
肿节风总黄酮调节MAPK/JNK信号通路促进巨核细胞分化成熟
2023-07-11 00:50尚广彬陈中张钟康卢震路千里严小军卢晓南江西中医药大学中医基础理论分化发展中心江西南昌000江西省中医病因生物学重点实验室江西南昌000江西中医药大学临床医学院江西南昌000江西中医药大学中医学院江西南昌000
中药新药与临床药理 2023年4期
尚广彬,陈中,张钟康,卢震,路千里,严小军,卢晓南(1.江西中医药大学中医基础理论分化发展中心,江西 南昌 000;2.江西省中医病因生物学重点实验室,江西 南昌 000;.江西中医药大学临床医学院,江西 南昌 000;.江西中医药大学中医学院,江西 南昌 000)
免疫性血小板减少症(Immune thrombocytopenia,ITP)属于自身免疫性疾病,以外周血小板计数减少为主要特征,其主要的发病机制是由于人体自身免疫耐受机制被打破,体液免疫和细胞免疫紊乱,产生抗自身血小板抗体,引发血小板破坏增多、巨核细胞分化成熟障碍及生成血小板减少[1-2]。其中巨核细胞分化成熟障碍越来越受到关注[3-5],它是指骨髓中巨核细胞数目正常或增多,但分化不足,成熟受阻,产板不良,导致血小板减少,是ITP的特征性改变,也是其发病和治疗的关键。
ITP 属于中医“血症”范畴,以随着外周血小板数量减少,或伴皮肤黏膜出血、紫癜及瘀斑等为特点,中医诊治专家共识建议对各证型ITP 适时、适度使用活血化瘀药物进行干预治疗[6]。肿节风[Sarcandraglabra(Thunb.)Nakai]是金粟兰科草珊瑚属植物,具有活血化瘀、凉血止血的功效,以及抗免疫性血小板减少性紫癜的药理作用[7-8]。本课题组前期研究[9-11]表明,肿节风总黄酮提取物能明显提高成熟巨核细胞的数量和促进巨核系祖细胞集落的形成,并可以通过调节骨髓微环境,改善骨髓巨核细胞分化成熟障碍状态,促进其分化成熟并生成血小板,但具体作用机制尚不清楚。故本研究拟利用体外巨核细胞分化成熟障碍模型,探索肿节风总黄酮通过丝裂原活化蛋白激酶(MAPK)/c-Jun N-末端激酶(JNK)信号通路促进巨核细胞分化成熟的作用机制。
1 材料与方法
1.1 细胞株人类巨核细胞白血病Dami 细胞株,购于中国科学院细胞资源中心。
1.2 药物及试剂肿节风总黄酮提取物,由本实验室前期制备[7]:将肿节风浸膏利用HDP-400 大孔树脂吸附,70%乙醇洗脱,冷冻干燥得到的固形物即为总黄酮,以芦丁作对照检测总黄酮含量为78.53%。兔抗大鼠血小板血清(APS,货号:201108),参考卢晓南等[10]方法由北京鼎国昌盛生物技术有限公司定制;佛波醇12-十四酸酯13-乙酸酯(PMA,货号:P1585),美国Sigma-Aldrich 公司;RPMI Medlium 1640 培养基(货号:C11875500BT),美国Hyclone 公司;胎牛血清(货号:11011-8611),浙江天杭生物科技股份有限公司;青链霉素混合液(货号:P1400),北京索莱宝科技有限公司;化学发光细胞活力检测试剂盒(货号:G9241),美国Promega 公司;Anti-Human CD41a PE(货号:12-0419-42)、Anti-Human CD42b FITC(货号:11-0429-42)、Anti-Human CD61 FITC(货号:11-0619-42),均购自美国 eBioscience 