
亚热带丘陵区人工湿地底栖动物群落结构动态特征与水环境因子关系
2023-07-04 06:33王浩李希孟岑徐英华谢陈刘铭羽李裕元吴金水
农业现代化研究 2023年2期
王浩 ,李希,孟岑,徐英华 ,谢陈,刘铭羽 ,李裕元 ,吴金水
(1. 中国科学院亚热带农业生态研究所,亚热带农业生态过程重点实验室,湖南 长沙 410125;2. 中国科学院大学,北京 100049;3. 吉首大学生物资源与环境科学学院,湖南 吉首 416000)
随着工业等点源污染不断得到治理,我国农业面源污染对环境污染的贡献日渐突出。由于农业面源污染存在量大、面广和随机性强的突出特点[1-2],因此很难采用集中的工程治理方法解决问题,人工湿地在治理农业面源污染方面具有得天独厚的优势。底栖动物作为湿地生态系统的重要组成部分,在污染物去除与能量转化中发挥着重要作用[3],因此探索人工湿地底栖动物群落结构及其与环境因子间的相互关系,为人工湿地生态功能的进一步提升提供理论依据具有重要意义。
人工湿地具有建造成本及运行费用低、易操作、水质净化效果好等明显优势,尤其对水体中氮磷等污染物的去除效果十分显著[4-5]。人工湿地对污染物的去除途径主要是通过底泥吸附、植物吸收、微生物作用等物理、化学和生物协同作用而实现[6-8],因此,目前大量学者对人工湿地植物、底泥和微生物的净化作用开展了较为系统、深入的研究[9-11],并取得重要进展。但是随着研究的不断深入,有研究发现,事实上底栖动物在湿地生态系统的物质代谢与能量传递中也发挥着重要作用[12-13],但针对人工湿地此方面的研究尚处于起步阶段,还未得到研究者的充分关注和重视。
底栖动物是表面流人工湿地生态系统的重要组成部分[14],是维持湿地健康运行的重要因素[15]。一方面,底栖动物可通过摄食、爬行、掘穴等生物扰动作用影响底泥(沉积物)的理化性质,促进沉积物中的氮磷释放,进而影响微生物活性,尤其是在不同程度上强化底泥的硝化—反硝化作用,从而提升湿地系统对氮的去除能力[16-17]。另一方面,底栖动物作为湿地生态系统食物网中的初级消费者,是链接生产者(植物)和分解者(微生物)的重要环节,在维持湿地生态系统物质循环和能量流动过程中发挥着重要作用,底栖动物的存在对于完善人工湿地生态功能具有重要意义[18-19]。此外,湿地在运行过程中,管理者还可通过对部分大型底栖动物(如螺蛳)适时适量的采收,从而带来一定的经济收益,从而实现生态效益与经济效益的有机结合。
目前针对人工湿地底栖动物群落特征与生态功能的研究还相对较少,已有研究主要是对河流湖泊等大型天然水体底栖动物相关功能的研究[20-21],而对人工湿地中底栖动物及其功能的研究还相对较为薄弱。鉴于此,本研究以湖南长沙亚热带丘陵区为研究区域,通过野外表面流人工湿地小区连续4 年控制试验,定位观测浮水植物、挺水植物和无植物浅水表面流人工湿地的水质处理效果和湿地底栖动物的群落特征及其变化,探讨其与水环境影响因子的关系,以期为人工湿地生态功能的进一步提升提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于湖南省长沙县开慧镇(113°24′ E、28°58′ N),属典型亚热带湿润季风气候,年平均气温17.2 ℃,最低气温-6.7 ℃,最高气温40.1 ℃,多年平均降水量1 394.6 mm,降水主要集中于4—7 月份。