冠幅、树高、胸径及坐标等。依据样地每木测量结果,先计算林分的平均胸径和平均树高,据此,选择栎类单株胸径和树高分别接近林分平均胸径和平均树高的活立木,采用生长锥钻取其木芯计数年轮,并获得其年龄,将此年龄作为林分年龄。记录样地内所有幼树(阔叶树种:1 m <H<3 m;针叶树种:0.3 m <H<3 m)树种名称、冠幅、树高、胸径(或地径)以及平面坐标[24]。
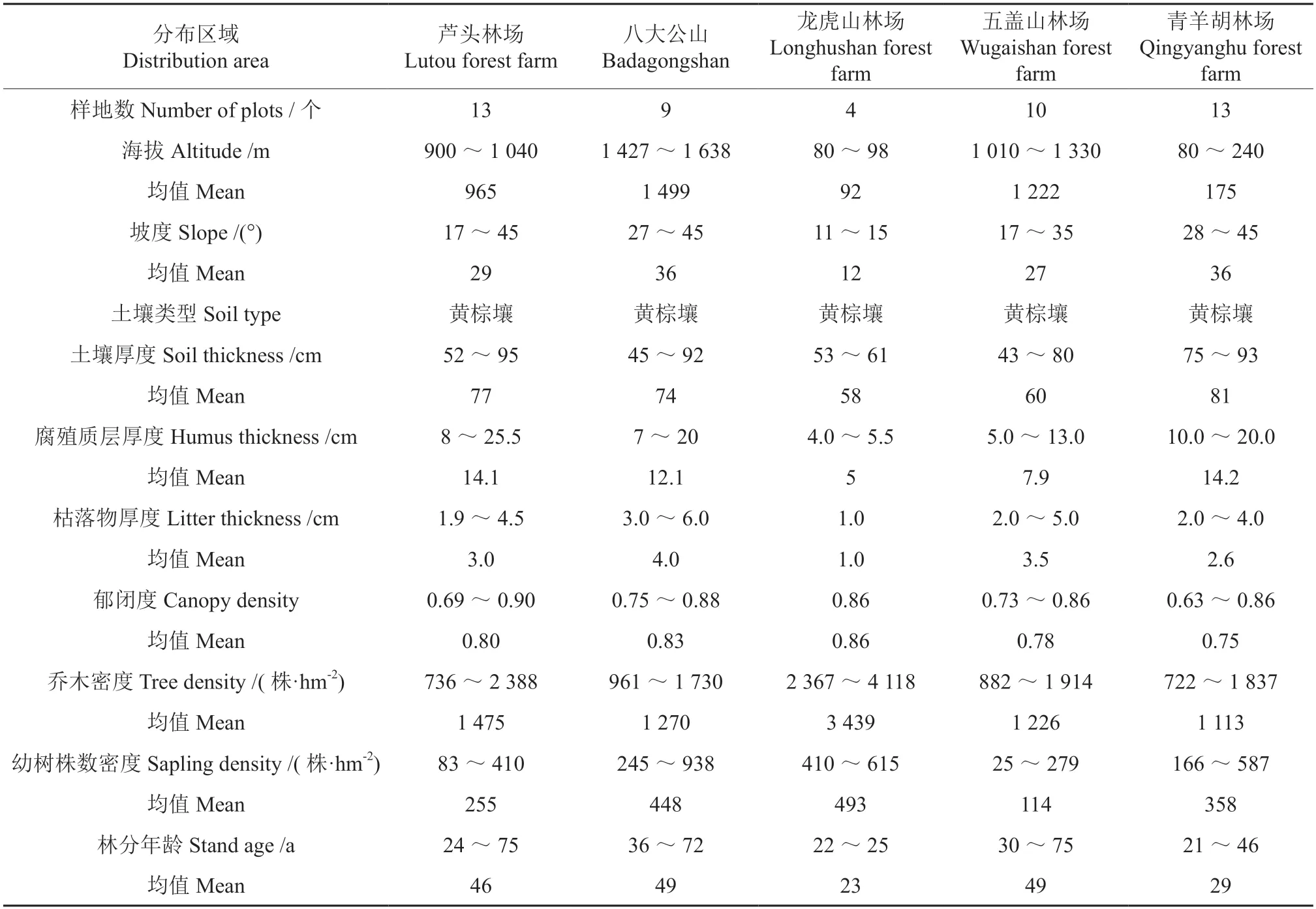
表1 调查样地的基本情况Table 1 Basic situation of the survey plots
2.2 林分空间结构指数的计算
选取林分空间结构参数中的混交度、聚集指数、开敞度(计算方法参考文献[25-27])和树冠指数。
树冠指数(Crown index)代表不同垂直位置冠层分配情况,参照欧芷阳等[28]的方法,将调查样地内树冠层分为主冠层MC(胸径大于12 cm)、下冠层BMC(胸径5~12 cm)和被压层OT(胸径小于5 cm)3 层。根据公式计算得出CI值:
式中:当CI值<1 时,林分树冠面积主要由小树构成;当CI=1 时,说明林分树冠面积在各树冠层平均分配;当CI>1 时,林分树冠面积主要由大树构成。
2.3 林下幼树物种多样性指数的计算方法
在参考相关文献的基础上,从丰富度、均匀度和多样性3 个方面分别选取Margalef 指数、Pielou 指数和Auclair & Goff指数研究林下幼树物种多样性[13,29-30]。
Margalef 指数:
Pielou 指数:
Auclair & Goff指数:
式中:S为样方内所有幼树物种的数目。Ni为幼树物种i的重要值,N为样方内幼树物种重要值的总和。Pi是第i种为相对重要值,Pi=Ni/N[31]。
幼树的物种多样性的重要值公式如下:
2.4 数据分析
首先,利用Pearson 相关分析解释混交度、聚集指数、开敞度、树冠指数、Margalef 指数、Pielou 指数和Auclair & Goff指数的相关关系。其次,为筛选出对物种多样性影响最大的关键指标,并分析各个林分空间结构指标对林下物种多样性的综合作用,采用多元逐步回归分析法,分别以每个幼树物种多样性指标为因变量,林分空间结构指标为自变量,逐个引入新变量。每引入一个新自变量时考虑已进入模型的自变量是否可以被剔除,直至不再引入新的自变量,利用回归统计效果检验指标(AIC,R2,P)选择最优回归模型,揭示多个自变量对因变量的影响[32]。最后,从总体上分析林分空间结构指标和林下幼树多样性指标2 组变量整体之间的相关性,运用典型相关分析(Canonical correlation analysis)构建综合指标,提取相关性最大的线性组合并得到相应的典型相关系数l,以此来判断2 组变量之间的相关性,典型变量相关系数l的显著性检验为Bartlettχ2检验[33]。数据整理与计算采用Excel 2016 和Winkelmass 软件,统计分析及制图在R 4.0.5 软件中进行。
