
不同利用方式对天山北坡中段山地草甸草地植物生物量分布的影响
2023-05-30 13:21夏停停苏比努尔吾麦尔江于昭文吕文钧吐尔逊娜依热依木
新疆农业科学 2023年4期
夏停停,苏比努尔·吾麦尔江,于昭文,李 宏,吕文钧,吐尔逊娜依·热依木
(1.新疆农业大学草业学院,乌鲁木齐 830052;2.新疆农业大学生命科学学院,乌鲁木齐 830052)
0 引 言
【研究意义】天山北坡中段山地草甸草地牧草种类丰富[1],形成了供春、夏、秋、冬四个季节放牧利用的草地类型[2]。山地草甸草地是天山北坡中段典型的草地类型之一,气候湿润,雨量充沛,土层深厚且富含有机质肥力高的优势[3]。目前逐渐向冬季舍饲、春夏秋季放牧方式转变[4-5]。重度放牧使植物高度、丰富度、地上和地下生物量下降,导致草地退化[6]。草地植物生物量是体现草地生态系统功能的重要参数[7]。研究游牧和定居放牧变化对草地植物生产力的影响,对阐释不同放牧方式对山地草甸草地生产力的作用具有重要意义。【前人研究进展】有对青藏高原高寒草地[8],川西北高寒草甸[9],内蒙古草甸草原[10]等地区进行放牧对草地生物量的影响的研究,研究均表明重度放牧导致植物地上地下生物量显著减少;而轻度放牧可以增加草地群落生产力[11]。张景慧等[12]和高永恒等[13]研究发现,轻度放牧下草甸草原群落地上生物量减少,而地下生物量显著增加。崔淑娟等[14]研究表明,在适度放牧下,冷季放牧比暖季放牧更能促进草地植物地上生物量的积累。沈禹颖等[15]研究表明,地下生物量与地上生物量的比值(R/S)反映了光合作用物质在植物体内的分布。McNaughton[16]认为适度放牧会通过补偿生长机制增大草地的生物量对草原生态系统产生积极的作用。【本研究切入点】前人关于天山北坡山地草甸草地放牧与生物量关系的研究,主要集中在传统放牧、禁牧、封育等,缺乏对游牧与定居放牧下草地生物量分配的研究。需研究游牧与定居放牧下草地地上和地下生物量的变化规律。【拟解决的关键问题】以天山北坡中段山地草甸草地冬季牧场为对象,研究游牧放牧与定居放牧分别在LG和HG下的地上、地下生物量及植物根冠比的变化,为草地畜牧业生产提供理论依据。
1 材料与方法
1.1 材 料
研究区位于天山北坡中段昌吉市阿什里哈萨克民族乡阿什里村的冬季牧场,E 84°65'~87°78',N 43°74'~44°37',海拔为1 900~2 200 m,属中温带干旱大陆性气候,降水量夏季多冬季较少,年均降水量400~600 mm,年均气温为6.8℃。土壤类型为山地黑钙土。草地类型为山地草甸草地,草地植物群落优势种为细果苔草(Carexstenocarpa)、溚草(Koeleriacristata)等,由于过度放牧,致使毒害草醉马草(Achnatheruminebrians)在局部成为群落优势种;伴生种有委陵菜(Potentillasp.)、针茅(Stipacapillata)、黄芪(Astragalussp.)、草原糙苏(Phlomispratensis)、野韭菜(Alliumsp.)。