区域内主要土壤类型为红壤和水稻土,主要土地利用类型为稻田、果茶园、菜地和次生天然林地等。当地有着悠久的养猪传统,部分村民以分散养殖模式养猪,存栏规模一般为几十头至两百头,且随着市场行情变化存栏规模波动较大,当地的养殖废水和农村生活污水一般经过沼气池处理后会作为有机肥加以利用,但也有少部分经过厌氧或沼气池简单处理后排入周边沟渠,因此分散养殖会不同程度地影响当地的水环境。
人工湿地建造前为正常耕种的水稻田,建造时底泥未经任何扰动,0~20 cm 土壤基本理化性质为:pH 值6.1,有机质23.6 g/kg,全氮1.01 g/kg,全磷0.37 g/kg,土壤质地为粉砂壤土,其中土壤砂粒(>0.05 mm)和黏粒(<0.002 mm)含量分别为41.6%和10.0%。小区进水为附近的一条生态沟拦截的地表径流,上游区域包含3 个小型家庭养猪场(存栏规模为30~50 头)和25 户居民的生活污水排放,养殖场均有沼气池和资源化利用系统,各户的生活污水也均建有三级或四级化粪池。
1.2 试验设计
本试验开始于2019 年4 月。试验选取浮水植物和挺水植物两种典型湿地植物类型构建人工湿地,设置5 个试验处理:1 个浮水植物绿狐尾藻(Myriophyllum elatinoides)人工湿地;3 个挺水植物人工湿地,分别为黄菖蒲(Iris psudacorus)湿地、水生美人蕉(Canna glauca)湿地和梭鱼草(Pontederia cordata)湿地;1 个无植物对照(CK)。每个处理设3 个重复,所有处理湿地均分为三级,进水方式为连续进水,水力停留时间为30 d。
1.3 样品的采集与分析
1.3.1 水样采集与水环境理化指标测定 每10 天采集水样一次,于进水口和每级出水口取样,样品采集后带回实验室,混匀后分为两份,一份用于直接测定总氮(TN)、总磷(TP)与化学需氧量(COD),另一份经抽滤和离心处理后用于测定氨氮(NH3-N)和硝氮(NO3--N)。TN 采用碱性过硫酸钾消解—流动分析仪法测定;TP 采用过硫酸钾消解—流动分析仪法测定;COD 采用重铬酸钾消解—紫外分光光度法测定;氨氮和硝氮采用流动分析仪法直接测定。水温(WT)、酸碱度(pH)、溶解氧(DO)、电导率(EC)和氧化还原电位(Eh)等水体理化参数均使用多参数水质分析仪现场测定。
1.3.2 底栖动物样品采集与鉴定 本试验从2019 年4 月开始,最后一批试验数据时间为2022 年7 月,底栖动物样品于4 月(春季)、7 月(夏季)、10 月(秋季)、12 月或次年1 月(冬季)每个季节采集一次。每个植物处理设有三级,每级三个重复组,故每个小区选取9 个代表性采样点。用1/16 m2彼德森采泥器进行采集,泥样经60 目尼龙筛筛洗后倒入封口瓶,加入10%甲醛溶液后密封保存,带回实验室后参照有关资料,鉴定底栖动物到种,并计数和称重,然后计算出底栖动物单位面积的个体数量(即丰度,ind/m2)和生物量(g/m2)[22]。
1.4 底栖动物群落特征指标计算
