
黄精植株构件生长与有效成分积累动态研究
2023-05-29 03:26:24靳鹏博常晖杨祎辰王二欢王继强刘峰马存德
安徽农业科学 2023年9期
靳鹏博 常晖 杨祎辰 王二欢 王继强 刘峰 马存德
摘要 [目的]明確黄精植株构件的生长规律与地下部分各部位有效成分含量的积累动态。[方法]以2年生黄精为研究对象,黄精出苗后连续采样,测量根茎、新生根茎、须根、茎、叶、果实等构件的形态、数量和生物量,并测定地下部分各部位的黄精多糖、总皂苷和浸出物含量。[结果]黄精出苗后根系生物量不断积累,地上部分出苗后40 d内迅速生长,之后大多生长指标小幅度增加,至8月左右地上部分茎叶有所脱落,10月上旬地上部分枯萎回苗;地下部分在出苗40~100 d迅速生长,之后增长幅度减缓;须根的黄精多糖、总皂苷和浸出物含量均显著低于根茎(P<0.05),但收获期的须根多糖含量可达到根茎的84.50%,总皂苷和浸出物含量分别仅为根茎的35.72%、48.68%,这为黄精须根以多糖类材料开发大健康产品提供了依据;与原生根茎相比,新生根茎在形成的60 d内多糖含量显著较低(P<0.05),总皂苷含量显著较高(P<0.05);收获期时,黄精地下部分各部位以上3类有效成分含量排序稳定,依次为根茎>新根茎>须根。[结论]该研究为黄精非药用部位的开发利用提供了理论依据。
关键词 黄精;植株构件;生物量;多糖;总皂苷;浸出物;积累动态
中图分类号 S567.23 文献标识码 A 文章编号 0517-6611(2023)09-0155-05
doi:10.3969/j.issn.0517-6611.2023.09.038
Abstract [Objective]To clarify the growth law of plant structural biomass of Polygonatum sibiricum and the accumulation dynamics of active component content in various parts of its underground part.[Method]Taking twoyearold P. sibiricum as the research object and sampling continuously after emergence, the morphology, quantity and biomass of rhizomes, new rhizomes, fibrous roots, stems, leaves, fruits and other structural biomass were measured, and the contents of polysaccharide, total saponins and extracts in various parts of the underground part of P. sibiricum. [Result]The root biomass accumulated continuously after the emergence of P.sibiricum, the aboveground part grew rapidly within 40 days after the emergence, and the most growth indexes increased slightly. The aboveground part of the stem and leaf fell off by about August, and the aboveground part withered and returned to the seedling in early October.The underground part growed rapidly from 40 d to 100 d after emergence, and then the growth rate slowed down.The contents of polysaccharide, total saponin and extract of P.sibiricum in fibrous roots were significantly lower than those in rhizomes (P<0.05), but the content of polysaccharide in fibrous roots during harvest could reach 84.50% of rhizomes, the content of total saponins and of extract was 35.72% and 48.68%, which provided a basis for P.sibiricum fibrous roots to develop healthy products with polysaccharides.Compared with the original rhizome, the polysaccharide content of the new rhizome was significantly lower (P<0.05) and the total saponin content was significantly higher (P<0.05).During the harvest period, the contents of the above three kinds of effective components in each part of the underground part of P.sibiricum were in stable order: rhizome > new rhizome > fibrous root. [Conclusion] This study provides a theoretical basis for the development and utilization of non medicinal parts of P.sibiricum.