3 结果与分析
3.1 林分空间结构指标与幼树物种多样性指标的计算结果与分异特征
由表2 可以看出,49 块样地的树冠指数、混交度、林下幼树丰富度和均匀度的变化较大,变异系数分别为62.06%、40.82%、61.85%和42.63%。
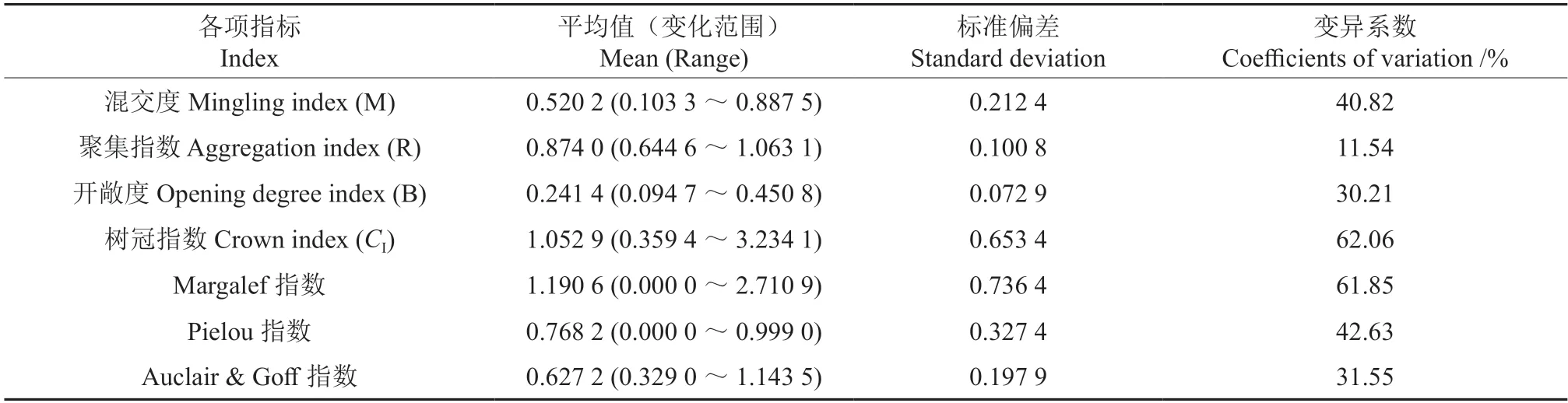
表2 幼树物种多样性指数与林分空间指标的基本数量特征Table 2 General quantitative characteristics of sapling diversity and forest spatial structure
3.2 林分空间结构与幼树物种多样性的Pearson相关分析
对青羊湖林场、龙虎山、芦头林场、八大公山和五盖山的49 块样地测算的林下幼树物种多样性指数以及林分空间指数进行Pearson 相关检验,结果如图1 所示。

图1 林分空间结构与林下幼树物种多样性之间的Pearson 相关关系Fig.1 The Pearson correlation between forest spatial structure and sapling diversity
由图1 可知,丰富度指数与混交度呈极显著相关,相关系数分别为0.387 5;与其余3 个空间结构指标无显著相关关系。均匀度指数与混交度和树冠指数呈极显著相关,其相关系数为0.404 0和-0.386 7;与聚集指数呈显著相关,其相关系数为-0.289 6;与开敞度无显著相关关系。多样性指数与混交度呈极显著相关,其相关系数为-0.487 1;与树冠指数显著相关,相关系数为0.344 0;与其余2 个空间结构指标无显著相关关系。
3.3 多元逐步回归分析结果
在Pearson 相关分析的基础上,以Margalef 指数、Pielou 指数和Auclair & Goff指数为因变量,以混交度、聚集指数、开敞度和树冠指数为自变量进行多元逐步回归,从而保留影响幼树物种多样性的主要解释变量。多元逐步回归分析结果(表3)表明,3 个多样性指数的回归方程都通过了显著性检验(P<0.05),Margalef 指数、Pielou 指数和Auclair & Goff指数回归方程都加入了树冠指数和混交度,这说明幼树物种多样性主要受垂直位置冠层分配情况和乔木层树种空间隔离程度的影响。3 个回归模型的决定系数R2分别为0.208 0、0.303 8 和0.350 1,这说明除了林分空间结构,还有其他因素影响林下幼树的物种多样性。