公司;Phospho- SAPK/JNK(Thr183/Tyr185,81E11)Rabbit mAb(货号:4668)、SAPK/JNK Antibody(货号:9252)、Phospho-c-Jun(Ser73,D47G9)XP®Rabbit mAb(货号:3270)、c-Jun(60A8)Rabbit mAb(货号:9165)、β-Tubulin(D2N5G) Rabbit mAb(货号: 15115)、GAPDH(D16H11)XP®Rabbit mAb(货号:5174),均购自美国Cell Signaling Technology 公司;辣根过氧化物酶标记山羊抗兔IgG 抗体(货号:SA00001-0),武汉三鹰生物技术有限公司;ECL 发光液(货号:CW0049S),江苏康维世纪有限公司。
1.3 主要仪器Forma 3111 型CO2培养箱,美国Thermo Scientific 公司;Spark 10M 型多功能酶标仪,瑞士Tecan 公司;Moflo XDP 流式细胞仪,美国BeckmanCoulter 公司;Mini-PROTEAN 蛋白电泳及转印系统、Chemi Doc XRS+化学发光凝胶成像系统,美国Bio-Rad 生命医学产品有限公司。
1.4 体外巨核细胞分化成熟模型的建立、分组及药物干预取状态良好的对数生长期Dami 细胞进行实验,利用终浓度为5 ng·mL-1的PMA 和体积分数为1%的APS 联合作用于Dami 细胞,构建体外巨核细胞分化成熟障碍模型。实验分为空白对照组(Control组)、PMA 诱导组(5 ng·mL-1)、APS 模型组(5 ng·mL-1PMA+1% APS,Model 组)以及肿节风总黄酮高、中、低剂量组(15.6、7.8、3.9 μg·mL-1肿节风总黄酮+5 ng·mL-1PMA+1% APS),细胞接种后将培养板移入37 ℃、5% CO2培养箱中。
1.5 化学发光法检测细胞增殖活力收集对数生长期细胞,以每孔5 000 个细胞(体积100 μL)接种于96 孔板中,分组同“1.4”项下,增加调零组,每组4 个复孔。在37 ℃、5% CO2细胞培养箱中培养48、72、96 h 后,以每孔100 μL 滴加化学发光试剂,摇床振荡反应10 min 后,置于多功能酶标仪中检测500 ms 的Luminesence 读数(L 读数)。计算:细胞相对增殖活力(%)=[(实验孔L 读数-调零孔L 读数)/(对照孔L 读数-调零孔L 读数)]×100%。
1.6 流式细胞术检测巨核细胞分化情况取对数生长期细胞,以每孔6 ×104个细胞(体积1 mL)接种于12 孔板,分组同“1.4”项下,每组4 个复孔。药物干预96 h 后,收集各组细胞至2 mL 离心管,用磷酸缓冲液(PBS)离心清洗2 次,用500 μL PBS 重悬细胞;再加入5 μL Anti-Human CD41a PE、Anti-Human CD42b FITC、Anti-Human CD61 FITC 染液,混匀后在室温下避光染色30 min。用细胞筛网进行过滤后,采用流式细胞仪检测巨核细胞分化成熟的表面标记分子CD41a、CD42b 及CD61 表达情况,结果用FolwJo 7.0 软件进行分析。