Key words Polygonatum sibiricum;Plant component;Biomass;Polysaccharide;Total saponins;Extract;Accumulation dynamic
基金项目 陕西省重点研发专项(2023-YBSF-414,2021SF-389);陕西省中医药管理局项目(2021-QYZL-01,2021-QYPT-001,SZY-KJCYC-2023-038);国家中医药管理局项目(财社〔2018〕43号,财社〔2019〕39号)。
作者简介 靳鹏博(1992—),男,陕西铜川人,主管药师,硕士,从事药用植物规范化栽培及药材质量标准研究。
*通信作者,主管药师,硕士,从事中药资源与生态种植研究。
植株是植物各个器官的总和,也可以说是其所有构件组成的统一整体,植物的生长发育和植株各个构件有着密切的关系[1]。在农业生产中,人们通过改变植株的一些构件的形态或数量,使目标器官形态或数量朝着人们期望的方向发展,甚至改变器官内部结构或一些成分的含量。李漓等[2]通过摘除花蕾提升了药材玄参的产量;张思唯等[3]研究发现打顶时期对雪茄烟叶的总氮、蛋白质和烟碱含量会有不同程度的影响,并且初花打顶可改善雪茄烟叶口感;赵国栋等[4]通过不同时期对苹果树进行拉枝,改变苹果树的构件结构,达到提升苹果产量、提升品质的效果。
黄精(Polygonatum sibiricum Red.)为百合科黄精属多年生草本植物,以根茎入药,习称“鸡头黄精”,分布于我国北方多省份,因其药食同源的特性,近年来已成为开发大健康产品的重要目标[5-6]。目前黄精生产上还存在种植周期长、种植多为山区、引入机械化难度大、人工栽培根茎“鸡头”性状变异、加工炮制无相关标准、产量和品質仍需提升等问题,与之相关的研究也不断报道。该研究对黄精植株的构件生长和有效成分积累变化进行了研究,并阐述黄精植株各构件的生长特性,为黄精非药用部位的开发利用提供了理论依据,以期解决目前黄精种植生产过程中存在的问题。
1 材料与方法
1.1 试验材料
试验所用材料为黄精无性繁殖得到的种根茎,由陕西步长制药有限公司黄精GAP基地提供,经陕西国际商贸学院马存德副主任药师鉴定为百合科植物黄精(Polygonatum sibiricum Red.)。
1.2 仪器和试剂
DHG-9243A鼓风干燥箱(上海精其仪器有限公司),MNT-150游标卡尺(上海美耐特实业有限公司),OHAUS-AX124ZH电子天平[奥豪斯仪器(上海)有限公司],UV-2600i紫外可见分光光度计[岛津企业管理(中国)有限公司],乙醇(天津市大茂化学试剂厂),蒽酮(国药集团化学试剂有限公司),无水葡萄糖(国药集团化学试剂有限公司),硫酸(天津市大茂化学试剂厂),挂牌,米尺,蒸发皿。
1.3 试验地概况