游牧放牧的转场时间:春季4月中旬至5月上旬从冬牧场转场至春场放牧;6月上旬转场到冬季牧场剪毛,6月中旬转到夏牧场放牧;9月上旬至10月中旬转场至秋牧场,11月上旬至次年4月中旬转场到冬牧场连续放牧6个月左右。定居放牧的转场时间:春季4月中旬至5月下旬从定居点转场至春场放牧,6月中旬转场到夏牧场,9月上旬至10月上旬转场至秋牧场,10月下旬至12月上旬冬季牧场放牧;12月中下旬至次年4月上旬在定居点舍饲圈养。游牧放牧与定居放牧主要区别是冬季牧场利用方式不同,定居牧户冬季牧场不放牧或放牧时间短(1~2月);其余牧场放牧时间、地点、草地面积基本与游牧牧户相同。
1.2 方 法
1.2.1 试验设计
在研究区内选择典型的山地草甸草地,设置2个试验区分别是游牧放牧区和定居放牧区。选择地形地貌、草地类型以及家畜种类、数量相近的游牧牧户(N)和定居牧户(S)各3户,分别标记N1、N2、N3和S1、S2、S3。将草地植物群落功能群分4类:多年生禾草(Perennial grasses)、多年生豆科草(Perennial legumes)、多年生莎草(Perennial sedges)、杂类草(Forbs)[17]。放牧强度依据植物群落组成变化、群落盖度和地上生物量的变化分别设置为2个放牧强度:轻度放牧(LG)、重度放牧(HG),围栏打草场作为对照(CK)。表1,表2

表1 冬季牧场基本信息
游牧牧户在中山带有夏季和冬季牧场,冬季在冬季牧场放牧+早晚补饲(饲料、干草);定居牧户在中山带有夏季和冬季牧场,定居牧户已经在平原区实现了冬季舍饲;在春秋季和夏季转场到春秋季和夏季牧场。游牧牧户冬季牧场放牧时间为2019年10月下旬到2020年4月下旬(约180 d)。定居牧户放牧时间2019年10月下旬至12月上旬(约60 d)。放牧草场以外的围栏区域作为对照CK(无放牧,2015年围栏禁牧)。每年打草刈割时间为8月上旬至下旬(20 d),刈割次数为1年1次。
1.2.2 测定指标
野外采样时间为2020年8月15日~9月20日。在游牧区和定居区每个放牧处理下记录每个重复样方内出现的植物种类数量;采用刈割法获取植物地上部分生物量;采用土钻法在获取地上植物生物量的样方内进行地下生物量取样,每个样方取5次。根据1 m×1 m的样方内的对角线,利用直径4 cm根钻在各样方选择深度0~10 cm、10~20 cm、20~30 cm分层取样。按照植物功能群分类编号将采集的样品分别装入信封袋中,带回实验室对不同处理的植物地上部分和根系分别烘干称重处理即为植物地上生物量和地下生物量。计算每个样方地下生物量与地上生物量比值的平均值作为植物的根冠比。