本研究底栖动物优势物种根据优势度指数(YI)来确定,底栖动物的数量特征采用单位面积底栖动物的数量(丰度)和生物量来表示;湿地物种多样性指数用Shannon-Weiner 指数(SWI)表示,群落中物种丰富程度用Margalef 指数(MI)表示,物种个体在群落中分配的均匀程度用Pielou 指数(PLI)表示;通过分析偏最小二乘回归分析模型的PLS 回归系数可以判断作用方向和量化作用大小,影响变量在解释指标的重要程度时可以用重要性指标(VIP)来测度,一般认为,VIP 大于1.0 极其重要,值越大,该变量的解释能力越强。各主要指数的计算方法为:
式中:YI 为优势度指数,当Y> 0.02 时,确定该物种为优势种[23];Ai为相对丰度,即种i的个体数占总物种个体数的比例;Fi为物种i出现的频率,即出现种i的样点个数占总样点个数的比例;S为总的底栖动物物种数;N为所有底栖动物物种的个体总数。
1.5 数据的统计与分析
本文所有数据均采用SPSS 25.0 软件进行单因素方差分析(One-way ANOVA)和Spearman 相关分析,用Canoco 5.0 软件进行冗余分析(Redundancy Analysis,RDA),用SMICA 14.0 软件完成偏最小二乘回归分析(Partial Least Squares,PLS)。
2 结果与分析
2.1 底栖动物物种组成、优势度及其动态变化分析
连续近4 年(2019 年4 月至2022 年7 月)的观测结果表明,研究区人工湿地中共采集到底栖动物35 种,隶属于3 门4 纲8 目17 科,主要包括节肢动物28 种,分属于1 纲4 目12 科;环节动物4种,分属于2 纲2 目2 科;软体动物3 种,分属于1 纲2 目3 科。从物种数的逐年变化来看,各处理底栖动物的种类总体上呈逐年下降的变化趋势,其中2019 年为32 种,2022 年已经降低至14 种,除摇蚊以外的其他昆虫占比均下降显著。从不同年份物种组成占比来看,昆虫纲物种数量在群落中占比最高,变化范围为40%~90%(图1)。从具体构成物种来看,2019 年昆虫纲的物种组成主要以摇蚊科(Chironomidae)、蜻科(Libellulidae Rambur)为主,而2020 年后摇蚊科和蜻科有明显减少,仅有对照处理中蠓蚊(Culicoidessp.)有一定的优势度。
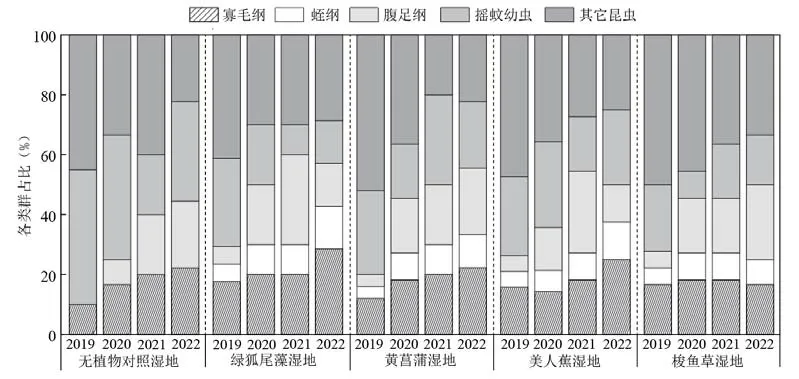
图1 不同类型人工湿地的底栖动物群落结构组成比较Fig. 1 Comparison of zoobenthos community structure in diff erent types of constructed wetlands
对各年份物种构成动态变化的分析结果表明,在不同季节与年际间各处理底栖动物的优势种组成基本一致,均为霍甫水丝蚓、苏氏尾鳃蚓(Branchiura sowerbyi)、黄色羽摇蚊(Chironomus f laνiplumus),其中霍甫水丝蚓在各个年份各处理的优势度值都远大于0.02,最高可达0.50(表1),为人工湿地中最主要的优势种。但是,在绿狐尾藻湿地中底栖动物群落结构有所不同,黄色羽摇蚊在春季的优势度明显低于其它处理(表2)。所以优势种的分布与季节、植物配置也有不同程度的关联性。总体而言,研究区人工湿地出现的底栖动物物种数明显偏少,主要以水生昆虫纲物种为主,缺乏大型底栖动物。