试验地点为陕西步长制药有限公司黄精GAP基地,位于陕西省略阳县五龙洞镇中川坝村,试验地坐标为33°30′14.78″N、106°12′32.42″E,海拔954 m,年日照时数1 558.3 h,年降水量860 mm,年均气温13.2 ℃,年无霜期236 d。
2020年11月对试验地清地、耕地、施基肥、起垄,基肥为生物有机肥,施用量4 500 kg/hm2,垄宽1.2 m,垄间留0.6 m宽的过道;2020年11月,将种根茎以2 250 kg/hm2分3行播入垄中,覆土;2021年4月中旬,在过道内以株距40 cm套种1行玉米。试验地地势平坦,远离公路,外围设有1 m宽的保护行。
1.4 试验设计
2021年4月6日,试验田的黄精不定芽已经形成茎尖冲出土层,随机采挖20株茎尖还未展开的黄精,作为生育期记录的起点;4月26日,出苗20 d,试验田里的黄精叶片大部分已经展开,有的已经生出花蕾,随机采挖20株叶片已展开的黄精;5月16日,出苗40 d,试验田里的黄精已全部结果,此后每月中旬随机采挖20株具有当前长势代表的植株;出苗160 d为9月中旬,最后一次采集完整的黄精植株;10月上旬,黄精植株地上部分回苗,10月和11月仅采挖地下部分。
采挖的植株清洗除去泥土,晾干水分,测量每个植株的形态特征(根茎长度、根茎直径、茎长度、5 cm处地径、中位叶长、中位叶宽),将植株的各个构件进行拆分,计数器官数量(须根条数、叶片数、果实数),称量各构件鲜重(根茎、新根茎、须根、芽、茎、叶、果实),随后105 ℃杀青,60 ℃烘干至恒重,称量各构件的干重。烘干后的地下部分按构件进行混合,粉碎,过5号筛,装袋,用于含量测定。
1.5 含量测定
1.5.1 浸出物与黄精多糖含量测定。
按照2020年版《中国药典》一部黄精项下方法完成浸出物与黄精多糖的含量测定[7]。黄精多糖的标准曲线制作,以吸光度A582为纵坐标、加硫酸前的反应液浓度(mg/mL)为横坐标,绘制无水葡萄糖的标准曲线,得出线性回归方程为Y=7.683 1X-0.033 2(R2=0.999 9)。
1.5.2 总皂苷含量测定。
参考廖念[8]的研究方法,取0.5 g样品粉末于10 mL离心管中,加入8 mL 80%乙醇,使用铁丝搅匀,超声提取30 min,4 000 r/min 离心10 min,收集上清液于50 mL容量瓶,从加入8 mL 80%乙醇开始再重复4次,80%乙醇定容,摇匀,即为样品的总皂苷提取液。
取0.2 mL提取液于25 mL刻度试管,置于80 ℃水浴锅15 min挥干提取液中的乙醇,试管取出放凉后加入0.2 mL 5%香草醛-冰醋酸溶液,冰浴加入0.8 mL 高氯酸,摇匀,置于60 ℃水浴锅保温15 min,试管取出放凉后加入5 mL冰醋酸,摇匀。以80%乙醇代替提取液调零,记录各样品的A550。