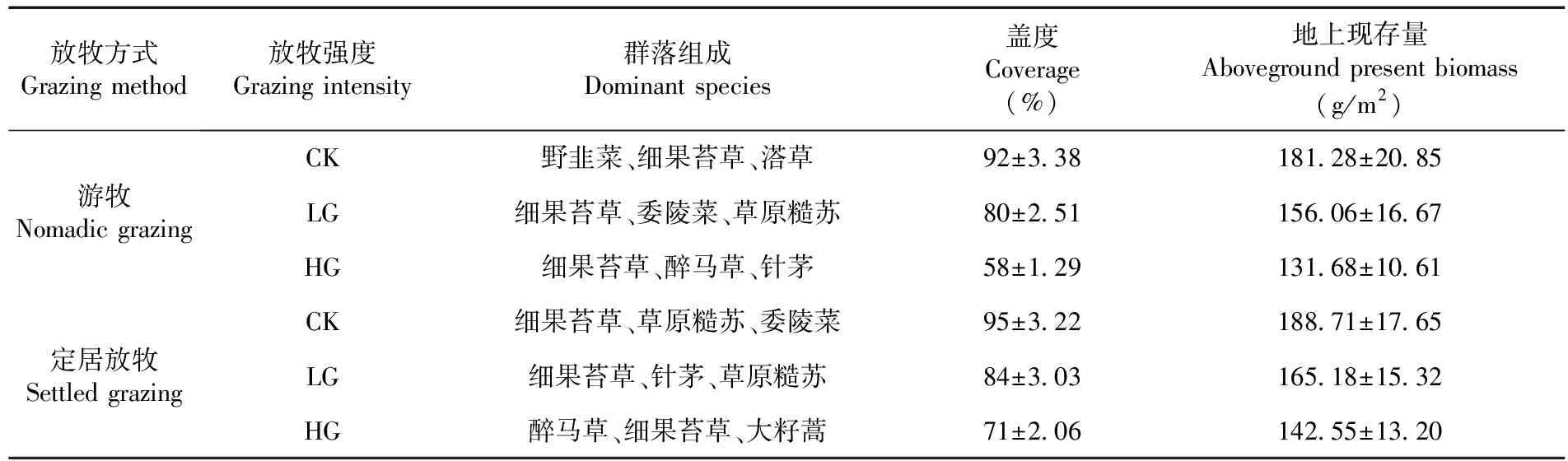
表2 不同放牧强度设置
1.3 数据处理
采用Microsoft Excel 2013对数据进行处理,SPASS 20.0统计软件对游牧与定居放牧下不同放牧强度下的植物地上生物量、地下生物量及根冠比进行One-Way ANOVA差异性检验,Origin 2019b绘制图表。
2 结果与分析
2.1 植物群落及功能群地上生物量的变化
研究表明,不同利用方式下多年生莎草和杂类草植物功能群地上生物量在LG下有显著差异(P<0.05),多年生豆科草植物地上生物量在HG下具有显著差异(P<0.05)。游牧放牧区内多年生禾草和多年生莎草功能群植物地上生物量在CK下高于定居放牧区;在LG、HG下,定居放牧区多年生禾草、多年生莎草、多年生豆科草植物功能群的地上生物量高于游牧放牧区。在LG下,定居放牧区多年生禾草、多年生豆科草植物功能群的地上生物量分别比游牧放牧区高出20.81%、1.52%,无显著差异。图1
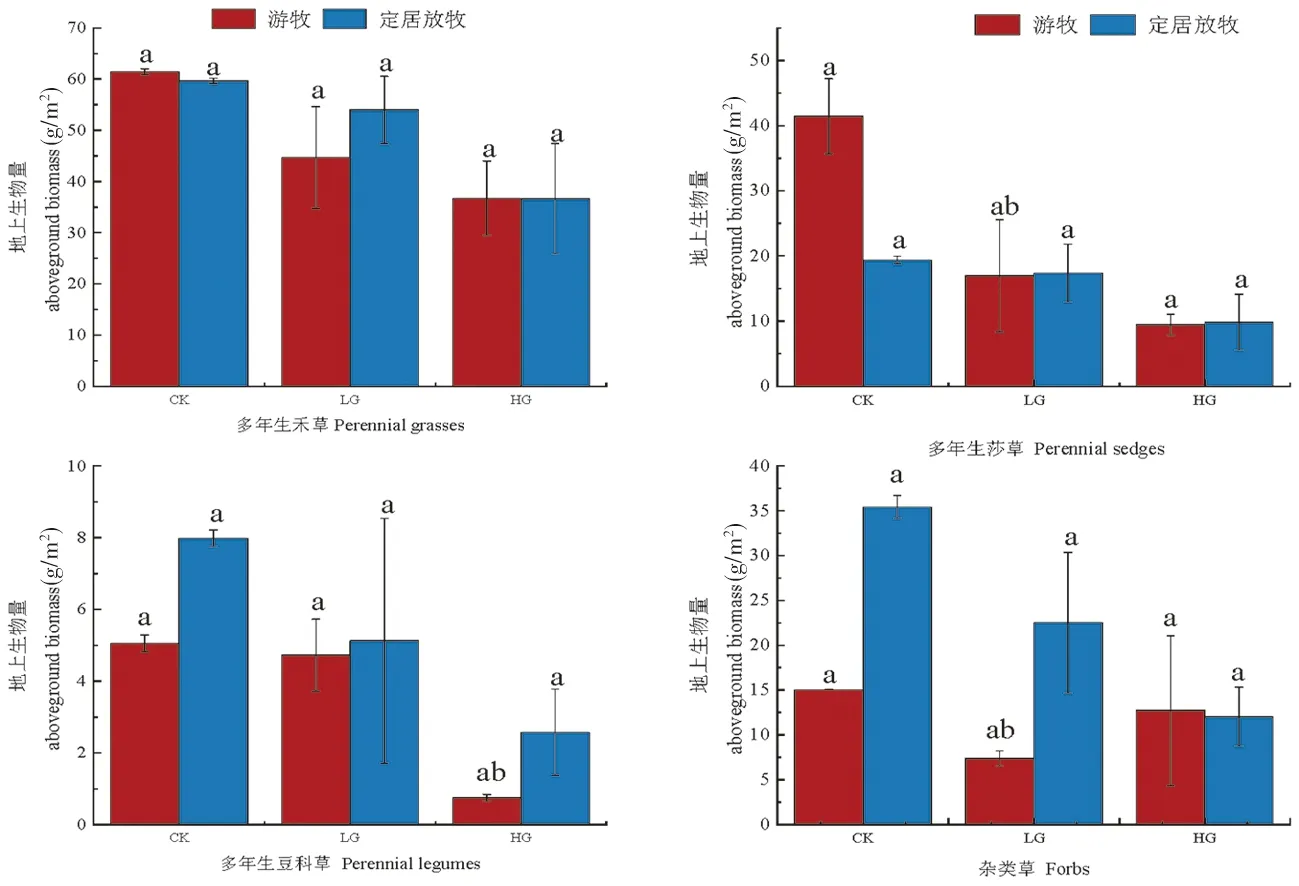
注:不同小写字母表示同一放牧方式不同放牧处理间的差异显著(P<0.05),下同
植物群落地上总生物量在不同放牧方式下有显著差异(P<0.05),且随放牧强度的增加均呈逐渐减少趋势,由大到小依次为CK>LG>HG,定居放牧区草地群落总生物量在LG和HG下分别比游牧放牧区高出10.72%、27.83%。图2

图2 植物群落地上总生物量变化
2.2 植物群落地下生物量的变化
研究表明,在不同放牧方式下,土层0~10、10~20、20~30 cm的植物地下生物量随放牧强度增加呈逐渐减少变化,随土层深度增加植物地下生物量呈逐渐减少变化。在CK、LG下,游牧放牧和定居放牧区内土层0~10、10~20、20~30 cm的植物地下生物量有显著差异(P<0.05),且定居放牧区植物地下生物量比游牧放牧区分别高出36.79%、31.55%、27.38%。在HG下,游牧和定居放牧区内土层20~30 cm的植物地下生物量无显著差异。在游牧放牧区内,土层0~30 cm的植物地下生物量范围在743.99~1 229.83 g/m2,土层0~10 cm占比最大为36.88%,土层10~20 cm占比最小为31.35%;在定居放牧区内,土层0~30 cm的植物地下生物量范围在864.33~1 550.95 g/m2,土层0~10 cm占比最大为37.93%,土层10~20 cm占比最小为28.73%。图3
2.3 植物群落地下地上生物量比值的变化