表3 幼树物种多样性指数与林分空间结构指标的多元逐步回归分析†Table 3 Multiple stepwise regression analysis of sapling diversity and forest spatial structure
3.4 林分空间结构与幼树物种多样性的典型相关分析
由表4 可知,3 对典型变量中只有第一对典型变量在a=0.01 水平上通过了Bartlettχ2检验,相关系数为0.647 1,说明幼树物种多样性指数与林分空间结构指标总体上有较强的相关性。表5 结果表明,第一对典型变量的线性组合是:

表4 典型相关系数和检验结果Table 4 The canonical correlation coefficients and their statistical tests

表5 3 对典型变量的因子载荷Table 5 The loads of three canonical variables
在第一对典型变量中,u1为栎类天然次生林林分空间结构指标的线性组合,其中混交度M的载荷最大,为-0.114 0;树冠指数CI的载荷较大,为0.081 9,说明混交度是影响林下幼树物种多样性的主导因子,树冠指数是重要因子。v1是林下幼树物种多样性指标的线性组合,Margalef 指数的载荷较大,为0.098 2,说明林下幼树的丰富度指数对乔木层林分空间结构指数较为敏感。其中Auclair & Goff指数的载荷最大,为0.212 2,说明林下幼树的多样性指数对乔木层林分空间结构指数最为敏感。
第一对典型变量中带入各样地的原始数据,得分散点图根据各样地得分值画出。从图2 可以看出,u1和v1之间线性相关较明显,两者的得分散点几乎分布于一条直线上,说明林分空间结构指标与幼树物种多样性指标在整体上的相关性显著。
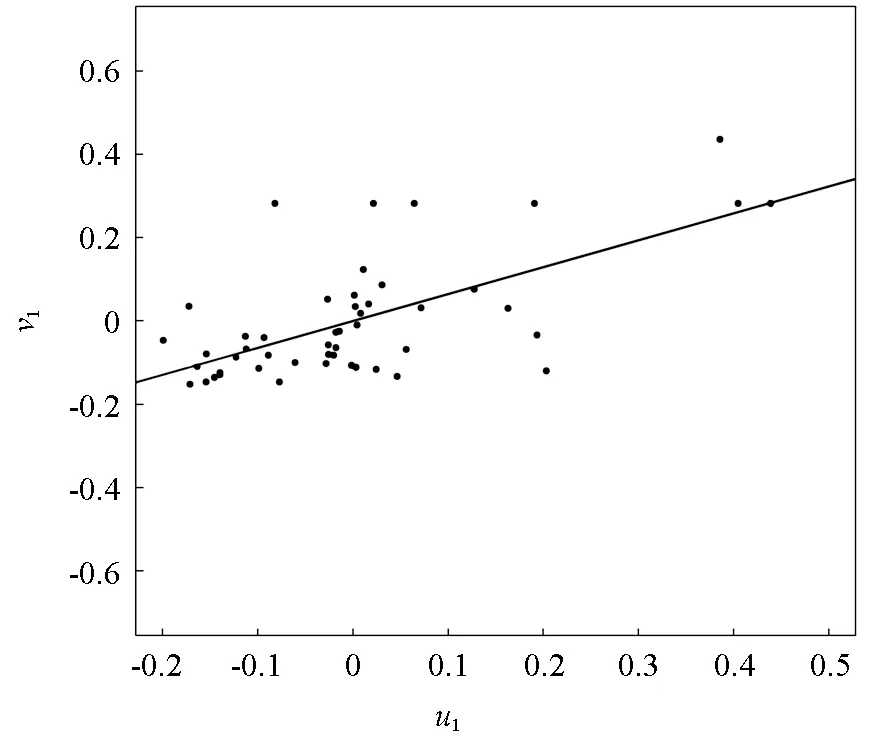
图2 林分空间结构与幼树多样性典型相关得分Fig.2 The score of canonical correlation between spatial structure and sapling diversity
4 结论与讨论
4.1 讨 论
4.1.1 Pearson 相关分析
1)混交度与幼树的丰富度指数、均匀度指数和多样性指数都有很强的相关性。当混交度增大,幼树物种丰富度、均匀度和多样性均增加,这与董莉莉等[34]的研究结果一致。可能是因为混交度的大小会影响林分各个树种的相互竞争程度,不同的树种因为竞争关系在生态位上有不同程度的相互抑制,为林下幼树更新释放了营养空间。因此,混交度高的林分,其更新潜力也大[35]。