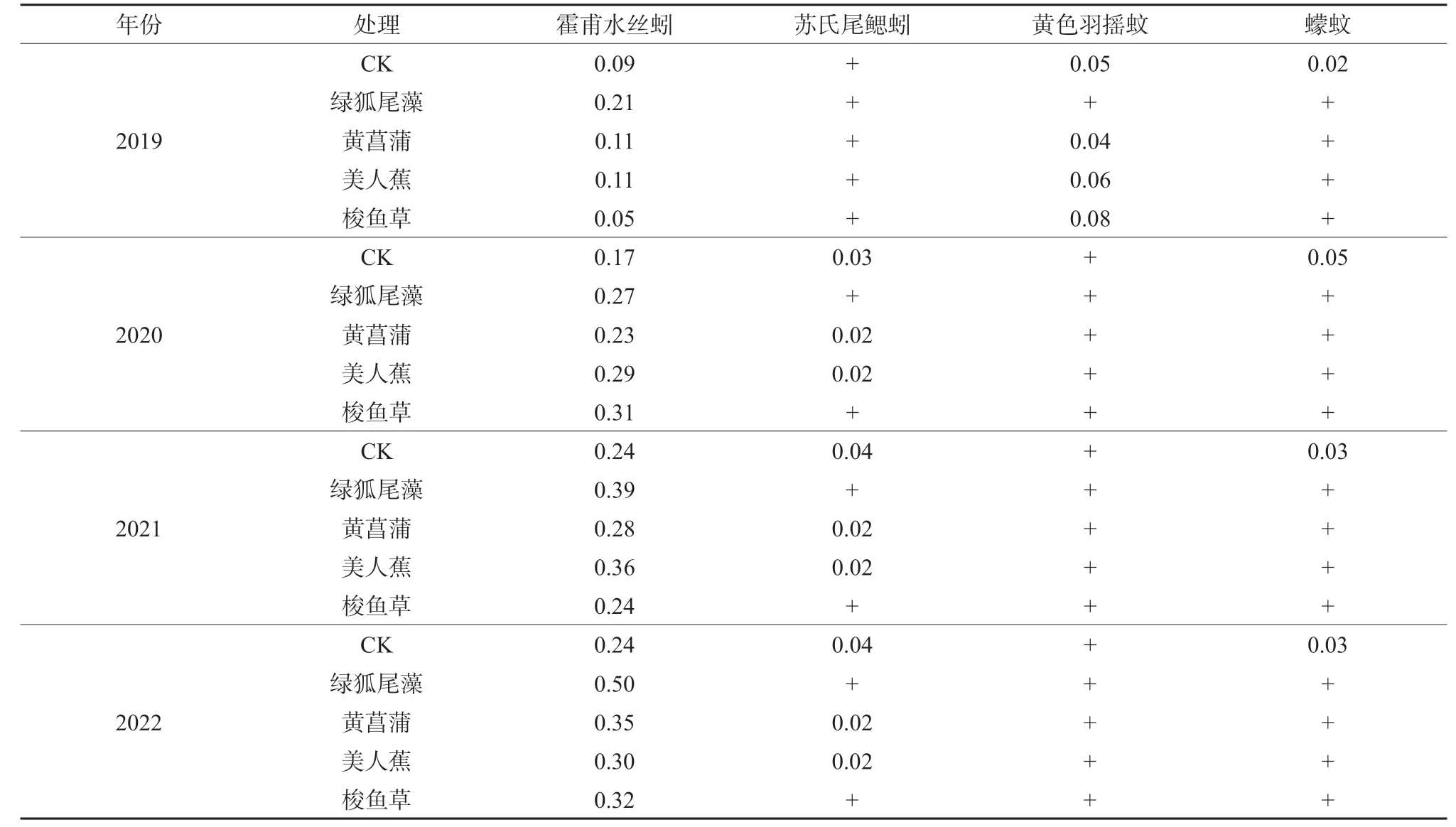
表1 人工湿地底栖动物优势种及优势度年际变化Table 1 Dominant species and interannual variations of zoobenthos in constructed wetlands
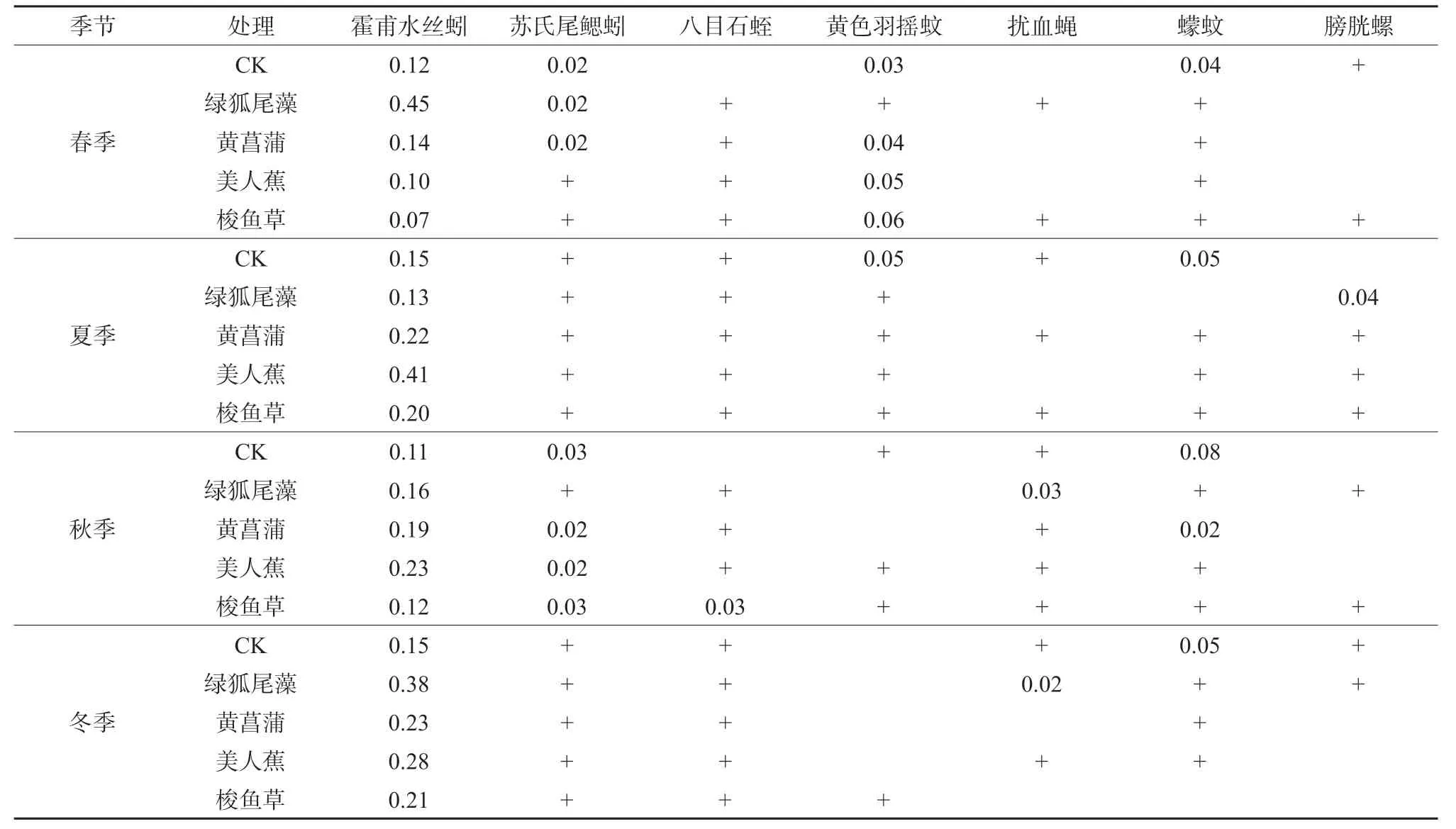
表2 人工湿地底栖动物优势种及优势度季节变化(2019—2022 年)Table 2 Dominant species and seasonal changes of dominance of zoobenthos in constructed wetlands (2019-2022)
2.2 物种丰度与生物量年际变化分析
底栖动物丰度结果显示,总体上,有植物配置的人工湿地其底栖动物丰度显著低于对照的无植物湿地(P<0.05),其中浮水植物绿狐尾藻湿地的底栖动物丰度为最低,其平均丰度仅为571 ind/m2,变化范围为85~1 079 ind/m2(图2);三种挺水植物黄菖蒲、美人蕉、梭鱼草湿地底栖动物丰度较为接近,4 年平均丰度分别为894、806 和740 ind/m2,统计差异不显著(P>0.05);而无植物对照的底栖动物丰度较高,平均丰度为1 532 ind/m2,与其他处理有显著差异(P<0.05)。从变化趋势上来看,5 个类型人工湿地中底栖动物丰度基本呈现出逐年下降的趋势,尤以三个挺水植物处理的变化更为明显,此外,绿狐尾藻人工湿地物种丰度的变化明显低于其它处理,变化幅度明显偏低。
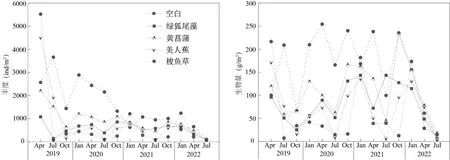
图2 人工湿地底栖动物丰度与生物量的动态变化Fig. 2 Dynamic changes of abundance and biomass of zoobenthos in constructed wetlands
底栖动物生物量结果显示,挺水植物黄菖蒲、美人蕉和浮水植物绿狐尾藻湿地的生物量总体上较为接近(图2),其中梭鱼草湿地底栖动物生物量最低,仅有61 g/m2,与无植物对照(158 g/m2)相比达到统计学差异水平(P<0.05)。从年际动态变化来看,由不同水生植物构建的湿地类型之间,虽然底栖动物生物量有一定差异,但变化趋势基本一致。总体而言,底栖动物丰度与生物量动态变化不尽一致,其中丰度基本呈现出明显的逐年下降趋势,而生物量则主要为季节波动,而降低趋势不明显。
2.3 底栖动物生物多样性变化分析
物种多样性的分析结果表明,多样性指数、均匀度指数、丰富度指数变化范围仅分别为0.59~1.25、0.34~0.60 和0.51~1.05(表3),但是同一年份处理间各指标的变化并不完全一致,2019 年有植物湿地多样性指数和丰富度指数的差异不明显(P> 0.05),而均匀度指数则表现为梭鱼草湿地显著高于黄菖蒲和美人蕉湿地(P< 0.05)。从不同年份的比较来看,黄菖蒲和美人蕉湿地中2019 年丰富度指数均显著高于2022 年(P< 0.05),而多样性指数则无显著差异(P> 0.05);黄菖蒲湿地中2022 年的均匀度指数显著高于2019 年(P< 0.05)。总体而言,人工湿地底栖动物生物多样性较低,波动范围不大。
