
雷公山自然保护区野猪塘水青冈种群结构动态分析
2023-05-29 02:30:54李登江郑德谋余德会古定豪谢镇国吴必峰
安徽农业科学 2023年9期
李登江 郑德谋 余德会 古定豪 谢镇国 吴必峰
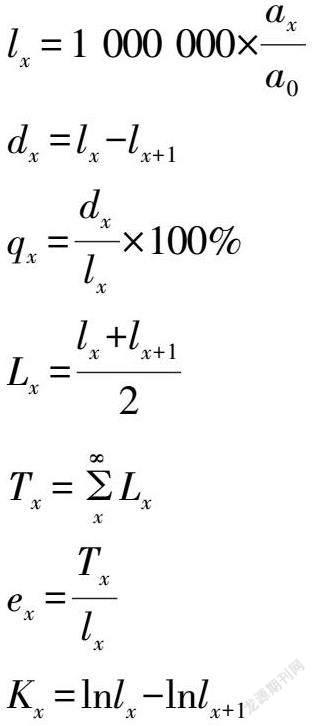
摘要 为揭示水青冈(Fagus longipetiolata)种群结构及其动态特征,选取天然种群集中分布地段,采用相邻格子法设置典型群落样地,从水青冈自然种群的胸径结构特征对其种群结构及数量动态变化进行分析。结果表明:水青冈种群结构整体呈金字塔形;种群的幼龄个体数充足,其个体数量占种群总数量的63.16%,为典型的增长型种群;存活曲线趋于Deevey-Ⅲ型。种群数量随着龄级结构的增加呈减小—增大—减小的趋势,而种群标准化存活个体数lx和标准化存活对数lnlx随着径级的增大逐渐减小,个体平均期望寿命ex随着径级的增加呈先增加后降低的趋势;种群死亡率qx和损失度Kx曲线变化趋势一致,均呈现出降低—升高—降低—升高—降低—升高的复杂动态变化,表明种群易受外界环境干扰,处于极不稳定的状态。
关键词 水青冈;种群结构;静态生命表;雷公山自然保护区
中图分类号 S759.9 文献标识码 A 文章编号 0517-6611(2023)09-0090-03
doi:10.3969/j.issn.0517-6611.2023.09.022
Abstract In order to reveal the population structure and dynamic characteristics of Fagus longipetiolata, the natural population concentrated distribution area was selected, and the adjacent grid method was used to set up typical community sample plots. The population structure and quantitative dynamic changes of Fagus longipetiolata were analyzed from the DBH structure characteristics of the natural population. The results showed that the population structure of Cyclobalanopsis glauca showed a pyramid shape as a whole; The population has sufficient young individuals, accounting for 63.16% of the total population, which is a typical growing population; The survival curve tended to deevey Ⅲ type. The population size showed a decreasing increasing decreasing trend with the increase of age class structure, while the number of standardized surviving individuals lx and the number of standardized surviving logarithm lnlx gradually decreased with the increase of diameter class, and the average life expectancy ex of individuals increased first and then decreased with the increase of diameter class. The trend of mortality qx and loss degree Kx curve is the same, showing a complex dynamic change of decreaseincreasedecreaseincreasedecrease, indicating that the population is vulnerable to external environmental interference and is in a very unstable state.