标准曲线的绘制:取薯蓣皂苷元标准物质20 mg,溶解于80%乙醇,并定容至100 mL,即得0.2 mg/mL薯蓣皂苷元标准溶液;分别取0.1、0.2、0.3、0.4、0.5 mL标准溶液,按照测定方法代替提取液进行反应。以试管内的总皂苷质量(mg)为横坐标、吸光度A550为纵坐标绘制标准曲线,得到线性回归方程为Y=14.26X+0.015 8(R2=0.999 9)。
1.6 数据处理
记录的数据使用 Microsoft Excel 和 IBM SPSS Statistics 19 处理分析。
2 结果与分析
2.1 黄精植株构件的形态生长
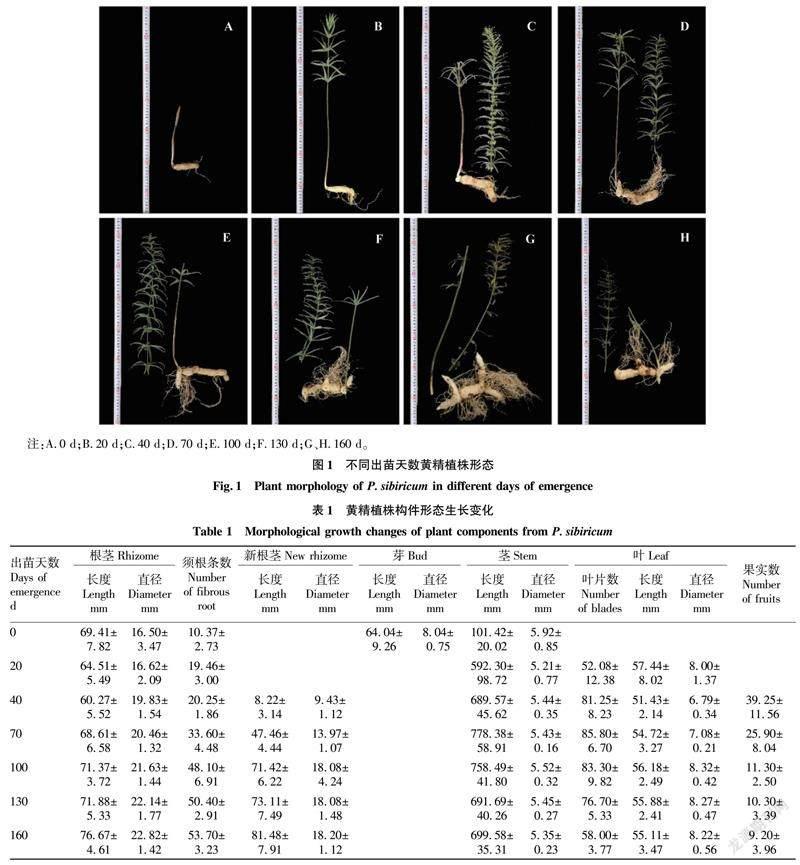
黄精植株构件形态生长变化和不同出苗天数植株形态分别见表1和图1。对不同时期黄精植株各器官形态或数量分析(表1)发现,随着生育期的推移,黄精根茎长度呈现先减少后增加的趋势,这是因为黄精在出苗后不久(出苗20 d前后),根茎最前端的不定芽和一部分根茎组织会生长和分化形成茎,随后(出苗40 d前后)在与茎连接的地方逐步形成新的根茎,茎和新根茎的形成均会导致老根茎长度折损,之后根茎长度持续增加,根茎直径在整个生育期内呈稳定持续增加的趋势;前一年冬季移栽至当年4月6日,黄精根部几乎不生长,之后须根条数持续增加,须根条数的增加为植株整个生育期的生长提供养分来源;黄精新根茎一般于花期前后(出苗20~40 d)形成,新根茎形成后至出苗100 d其长度和直径快速增加,之后增速变缓。
4月6日采挖的黄精出苗期植株带有一段茎,该段茎埋于土中,连接根茎与茎尖(芽),从芽和茎的形态数据(表1)来看,芽的直径大于茎,这是因为茎尖除了外围有2~3层鞘状的芽膜外,内部还包有正在生长的茎和未完全分化的花与叶;茎的长度和直径在出苗20 d至植株10月份回苗均表现为先增加后减少,出苗70和100 d分别为黄精植株叶片展开后茎的长度和直径最大值的时间点,此后茎长度和直径减少,是因为黄精植株在完成地上部分營养器官的完全展开后,顶部茎叶或受环境因素会发生不同程度的脱落,也有可能与营养生长转向生殖生长或地上部分营养转至地下部分储藏有关。
叶片数从出苗20 d至出苗70 d不断增加,之后植株叶片不同程度脱落,至10月份回苗地上部分全部枯萎,黄精植株叶片由底轮至顶轮表现为逐渐变小,出苗20 d黄精植株叶片仍有未完全展开的,此时中位叶的位置靠近底轮,因此中位叶的叶长叶宽数值较大,出苗40 d植株叶片完全展开,中位叶位置稳定,叶长叶宽此后不断增加,出苗100 d后,中位叶的叶长和叶宽以微小幅度减少。
黄精植株上的果实数自出苗40 d起不断减少,这是由于果实生长对植株养分消耗增加,加上环境因素如高温、降雨的干扰,植株上的果实不断脱落,以出苗40~100 d果实脱落最为明显,之后植株养分供应稳定,仅受环境因素干扰,果实脱落数较少。
2.2 黄精植株构件生物量积累