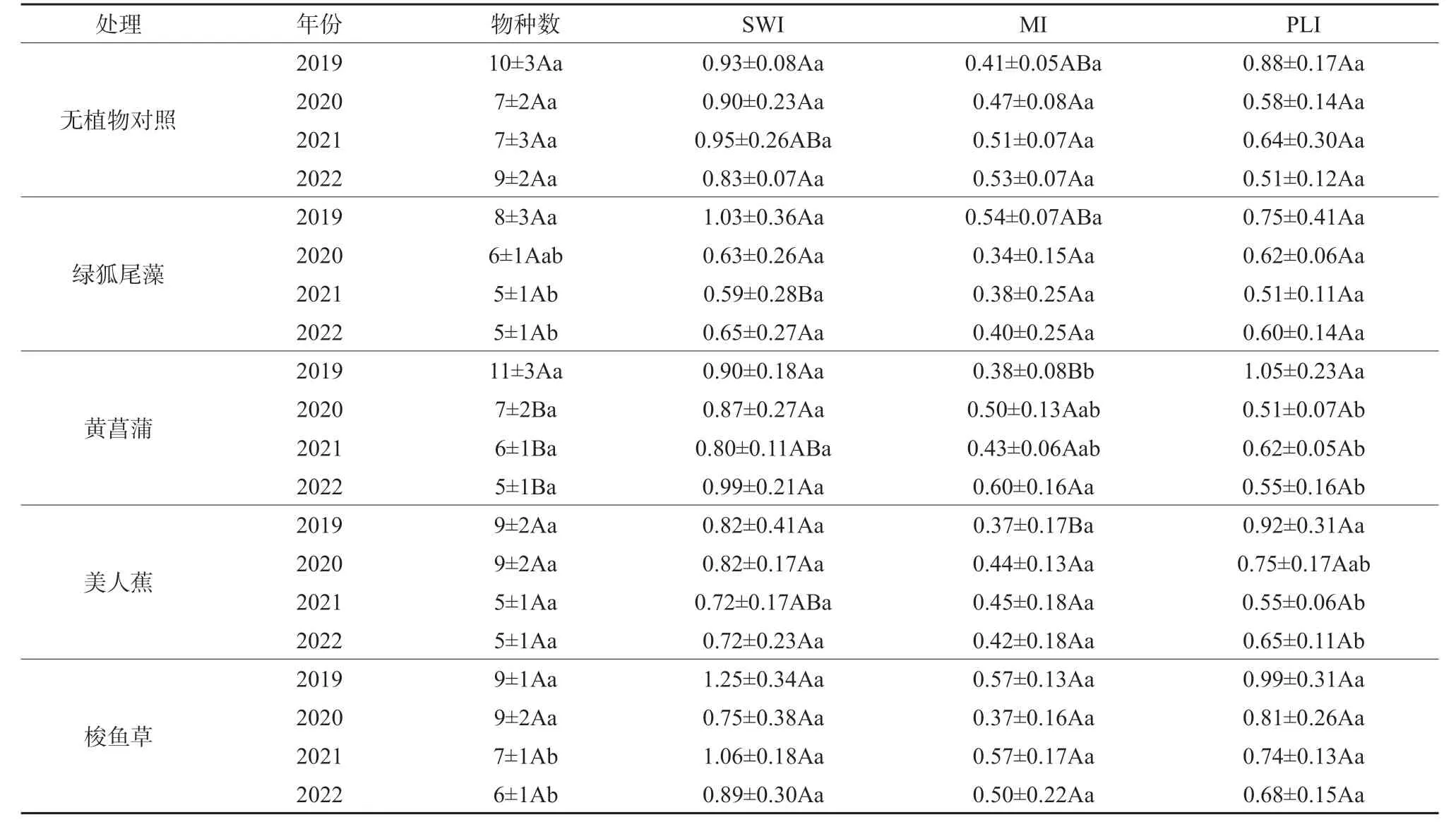
表3 人工湿地底栖动物生物多样性指数Table 3 Biodiversity index of zoobenthos in constructed wetlands
2.4 水环境因子年际变化分析
湿地主要污染物出水水质变化见表4。试验初期(2019 年)各植物处理湿地出水水质状况相对较差,出水均为地表劣V 类;由于受非洲猪瘟影响,2020 年以后区域内养殖规模大幅减小,相应地废水产生量也急剧下降,湿地进水主要以生活污水和农田排水为主,有植物的各湿地处理出水水质均有较大提升,出水TN、NH3-N 和COD 浓度呈现出明显的降低趋势,均达到地表IV 类水质标准(GB 3838—2002),TP 只有绿狐尾藻湿地可达到IV 类水质标准。总体而言,有植物配置湿地与无植物对照湿地处理效果差异显著(P< 0.05),湿地对污水的处理效果良好,绿狐尾藻湿地处理效果更加突出。
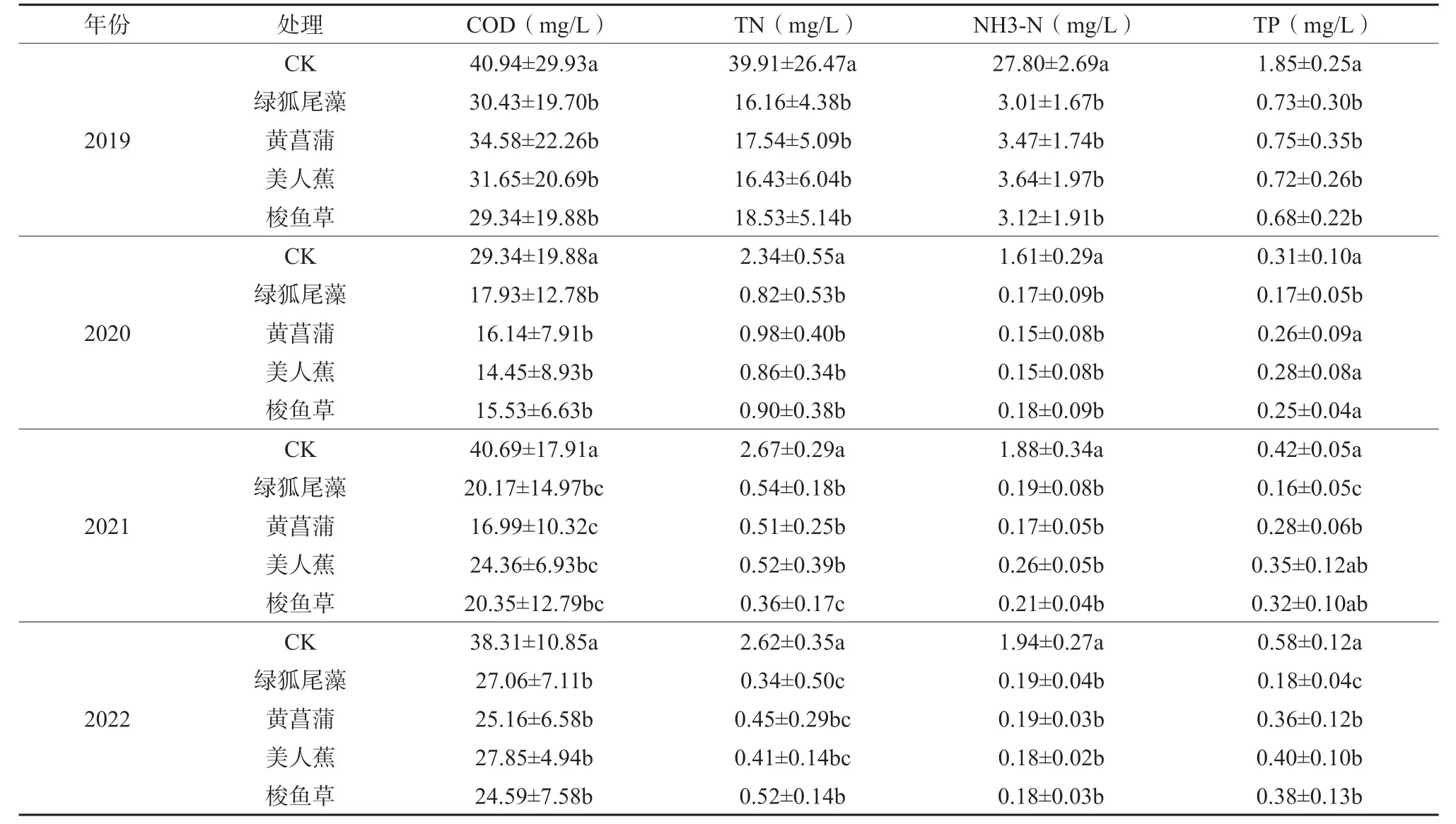
表4 人工湿地不同处理出水主要污染物浓度动态变化(2019—2022 年)Table 4 Dynamic changes in main pollutant concentrations in the effl uent of diff erent treatments in constructed wetlands(2019-2022)
湿地水体物理性质的观测结果表明,人工湿地温度变化范围为6.16~33.10 ℃,温度随季节变化但年际间差异不大;有植物配置湿地DO 显著高于无植物湿地(P< 0.05),浮水植物与挺水植物湿地间差距较小;有植物配置湿地pH 均处于7.0 左右,无植物对照湿地受到藻类影响,pH 呈弱碱性。Eh变化范围为-111.13~37.82 mV,而EC 在2020 年之后则显著下降,但后续波动不大(图3)。
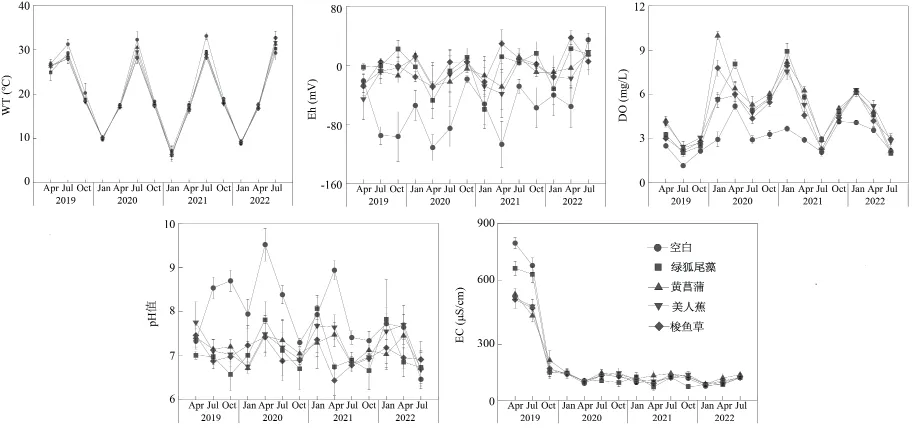
图3 人工湿地水体理化指标动态变化Fig. 3 Dynamic changes in water body physical and chemical indicators in constructed wetlands
2.5 底栖动物与环境因子的相关性分析
底栖动物群落指标与主要水环境因子的关联性分析结果表明,对于不同的底栖动物群落及多样性指标其所受的主导环境因素明显不同,其中,底栖动物物种丰度与种数基本一致,与NH3-N、TN、EC 呈显著正相关;生物量则与pH 呈显著正相关,而与Eh、WT 呈显著负相关(表5)。从不同的多样性指标来看,对丰富度指数和均匀度指数起关键影响的水环境因子均为pH、WT 和Eh,其中与pH呈负相关,与WT 和Eh 呈显著正相关。可见,湿地底栖动物丰度与物种数主要与氨氮与总氮关系更为密切,而区域群落特征指数则主要收到湿地水体物理指标影响。