2)聚集指数与幼树均匀度指数显著负相关。聚集指数越大,林分的水平分布越趋近于均匀分布[36],可能不利于一些喜光树种的生长发育,影响更新幼树物种的分布均匀程度。
3)开敞度与幼树物种丰富度、均匀度和多样性没有显著相关性。开敞度越大,林下光照条件越好,幼树物种均匀度和多样性反而降低。沈海龙等[37]研究发现林分开敞度在1.0~2.0 比较适宜林冠下红松幼株的生长。本研究调查样地的开敞度较小,其平均值为0.241 4,林分郁闭程度较高,可能不利于林下幼树更新。
4)树冠指数与幼树的物种均匀度指数和多样性指数(Auclair & Goff指数是一个反向指标),分别呈极显著负相关和显著正相关。树冠指数增大,幼树物种多样性和均匀度都减小,这与欧芷阳等[28]研究类似。可能是因为随着树冠指数增大,上层和中间层的树冠总面积增加,导致穿透中、上林冠到达林下的总光照量降低,对幼树获取光资源产生不确定性影响[38]。
4.1.2 多元线性逐步回归分析
Pearson 相关分析探究了各变量之间的相关性,而多元线性逐步回归从多元角度揭示了多个林分空间结构指标对林下幼树物种多样性的影响规律。分别以丰富度指数、均匀度指数和多样性指数做因变量时,都筛选出树冠指数和混交度2 个空间结构指标,说明树冠指数和混交度是影响林下幼树物种丰富度、均匀度和多样性的关键指标。不同垂直位置冠层面积分配比例和林分树种混交程度对林下幼树物种丰富度、均匀度和多样性影响较大。
4.1.3 典型相关分析
研究结果揭示了林分空间结构与幼树物种多样性这2 组变量之间整体的线性相关关系。第一对典型变量的混交度和树冠指数的载荷较大,说明混交度和树冠指数对林下幼树物种多样性影响较大,这与多元线性逐步回归分析的结果一致。曹小玉等[39]采用多元线性逐步回归分析法和典型相关分析法分析林分空间结构对土壤养分的影响,发现2 种分析方法的结果也一致。
目前,关于林下植被物种多样性的研究主要是集中于灌木和草本的物种多样性,而本研究首次发现混交度和树冠指数是影响湖南栎类天然次生林林下幼树物种多样性的主要驱动因子,树种的混交程度越高,林下幼树物种的丰富度、均匀度和多样性均增大;树冠的上层和中间层的总面积增加,林下幼树物种的均匀度和多样性减小。有研究表明海拔等立地因子也会显著影响幼树的物种多样性[40],但在森林经营管理中,海拔、坡度等立地因子难以调控,而林分空间结构是较易于调控的因子,因此,可以采用补植、间伐和疏伐等调控措施来提高树种混交程度和减小树冠的上层、中层的总面积,进而丰富林下幼树的物种多样性和提升林分的更新潜力。