对不同时期黄精植株各器官的鲜重和干重分析(表2)可以发现,整个生育期内黄精根茎鲜重和干重总体呈上升趋势,以出苗70~100 d根茎的增加量最大,出苗130~160 d老根茎鲜重和干重有所下降,这可能是受气候原因,植株即将回苗,将生物量转移至新根茎上,利于新根茎不定芽的生长和次年出苗。黄精须根的鲜重和干重在整个生育期内呈持续上升趋势,出苗至100 d,须根的干/鲜比持续增加,之后至出苗130 d,须根的干/鲜比大幅降低,这可能是因为8月前后为当地的降雨主要时间,土壤含水较多,且新长出的根茎上此时有较多的须根生出,新生出的须根质地嫩,植株须根的总体水分含量较高,可能导致出苗130 d须根的干/鲜比骤降。新根茎形成是黄精生物量积累以及产量增加最主要的表现形式,表2显示自新根茎形成以后,新根茎鲜重的增加速度极快,且增长稳定。
茎是植株地上部分空间结构的骨架,出苗后茎迅速生长,出苗40 d茎的鲜重和干重分别已达整个生育期最大值的97.1%和92.4%,之后小幅增长或稳定,出苗130 d后茎的鲜重和干重均减少。叶鲜重和干重的变化均呈现先增后减的趋势,叶片形成后至出苗100 d,叶片数明显增多,叶片的长宽和厚度有所增加,使叶生物量稳定增长,出苗130~160 d,叶片脱落导致叶片数减少,叶长叶宽微微减小,叶的干重降低,鲜重在出苗130 d后降低。果实自形成后,数量一直在减少,果实的生物量却呈现先增加后减少的变化趋势,这是因为出苗40 d黄精刚进入果期,果实不断生长发育,至出苗70 d虽有约1/3的果实脱落,但单个果实生物量大幅度增加,果实总生物量呈上升趋势,出苗70~100 d为黄精种子开始形成和发育的阶段,果实内种子生物量占比增加,果肉量相对减少,单个果实的生物量小幅度增加,此间约有50%的果实脱落,果实总生物量下降,出苗100 d后果实小幅度脱落,果实内种子逐渐发育,果实的干/鲜比持续增加(表2)。
2.3 黄精地下部分有效成分变化
2.3.1 各部位多糖含量变化。
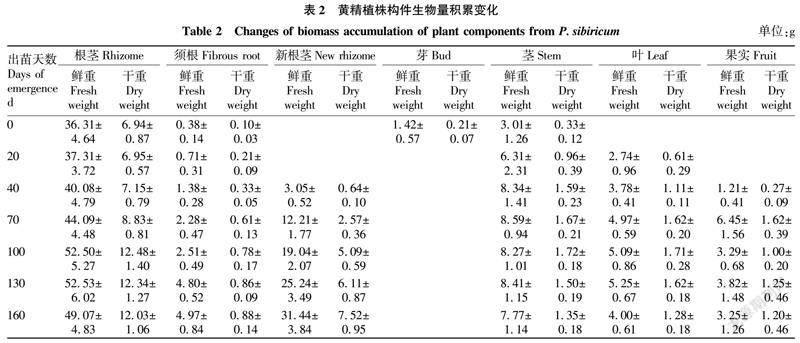
从不同时期黄精植株地下各部位的多糖含量变化(图2)可以看出,整个生育期内根茎部位的多糖含量表现为出苗70~160 d黄精植株地上部分形成并正常生长的时期多糖含量较低,出苗70 d之前和160 d之后多糖含量较高。新根茎自形成以后,其多糖含量持续增加,出苗100 d后与根茎的多糖含量无显著差异(P>0.05);须根的多糖含量变化趋势与根茎的一致,但其多糖含量显著低于根茎(P<0.05)。以10月中旬为采收期(出苗190 d)采挖的黄精,其地下各部位的多糖含量表现为根茎>新根茎>须根,须根多糖含量可达到根茎的84.50%。
2.3.2 各部位总皂苷含量变化。
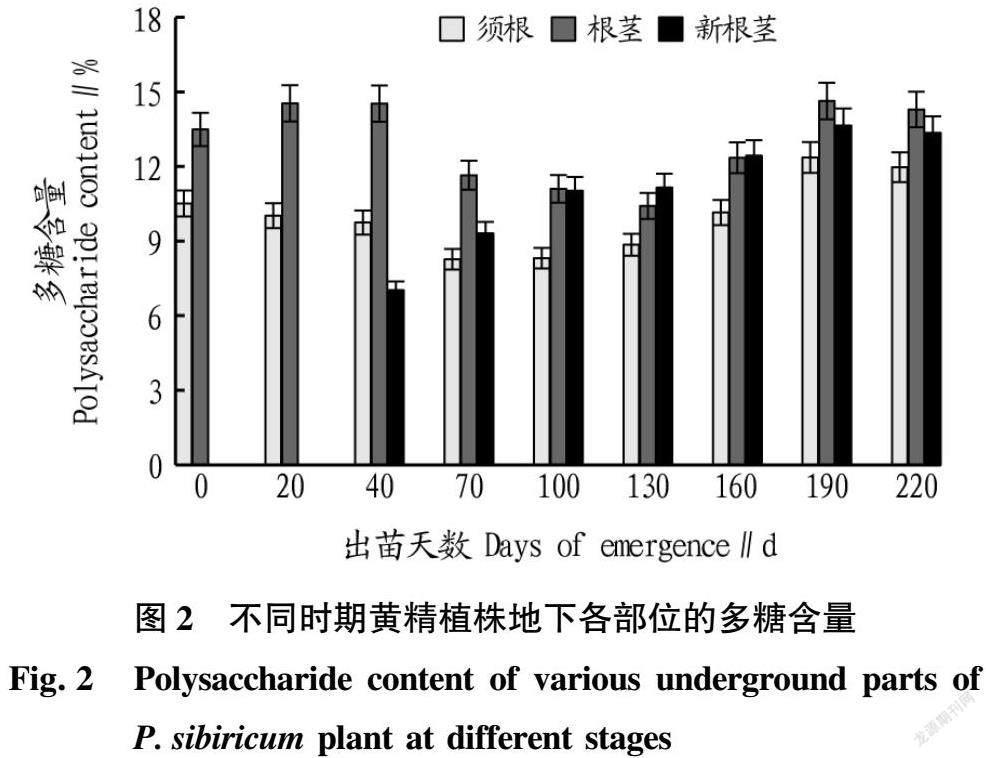