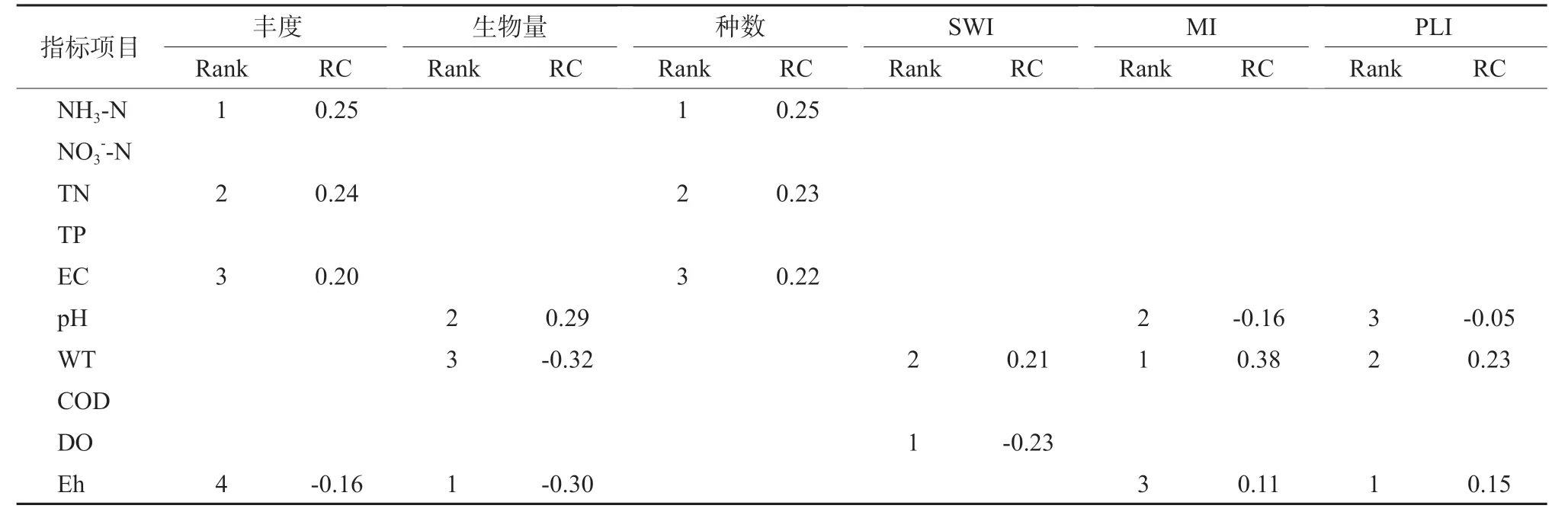
表5 对底栖动物群落指标具有关键影响的水环境因子的重要性排序和回归系数(VIP > 1)Table 5 The importance ranking and regression coeffi cient of water environmental factors inf luencing zoobenthos community indicators (VIP > 1)
底栖动物物种类群与环境因子的冗余分析结果表明,从化学指标来看,腹足纲与昆虫纲基本一致,与TP、COD 呈显著正相关,蛭纲则截然相反,呈显著负相关;寡毛纲主要与TN、NH3-N 呈显著正相关(图4)。从物理指标来看,所有类群均受到WT 和pH 的显著影响,其中腹足纲和除摇蚊以外的其它昆虫与WT、pH 呈正相关,摇蚊则只与WT呈显著正相关,寡毛纲与pH 呈显著正相关。可见,不同类群底栖动物所受的主导环境因子明显不同,因此底栖动物群落结构是对湿地水环境的综合反映。
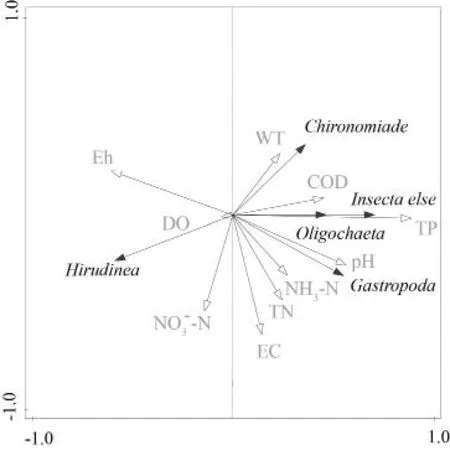
图4 底栖动物主要类别与湿地环境因子的RDA 分析Fig. 4 RDA analysis of main types of zoobenthos and wetland environmental factors
3 讨论
3.1 亚热带丘陵区人工湿地底栖动物群落构成的稳定性
亚热带丘陵区不同处理人工湿地底栖动物的动态观测结果表明,其物种种类呈现出逐年减少的趋势,丰度和生物量的变化趋势也基本一致。结合水质分析结果发现,底栖动物的动态趋势可能与湿地进水水质的好转有一定关系。随着湿地进水水质的向好,底栖动物的物种数量、丰度和生物量均表现出一致的下降趋势,表明底栖动物对水质变化的反映十分敏感。芦康乐等[24]在黄河三角洲湿地的研究表明,底栖动物因其对外界环境变化较为敏感,且迁移能力较差,进而其群落组成容易受自然和人为干扰的影响。秦珊等[25]对白洋淀底栖动物群落特征的研究表明,随着白洋淀水质的变好,其底栖动物群落的优势种从耐污种向清洁种和耐污种共存的方向转变,这与本研究人工湿地耐污种减少导致底栖动物种类减少,群落种类由以耐污种为主转向以清洁种为主的规律基本一致。其主要原因可能在于,在水环境整体处于较优质的情况下,耐污种无法获取足够的养分,再加之一些清洁种本身就适应更为干净的水体,在竞争后甚至可从耐污种类比例的降低中获利。这与王丑明等[26]对洞庭湖群落变化的研究结果基本相似,即群落呈单一化演变趋势,最主要的特征体现在湿地系统对敏感种的丧失和耐污种群的衰退,进而导致底栖动物物种群落结构出现简单化的变化趋势。因此,亚热带丘陵区人工湿地底栖动物的群落构成与水质条件具有密切关系,呈现出随环境变化的非稳定性特征。