研究表明,不同放牧方式下的植物根冠比在LG、HG下有显著差异(P<0.05)。其中游牧放牧区植物根冠比由大到小依次为HG>CK>LG,比值分别为31.55、31.08、46.86;定居放牧区植物根冠比由大到小依次为HG>CK>LG,比值分别为41.81、37.10、51.51。在LG下,定居放牧区植物根冠比高游牧放牧区13.98%;在HG下,与游牧放牧区相比,定居放牧区植物根冠比高出9.91%。与CK相比,游牧放牧区植物根冠比在LG、HG下有显著差异(P<0.05),反之定居区植物根冠比无显著差异。随着放牧强度的增加,游牧和定居放牧区植物总生物量在地下分布先减少后增加,与CK、LG相比,HG下根冠比最大。图4

注:A:土层0~10 cm;B:土层10~20 cm;C:土层20~30 cm

图4 植物根冠比变化
3 讨 论
3.1 游牧和定居放牧下植物地上生物量变化
在一定时间内单位面积的草地植物群落积累产生的地上和地下生物量的总量即草地植物群落生产力,其影响因素包括不同放牧方式的和放牧强度[18]。草地植物生物量对草地生态系统功能具有重要的作用[19],其中地上生物量是决定天然草地生产力的主要因素之一。研究表明,草地群落地上生物量随放牧强度的增加而显著降低(P<0.05),放牧强度对地上生物量的影响是巨大的[20]。研究表明,游牧和定居放牧下随着放牧强度的增加,植物群落地上总生物量呈现逐渐减少趋势,即轻度放牧有利于刺激植物的生长[21]。在重度放牧下,由于家畜过度啃食,影响植物光合作用并导致植物不能正常生长发育,提供植物根系营养物质减少[22]。游牧区随放牧强度的增加,多年生禾草、多年生豆科草和多年生莎草植物功能群的地上生物量逐渐减少,由于多年生禾草植物适口性好,期间补偿生长不能够增加由于家畜采食失去的生物量[23],与段敏杰等[24]放牧对藏北紫花针茅高寒草原植物群落特征的影响研究结论一致,即随着放牧强度的增加,多年生禾草植物的生物量逐渐减少,逐渐被一些耐牧性较强的杂类草或毒害草取代。游牧与定居放牧区多年生禾草、多年生豆科草和多年生莎草的植物地上生物量与放牧强度呈负相关,在重度放牧下达到最小,由于定居区放牧时间较游牧区短,家畜啃食少,促进禾本科草类和莎草科草类植物生长,研究中游牧区植物地上生物量小于定居放牧区。反之,游牧区杂类草地上生物量在重度放牧下达到最大,原因是杂类草适口性差,耐践踏,放牧抑制其它适口性好的植物的生长,使杂类草能有更多的空间和养分资源,促进了物质能量的积累[25-26],故随放牧强度的增加,杂类草的地上生物量先减少后增加。
3.2 游牧和定居放牧下植物地下生物量变化