1.7 Western Blot 法检测细胞MAPK/JNK 信号通路相关蛋白的表达情况取对数生长期细胞,以每孔1×105个细胞(体积2 mL)接种于6 孔板,分组同“1.4”项下,每组3 个复孔。药物干预96 h 后收集各组细胞,离心清洗后加入200 μL RIPA 裂解液(使用之前加入蛋白酶抑制剂和磷酸化蛋白酶抑制剂),冰上裂解30 min;以13 500×g离心15 min,取上清蛋白液,采用BCA 酶标法测定蛋白浓度。以每孔20 μg 蛋白标准配制,加入6×Loading Buffer,混匀,100 ℃加热10 min。上样进行SDS-PAGE 电泳分离,将蛋白转移至PVDF 膜上;用5% BSA 室温下封闭1 h 后,分别加入Phospho-SAPK/JNK(1∶1 000)、SAPK/JNK(1∶2 000)、Phospho-c-Jun(1∶1 000)、c-Jun(1∶2 000)、β-Tubulin(1∶2 000)、GAPDH(1∶2 000)一抗,4 ℃下孵育过夜;TBST 洗膜3 次,每次10 min,加入辣根过氧化物酶标记山羊抗兔IgG二抗(1∶4 000),室温下孵育1.5 h;TBST 洗膜3 次,每次10 min,采用ECL 发光液于凝胶成像仪显影,扫描;以β-Tubulin/GAPDH 为内参,使用ImageJ 1.53 软件对目的蛋白进行半定量分析。
1.8 统计学处理方法采用GraphPad Prism 8.0 统计软件进行数据分析;计量资料以均数±标准差(±s)表示;多组间比较采用单因素方差分析(One-way ANOVA),两两比较采用LSD 检验;以P<0.05 为差异有统计学意义。
2 结果
2.1 肿节风总黄酮对Dami 细胞增殖活力的影响结果见表1。与空白对照组相比,PMA 诱导组不同时间点的细胞活力均显著降低(P<0.01),与PMA 诱导巨核细胞分化成熟增殖受抑制有关。与PMA 诱导组相比,APS 模型组48 h 的细胞活力明显下降(P<0.05),但随时间延长抑制作用减弱,72、96 h 的差异无统计学意义(P>0.05)。与APS 模型组比较,肿节风总黄酮中、高剂量组48、72 h 的细胞活力均受到明显抑制(P<0.05,P<0.01),肿节风总黄酮高剂量组96 h 的细胞活力受到显著抑制(P<0.01)。结果表明,肿节风总黄酮对体外巨核细胞分化成熟模型Dami 细胞的增殖活性有明显的抑制效应,可能与其促进巨核细胞分化成熟有关。
表1 肿节风总黄酮对Dami 细胞增殖活力的影响(±s,n=4)Table 1 Effect of flavonoids from Sarcandrae glabra on Dami cell viability(±s,n=4)
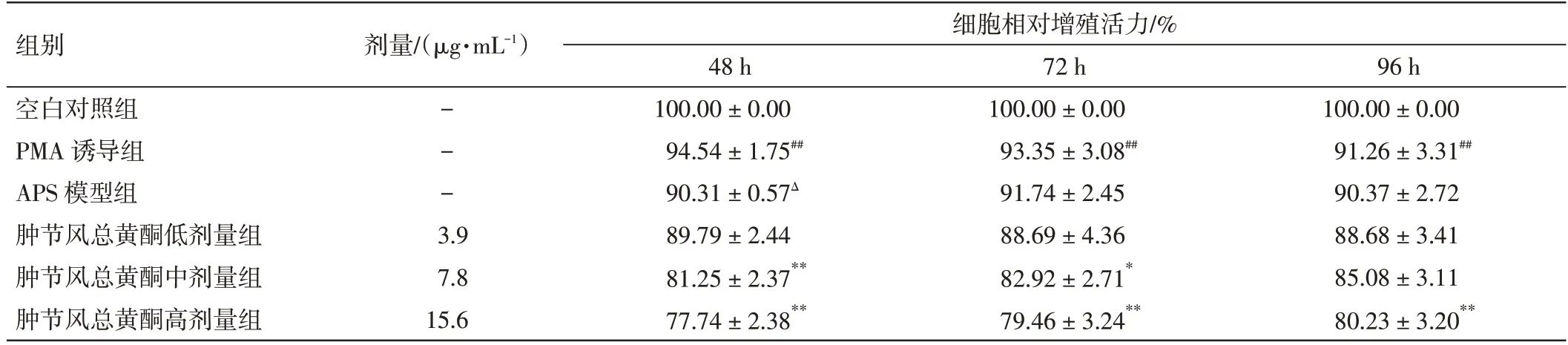
表1 肿节风总黄酮对Dami 细胞增殖活力的影响(±s,n=4)Table 1 Effect of flavonoids from Sarcandrae glabra on Dami cell viability(±s,n=4)
注:与空白对照组比较,##P<0.01;与PMA 诱导组比较,ΔP<0.05;与APS 模型组比较,*P<0.05,**P<0.01
组别空白对照组PMA 诱导组APS 模型组肿节风总黄酮低剂量组肿节风总黄酮中剂量组肿节风总黄酮高剂量组剂量/(μg·mL-1)细胞相对增殖活力/%96 h 100.00±0.00 91.26±3.31##90.37±2.72 88.68±3.41 85.08±3.11 80.23±3.20**---3.9 7.8 15.6 48 h 100.00±0.00 94.54±1.75##90.31±0.57Δ 89.79±2.44 81.25±2.37**77.74±2.38**72 h 100.00±0.00 93.35±3.08##91.74±2.45 88.69±4.36 82.92±2.71*79.46±3.24**