不同类型人工湿地底栖动物优势种的分析结果表明,霍甫水丝蚓为典型的广布性优势种,在不同类型的湿地中均占居绝对的优势地位。有研究表明,霍甫水丝蚓为耐污性较强的物种,是水体污染的主要指示物种,其丰度的高低在一定程度上与水体的污染程度有正相关关系[27-28]。但是在不同年份和不同植物配置条件下优势物种组成有所差异,本试验人工湿地与大型流域、湖泊相比,底栖动物生物多样性总体上较低,体现在优势种上则是螺类较少,以摇蚊类底栖动物为主。2019 年黄色羽摇蚊也是在除绿狐尾藻湿地外其他植物湿地的优势种,且黄色羽摇蚊丰度显著高于霍甫水丝蚓,蠓蚊在各个年份的无植物对照湿地中均为主要优势种,表明湿地植物的郁闭度或去污能力对底栖动物分布会产生较大影响。进一步从连续几年的水质变化与物种组成的比较来看(表1 和表4),只有无植被对照处理蠓蚊的优势度较高,反映出水面开阔度是影响蠓蚊分布的关键因素。而对于黄色羽摇蚊而言,主要与水质的变化关系更为密切,水质越差,黄色羽摇蚊优势度越高。表明不同的底栖动物,其主导影响因素也并不完全一致,这主要归因于底栖动物自身的生物学特性。可见,尽管底栖动物群落构成呈现出随环境变化的非稳定性特征,但是以寡毛纲的霍甫水丝蚓和苏氏尾鳃蚓为主要优势种却相对稳定。
3.2 底栖动物群落特征与水环境因子的关系
底栖动物群落特征受水质和气候条件变化的双重影响,不同的群落构成指标受环境因子的影响显著不同,其中底栖动物丰度和物种数在一定程度上与NH3-N、TN 浓度呈显著正相关,物种丰富度和均匀度指数与水温呈显著正相关,而与pH 呈负相关,表明在一定程度上水体污染环境更有利于底栖动物的生长,同时底栖动物对于气候条件的变化也较为敏感。陆晓晗等[29]在山东省付疃河流域的研究结果与本研究的发现基本一致,并认为主要原因在于高浓度营养盐进入水体,加重了藻类的滋生蔓延,从而为耐污性底栖动物提供了足量的食物来源,给予其繁殖增生条件。有研究表明,水温在很大程度上影响底栖动物的密度与分布,当食物充足时,水温降低会抑制大型底栖动物的呼吸和代谢效率,使得底栖动物丰度减少,从而导致了丰富度指数和均匀度指数的降低[30-31];pH 值对底栖动物群落的繁殖能力影响较大,会导致底栖无脊椎动物多样性的显著降低[32]。
本研究发现,不同类群的底栖动物受环境因子的影响显著不同,寡毛纲与TN、NH3-N 呈显著正相关,意味着寡毛纲能够忍受一定程度的富营养化环境。这与张又等[33]对巢湖流域不同水系大型底栖动物群落的研究结果基本一致,在富营养化较为严重的下游河流中,寡毛纲的丰度和生物量较大,表明其对水体污染状况具有较强的指示意义。腹足纲与昆虫纲的变化趋势基本一致,均与TP、COD呈显著正相关,这与刘祥等[34]在江苏省淮河流域的研究略有不同,认为腹足纲对污染较为敏感,主要分布在水质较好的支流中,与固体悬浮物呈负相关,而昆虫纲则表现出较强的水环境适应性,与TN、COD 呈正相关。表明不同类群的底栖动物与环境因子的关系较为复杂,会受到多种因素的综合影响,具体影响机理仍需要进一步深入研究。由于底栖动物全部生活在水面以下,因此各个类群均受到水温和pH 的显著影响,其中全变态昆虫摇蚊受水温的影响最大,其生活史有明显的季节变化规律,在冬春季节以摇蚊为主的水生昆虫主要以幼虫形态存在于人工湿地水体或底泥之中,尤其是春季气温回暖,大量孵化滋生,导致种群丰度迅速升高,而夏秋季节则会大量羽化为成虫,离开水体,从而导致冬、春季节底栖动物总丰度显著高于夏、秋季的现象。因此,底栖动物群落结构特征是水质条件与气候环境的综合体现。
3.3 底栖动物对强化人工湿地功能的作用