从不同时期黄精植株地下各部位的总皂苷含量变化(图3)可以看出,根茎的总皂苷含量在出苗至20 d较低,之后增加并保持在一个稳定的范围,出苗130 d后持续降低;新根茎形成至出苗70 d,总皂苷含量持续增加,且显著高于根茎(P<0.05),随后新根茎总皂苷含量降低,190 d开始新根茎的总皂苷含量再次增加,但在130 d之后其总皂苷含量显著低于根茎(P<0.05);须根的总皂苷含量大幅低于根茎,刚出苗、出苗100和190 d分别为整个生育期内黄精须根总皂苷含量的3个峰值。以10月中旬为采收期(出苗190 d)采挖的黄精,其地下各部位的总皂苷含量表现为根茎>新根茎>须根,须根的总皂苷含量仅为根茎的35.72%。
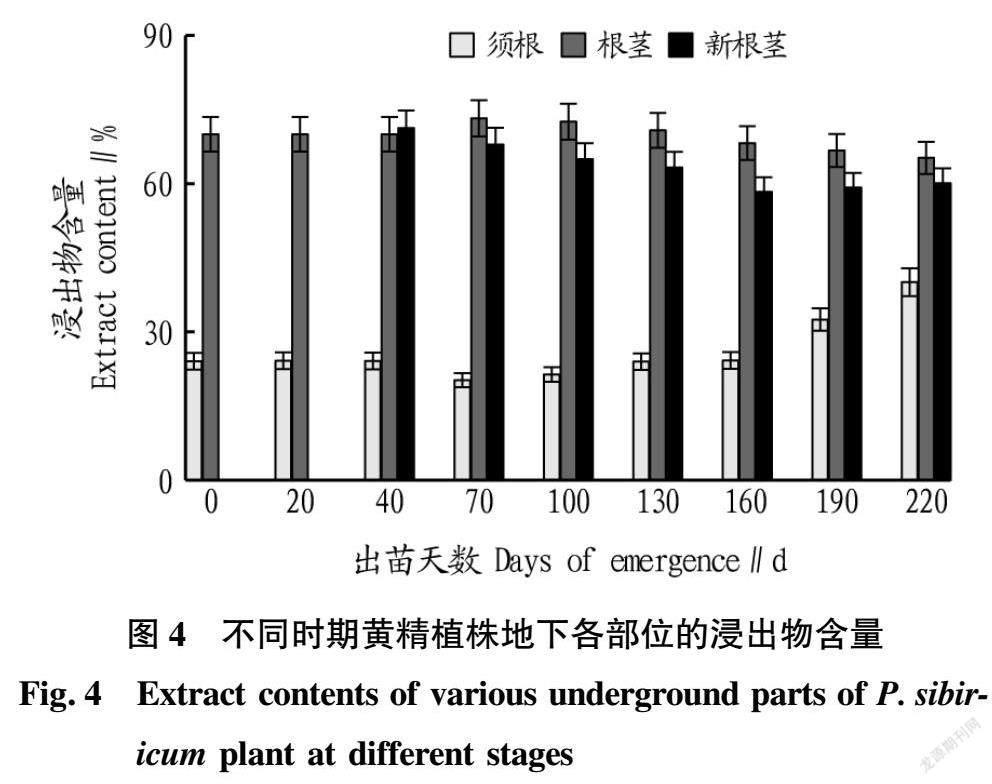
2.3.3 各部位浸出物含量变化。
从不同时期黄精植株地下各部位的浸出物含量变化(图4)可以看出,根茎和新根茎的浸出物含量在整个生育期变化趋势大致相同,根茎的浸出物含量在出苗70 d之前呈现略微上升趋势,随后持续下降;新根茎自形成以后,其浸出物含量持续下降;须根的浸出物含量大幅低于根茎和新根茎,出苗40 d前须根的浸出物含量稳定,随后下降,出苗70 d后持续上升,至出苗220 d,须根的浸出物含量达最大值。以10月中旬为采收期(出苗190 d)采挖的黄精,其地下各部位的浸出物含量表现为根茎>新根茎>须根,须根的浸出物含量仅为根茎的48.68%。
3 讨论
3.1 黄精植株构件生长规律
研究植株构件生长可明确植物不同时期的生长变化规律,便于种植栽培过程中点对点的管理或调控,也可为同属近缘植物植株构件的相关研究提供参考[9]。该研究表明,黄精植株出苗后的40 d内,地上部分迅速生长,至出苗40 d,茎叶总鲜重可达到其周期内最大值的88.73%,与前人在黄精生物学特性的相关研究结果基本一致[10-11]。该研究还发现在整个生育期内,黄精的根系都在不断生长,以茎叶完成伸展后的60 d内(即出苗40~100 d)地下部分积累生物量最为快速,新的根茎在黄精出苗40 d内的花期前后(5月中上旬)形成,受生理和环境因素影响,8月前后的黄精植株茎叶有折损和凋落,生物量减少,由于该研究观察对象为2年生黄精,植株根系布局时间短,养分供应不足,果实形成后不断脱落。
3.2 黄精地下部分有效成分积累规律
随着国家对药用植物非药用部位开发利用的呼吁,已有多种药用植物的非药用部位展开相关研究[12],黄精植株非药用部位的开发利用将会成为下一步研究的目标。通过对整个生育期黄精地下部分各部位的有效成分含量进行动态监测,该研究表明,黄精原生根茎在6—9月份(出苗70~160 d)的多糖含量相对较低,总皂苷含量在4—5月份(出苗0~40 d)相对较低,浸出物含量在6月份之前(出苗0~70 d)较稳定,随后至11月(出苗220 d)持续降低;黄精须根中黄精多糖含量与药用部位差异相对较小,总皂苷和浸出物的含量远低于药用部位,10月收獲期(出苗190 d)的黄精须根多糖含量可达到根茎的84.50%,总皂苷和浸出物含量分别仅为根茎的35.72%和48.68%,这为黄精须根以多糖类材料开发大健康产品提供了理论依据;新根茎形成后的2个月(出苗40~70 d),多糖含量低于根茎,总皂苷含量高于根茎,浸出物含量与根茎相近,随后以上3种成分含量变化趋势与根茎相同;10月份(出苗190 d)黄精地上部分枯萎,进入药材的采收期,黄精地下各部位的3类有效成分含量排序均为根茎>新根茎>须根。