3 种统计分析方法可以全面直观地反映出林分空间结构指标和幼树物种多样性之间的单因素相关性和整体上的综合影响,但只分析了林分空间结构对幼树物种多样性的直接影响规律。在今后的研究中,可以尝试采用结构方程等方法,同时探究林分空间结构对幼树物种多样性的间接和直接的影响规律,进而更深层次地挖掘两者之间的相关关系。另外,本研究是以湖南栎类天然次生林为对象,对于其他树种林下幼树物种多样性和林分空间结构的关系还有待进一步探究。
4.2 结 论
本研究采用3 种统计分析方法探讨了林分空间结构对其幼树物种多样性的影响规律,相关结论如下:
1)丰富度指数与混交度呈极显著正相关;均匀度指数与混交度、树冠指数和聚集指数分别呈极显著正相关、极显著负相关、显著负相关;多样性指数与混交度和树冠指数呈极显著负相关和显著正相关。混交度对林下幼树物种多样性影响最大,树冠指数次之,聚集指数对林下幼树物种多样性的影响最小。
2)分别以丰富度指数、均匀度指数和多样性指数做因变量的3 个多元逐步回归方程,在筛选因子后均只剩下混交度和树冠指数,说明混交度和树冠指数是影响林下幼树物种多样性的主要因子。
3)第一对典型变量的线性组合中,林分空间结构指标中的混交度载荷最大,树冠指数载荷较大,表明混交度对林下幼树物种多样性的影响最大,树冠指数次之。
综合3 种分析方法的研究结论,乔木层林分空间结构中的混交度和树冠指数是影响林下更新幼树物种多样性的主要驱动因子,混交度对幼树物种多样性的影响最大,乔木层林木混交度越高,其林下幼树物种的丰富度指数,均匀度指数和多样性指数越大;树冠指数对幼树物种多样性的影响较大,树冠的上层和中间层的总面积增加,其林下幼树物种的均匀度指数越小,多样性指数越小。因此,可以通过采用调整树种结构为主,综合考虑树冠层结构的森林经营措施来提高栎类天然次生林林下更新幼树物种多样性。
猜你喜欢
阿来研究(2020年1期)2020-10-28 08:10:22
防护林科技(2020年6期)2020-08-12 13:34:40
河北果树(2020年1期)2020-02-09 12:31:18
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
现代园艺(2017年11期)2017-06-28 11:32:57
新世纪水泥导报(2016年1期)2016-07-01 03:59:37
中央社会主义学院学报(2016年2期)2016-05-04 04:18:28
土壤与作物(2015年3期)2015-12-08 00:46:58
亚热带资源与环境学报(2015年3期)2015-01-22 07:05:36