底栖动物作为湿地的重要构成要素,在水体、水—泥界面、沉积物等介质中进行扰动、摄食、排泄等生物活动,增加溶解氧在沉积物中的渗透深度,使上覆水中有机质进入沉积物,在不同程度上加速氮、磷等主要污染物在沉积物与上覆水之间的交换速率,从而改变污染物在湿地系统中的转化途径。有研究表明,在人工湿地中加入底栖动物水丝蚓,水丝蚓通过爬行和掘食行为在沉积物中产生过水通路,改变沉积物孔隙度、理化性质,显著增强湿地的污染物去除能力[35]。同时,有研究表明,底栖动物会显著改变底泥微生物的群落结构,如泥鳅(Isgurnus anguillicaudatus)会显著降低底泥中的反硝化菌丰度,但能增加Beta 变形菌的数量,而田螺能显著增强底泥中微生物硝化反硝化作用,使得湿地系统对TN 和NO3--N 平均去除率提升22.98%和12.46%[36]。Carpintero 等[37]研究发现,在河口地区含有摇蚊幼虫和蜾蠃蜚(Corophium insidiosum)类底栖动物的沉积物中,其氮的去除能力均保持较高水平,表明底栖动物的生态特征和特定地点的沉积物特征对氮相关微生物群落和沉积物氮循环的调节机制有着不同的影响。因此,对人工湿地添加底栖动物,尤其是容易收集和投放的螺类、蚌类等大型底栖动物,对于强化人工湿地污染去除能力具有一定作用,也是提升人工湿地经济效益的重要途径,值得进一步深入探索。
4 结论
1)亚热带丘陵区人工湿地底栖动物群落结构相对较为简单,试验期内共采集到底栖动物36 种,其中霍甫水丝蚓为不同类型湿地的强优势种,优势度最高可达0.5,而底栖动物平均丰度和生物量相对较低,分别仅为909 ind/m2和96 g/m2。
2)底栖动物群落特征受水质和气候条件变化的双重影响,呈现出随环境变化的非稳定性特征,其中丰度和物种数在一定程度上与NH3-N、TN 浓度呈显著正相关,物种丰富度和均匀度指数与水温呈显著正相关,而与pH 呈负相关。但是不同类群所受的主导环境因子明显不同,寡毛纲与TN、NH3-N 呈显著正相关,而腹足纲和昆虫纲则主要与TP、COD 呈显著正相关。
3)底栖动物作为人工湿地的重要构成要素,对于强化人工湿地对污染物的消纳转化功能具有重要生态作用,通过添加底栖动物提升人工湿地的综合效益是一个重要途径,值得深入探索。
致谢:从2019 年试验开展以来,本所周训军老师在物种鉴定方面给予了全面、悉心的指导,课题组前期毕业的研究生夏梦华、赵聪芳、陈坤、郭宁宁、蒋磊、叶磊、凡翔、彭健、邓越以及在读研究生李情、白姣杰、贾中政等同学也参与了试验的大量工作,在此谨表示衷心感谢!
猜你喜欢
云南畜牧兽医(2021年1期)2021-02-24
今日农业(2020年19期)2020-12-14
科学大众(中学)(2019年3期)2019-05-17
意林·少年版(2019年1期)2019-01-16
汽车观察(2018年10期)2018-11-06
科技知识动漫(2017年1期)2017-02-06
天津师范大学学报(自然科学版)(2016年6期)2016-12-14
中国科技信息(2015年2期)2015-11-16
少儿科学周刊·少年版(2015年1期)2015-07-07
植物营养与肥料学报(2014年1期)2014-03-11