参考文献
[1] 泽让东洲.不同行距和播种量对阿坝垂穗披碱草植株构件组成和种子产量的影响[D].雅安:四川农业大学,2019.
[2] 李漓,周小江,袁志鹰,等.不同种植方式对玄参产量及质量的影响[J].时珍国医国药,2020,31(1):189-191.
[3] 张思唯,李金奥,刘博远,等.打顶方式对雪茄烟烟叶氮素积累及品质的影响[J].作物杂志,2022(1):184-189.
[4] 赵国栋,贾林光,张新生,等.不同时期拉枝对‘宫崎短枝富士’幼树生长和产量、品质的影响[J].北方园艺,2021(13):57-61.
[5] 马存德,常晖,杨祎辰,等.经典名方中黄精的本草考证[J].中国实验方剂学杂志,2022,28(10):193-206.
[6] 翁露,苏娟,赵艳娟,等.药用黄精的栽培和产品开发研究进展[J].安徽农业科学,2022,50(11):8-10.
[7] 国家药典委员会.中华人民共和国药典:2020版一部[S].北京:中国医药科技出版社,2020:319.
[8] 廖念.多花黄精产地加工炮制及其质量标准的研究[D].长沙:湖南中医药大学,2018.
[9] 郭文婷,王国华,缑倩倩,等.河西走廊荒漠绿洲过渡带3种典型一年生藜科植物构件生长及生物量分配特征[J].草业学报,2022,31(2):25-38.
[10] 刘佩.黄精幼苗生长特性及成分积累研究[D].杨凌:西北农林科技大学,2014.
[11] 李勇刚.黄精生物学特性及种子休眠特性的研究[D].杨凌:西北农林科技大学,2009.
[12] 谢蕾,张羽师,李卫东.药用植物非药用部位开发利用现状与展望[J].中药材,2019,42(2):470-473.
猜你喜欢
广州化工(2022年1期)2022-01-26 03:22:32
昆明医科大学学报(2021年6期)2021-07-31 07:39:56
广州化工(2020年23期)2020-12-16 10:57:34
Digital Chinese Medicine(2020年3期)2020-11-03 03:12:12
云南农业科技(2020年5期)2020-10-14 07:24:50
贵州科学(2020年5期)2020-09-02 09:58:08
中成药(2018年6期)2018-07-11 03:01:10
中成药(2017年8期)2017-11-22 03:18:44
特产研究(2016年3期)2016-04-12 07:16:17
药学与临床研究(2015年4期)2015-06-05 11:35:51