地下生物量是植物总生物量的组成部分,草地植物群落的主要生物量都分配于地下[27]。李怡等[22]研究表明,植物地下生物量分配情况为垂直分布且呈“T”型,不同土层深度的生物量由高到低逐渐递减。在不同放牧强度下,通过对植物光合产物调整其在地上、地下部分的分配规律,体现不同的适应性。研究表明,游牧和定居放牧区内,土层0~10、10~20、20~30 cm的植物地下生物量随着放牧强度的增加,植物地下生物量呈逐渐减少趋势,与王悦骅等[28]、宇万太等[29]研究结论一致,即草地植物地下生物量主要集中分布在浅层土壤中,随着土层深度逐渐增加植物地下生物量呈减少变化。轻度放牧下会导致植物根系浅表层化分布,重度放牧下植物根系呈深层化分布。
3.3 游牧和定居放牧下植物根冠比的变化
根冠比是植物生物量在地下与地上的分配比例,可以用来反映植物的生长策略[30]。随着植物根冠比增加,地上生物量逐渐减少导致出现草地退化现象[31]。Deng等[32]的研究表明植物的地下生物量对不同放牧强度响应不显著,两者呈负相关。在重度放牧下,家畜啃食大量适口植物导致顶端优势丧失,进而将更多的营养物质提供植物根系,使得地下生物量以及根冠比显著增加,为重度放牧后植物的继续生长提供所需营养物质和能量基础[33]。研究表明,随放牧强度增加植物根冠比呈现逐渐增加的趋势,由于家畜的采食和践踏使得地上部分不能充分生长,地上生物量减少的幅度常大于地下部分,造成根冠比随放牧强度的增大而增大。放牧虽然一定程度上激发了植物的补偿性生长,但并不能弥补家畜采食对植物光合能力造成的影响,地上生物量不能及时再生[34]。
4 结 论
4.1随着放牧强度增加,游牧和定居放牧区植物群落地上总生物量逐渐减少,有显著差异(P<0.05);定居放牧区多年生豆科草植物地上生物量在HG下显著高于游牧放牧(P<0.05),游牧和定居放牧区多年生莎草和杂类草在LG下有显著差异(P<0.05)。
4.2游牧和定居放牧区植物地下生物量随土层深度增加逐渐减少,由大到小依次为0~10 cm>10~20 cm>20~30 cm,且定居放牧区植物地下总生物量高于游牧放牧区。在CK、LG下,游牧和定居放牧区内土层0~10、10~20、20~30 cm的植物地下生物量有显著差异(P<0.05)。
4.3游牧和定居放牧区植物根冠比随放牧强度增加呈先减少后增加趋势,由大到小依次为HG>CK>LG。游牧放牧区植物根冠比在CK与LG、HG之间有显著差异(P<0.05),反之,定居放牧区植物根冠比没有显著差异。
猜你喜欢
ELLE世界时装之苑(2021年2期)2021-02-08
中国设备工程(2020年3期)2020-03-27
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
西夏学(2017年1期)2017-10-24
现代农业科技(2017年16期)2017-09-22
公民与法治(2016年1期)2016-05-17
丝绸之路(2015年16期)2015-12-26
Coco薇(2015年1期)2015-08-13
江苏农业科学(2014年1期)2014-07-18