2.2 肿节风总黄酮对巨核细胞分化成熟标志物表达的影响结果见图1、图2、图3。与空白对照组比较,PMA 诱导组Dami 细胞表面的CD41a、CD42b 和CD61 表达水平均显著升高(P<0.01)。与PMA 诱导组比较,APS 模型组Dami 细胞表面的CD41a、CD42b 和CD61 表达水平均显著降低(P<0.01)。与APS 模型组比较,肿节风总黄酮高、中剂量组Dami细胞表面的CD41a、CD42b 和CD61 表达水平均显著升高(P<0.01)。结果表明,利用PMA 及APS 诱导Dami 细胞成功建立体外巨核细胞分化成熟障碍模型,而肿节风总黄酮能够改善巨核细胞的分化成熟障碍状态,促进细胞表面CD41a、CD42b 和CD61表达。

图1 肿节风总黄酮对Dami 细胞表面CD41a 表达的影响(±s,n=4)Figure 1 Effect of flavonoids from Sarcandrae glabra on the expression of CD41a on the surface of Dami cell(±s,n=4)

图2 肿节风总黄酮对Dami 细胞表面CD42b 表达的影响(±s,n=4)Figure 2 Effect of flavonoids from Sarcandrae glabra on the expression of CD42b on the surface of Dami cell(±s,n=4)
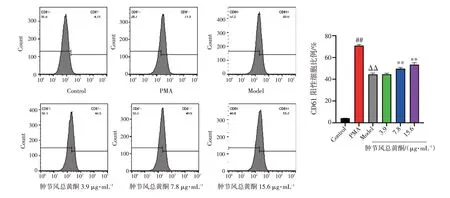
图3 肿节风总黄酮对Dami 细胞表面CD61 表达的影响(±s,n=4)Figure 3 Effect of flavonoids from Sarcandrae glabra on the expression of CD61 on the surface of Dami cell(±s,n=4)
2.3 肿节风总黄酮对Dami 细胞MAPK/JNK 信号通路相关蛋白表达的影响结果见图4。与PMA 诱导组比较,APS 模型组Dami 细胞的p-JNK/JNK 及p-c-Jun/c-Jun 蛋白表达显著上调(P<0.01)。与APS模型组比较,肿节风总黄酮中、高剂量组Dami 细胞的p-JNK/JNK 蛋白表达明显下调(P<0.05,P<0.01);肿节风总黄酮低、中、高剂量组Dami 细胞的p-c-Jun/c-Jun 蛋白表达显著下调(P<0.01)。结果表明,肿节风总黄酮可能通过抑制MAPK/JNK 信号通路的激活,改善体外巨核细胞分化成熟障碍模型的分化受阻状态。

图4 肿节风总黄酮对Dami 细胞p-JNK、JNK、c-Jun 及p-c-Jun 蛋白表达的影响(±s,n=3)Figure 4 Effect of flavonoids from Sarcandrae glabra on the protein expressions of p-JNK,JNK,p-c-Jun and c-Jun of Dami cell(±s,n=3)
3 讨论
免疫性血小板减少症(ITP)属于机体免疫失衡造成的血液性疾病,主要发病机制是由于各种原因引发体液及细胞免疫异常活化,产生抗自身血小板抗体,导致体内血小板破坏增多或/和巨核细胞分化成熟障碍[1]。有研究[3]认为,抗自身血小板抗体与巨核细胞结合,使之分化成熟受阻,无法生成血小板才是血小板减少的主要原因。因此,利用含抗血小板抗体的血清(APS)作为一种成熟的ITP 动物模型诱导剂,能抑制骨髓内巨核细胞分化成熟,有效减少模型动物的血小板,可以较好地模拟ITP 发病时的状态[12-14]。目前对体外巨核细胞分化成熟障碍模型的研究相对不足,因此本研究利用巨核细胞分化经典诱导剂佛波醇12-十四酸酯13-乙酸酯(PMA)作用于体外Dami 巨核细胞,诱导其快速分化成熟,同时结合APS 阻逆巨核细胞分化成熟。结果显示,模型组Dami 细胞表面的巨核细胞分化标志分子CD41a、CD42b 和CD61 表达量显著降低,表明成功构建了体外巨核细胞分化成熟障碍模型;而肿节风总黄酮干预后,可显著上调模型组Dami 细胞表面CD41a、CD42b 和CD61 表达,促进巨核细胞的分化成熟。
丝裂原活化蛋白激酶(MAPK)信号通路能够调节细胞增殖、分化、细胞凋亡以及病理条件下的应激反应,可分为4 个亚族:细胞外信号调节激酶1/2(ERK1/2)、p38 丝裂原活化蛋白激酶(p38 MAPK)、c-Jun 氨基末端激酶(JNK)和细胞外信号调节激酶5(ERK5)。研究表明,MAPK 信号通路直接或间接参与巨核细胞的增殖、分化和成熟过程[15-17];ITP 诊治指南推荐的二线治疗药物艾曲波帕可以通过MAPK信号通路中ERK1/2 信号分子的活化促进巨核细胞分化成熟,加速生成血小板[18-19]。本研究结果显示,空白对照组及PMA 诱导组Dami 细胞的p-JNK、p-c-Jun 蛋白基本不表达,而APS 诱导后模型组Dami 细胞JNK、c-Jun 蛋白磷酸化水平显著升高。表明APS能够促进巨核细胞中JNK 及c-Jun 蛋白磷酸化,激活MAPK 信号通路,抑制巨核细胞分化成熟。而肿节风总黄酮干预后,APS 模型组细胞的p-JNK、pc-Jun 蛋白表达明显下调,提示肿节风总黄酮可能通过调节MAPK/JNK 信号通路来改善巨核细胞分化成熟障碍状况。
骨髓内巨核细胞分化成熟形成血小板是一个复杂的动态过程[20],MAPKs 在该过程中的调节作用亦复杂,且取决于多个因素,因为巨核细胞分化成熟过程本身涉及不同的连续步骤[21]。因此,本课题组后续将利用MAPK/JNK 信号通路激动剂、拮抗剂等进行对照实验,以更深入探讨肿节风总黄酮促巨核细胞分化成熟的机制。另外,MAPK 信号通路调控巨核细胞分化成熟的关键靶点也需要进一步验证,其上下游复杂信号转导通路的分子机制还有待更深入的研究。
猜你喜欢
清华金融评论(2022年4期)2022-04-13
服饰导报·鞋世界(2021年4期)2021-05-17
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
中国药理学通报(2019年5期)2019-01-11
中成药(2017年8期)2017-11-22
中成药(2017年10期)2017-11-16
中成药(2017年4期)2017-05-17
湖南师范大学自然科学学报(2015年1期)2015-02-27
中国药理学通报(2014年2期)2014-05-09