Key words Fagus longipetiolata;Population structure;Static life table;Leigong Mountain Nature Reserve
基金項目 贵州省天然林保护工程管理中心项目“贵州雷公山常绿落叶阔叶混交林生长演替监测”。
作者简介 李登江(1989—),男,贵州湄潭人,助理工程师,从事森林资源保护与利用研究。
*通信作者,高级工程师,硕士,从事植物分类与生态研究。
种群是物种存在的基本单位[1],种群结构是其最基本特征之一,既可反映种群不同年龄个体的数量分布和配置状况,也可反映种群与环境之间的相互关系[2-3],种群结构与动态是种群生态学研究的核心内容[4],基于种群生命表与其构建的存活曲线以及生存分析理论不仅能反映种群目前的存活状态,也能反映研究物种与环境之间的适合度[5-8],对分析种群现实特征、评估历史干扰、预测其未来演变趋势以及种群的保护和开发利用具有重要意义。
水青冈(Fagus longipetiolata)属壳斗科(Fagaceae)水青冈属(Fagus)落叶乔木[9]。水青冈林既是较好的用材林,又是重要的水源涵养林,产于陕西、浙江、四川、贵州、湖南、广东、福建等省区[10]。此前,有学者对雷公山水青冈林乔木层主要植物种群空间分布格局进行了研究[11],但对雷公山地区水青冈种群结构动态分析方面鲜见报道。鉴于此,笔者以雷公山自然保护区野猪塘水青冈种群为研究对象,从种群年龄结构、静态生命表、存活曲线、死亡率曲线和损失度曲线等方面对其种群结构及数量动态变化进行分析,揭示该区水青冈种群数量变化动态特征、种群动态变化趋势,为水青冈种群下一步的保护与研究提供重要理论支撑。
1 研究区概况
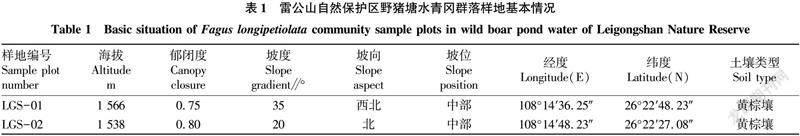
研究区位于雷公山自然保护区内的野猪塘(小地名),该保护区地跨贵州省雷山、台江、剑河、榕江4县,是构成清水江和都柳江主要支流的发源地,处于长江水系与珠江水系的分水岭地带。地理坐标:108°09′~108°22′E,26°15′5~26°22′N,面积47 300 km2,最高海拔2 178.8 m,最低海拔650.0 m。以黄壤为主,黄棕壤为辅,植被为中亚热带东部偏湿性常绿阔叶林、常绿落叶阔叶混交林和山顶苔藓矮林等。年日照时数为1 513.2 h,年太阳总辐射值3 642.5~3 726.3 MJ/m2,无霜期281~290 d,年均温度9.2~16.3 ℃,年均湿度85%~91%,年降雨量1 300~1 600 mm,具有明显立地气候和区域小气候的特点。
2 研究方法
2.1 样地设置与调查方法
在野外踏查的基础上,选取雷公山自然保护区野猪塘水青冈天然群落为研究对象,采用典型样方法,于2019年进行调查,分别设置具代表性的样地2个,每个样地大小为20 m×30 m,总样地面积为1 200 m2,分别记录每个样地的经纬度、海拔、坡度、坡向、坡位等生境因子(表1)。每个样地采用相连格子法,乔木层设置6 m×10 m的小样方10个,调查记录物种名、树高(m)、胸径(cm)、冠幅(m)等因子,灌木层和草本层在样地4角及样地中心位置分别设置1个灌木层(5 m×5 m)和1个草本层(1 m×1 m)样方对灌木层和草本层进行调查。并对样地内胸径(DBH)≥5 cm的水青冈进行每木检尺,统计胸径<5 cm个体数量。
2.2 径级划分与动态量化方法
关于种群大小结构立木径级的划分,许多学者提出了不同的标准[12-18],为避免破坏水青冈野生植物资源,采取空间替代时间的方法,选择径级结构代替年龄结构分析种群动态,即依林木胸径(DBH)大小分级,即Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅶ径级分别对应1、2、3、4、5、6、7龄级。根据样地调查数据和水青冈的生物学特性,将其划分为7个径级分别对应7个龄级,径级划分方法:Ⅰ级(0<DBH<5 cm),Ⅱ级(5 cm≤DBH<10 cm),Ⅲ级(10 cm≤DBH<15 cm),Ⅳ级(15 cm≤DBH<20 cm),Ⅴ级(20 cm≤DBH<25 cm),Ⅵ级(25 cm≤DBH<30 cm),Ⅶ级(DBH≥30 cm)。
2.3 静态生命表的编制
由于静态生命表是反映多个世代重叠的年龄动态历程中的一个特定时间,而不是对这一种群的生活史的追踪,并且调查中存在系统误差,在生命表中会出现死亡率为负的情况。因此,笔者参照江洪[19]在云杉种群生态学中所用的方法对水青冈的初始调查数据进行匀滑处理,进而编制水青冈的种群静态生命表,进而分析其种群结构动态变化。静态生命表的编制遵循以下计算关系式:
式中,a0为种群开始时个体数量,Ax为在x龄级内的现有个体数;ax为匀滑后x龄级内的现存个体数;lnlx为在x龄级开始时的标准化存活个体数;dx为从x到x+1龄级间隔期内的标准化死亡数;qx为从x到x+1龄级间隔期间的死亡率;lx为从x到x+1龄级间隔期间存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级个体的生命期望或平均期望寿命;Kx为消失率(损失度)。
3 结果与分析
3.1 种群分布现状及结构特征
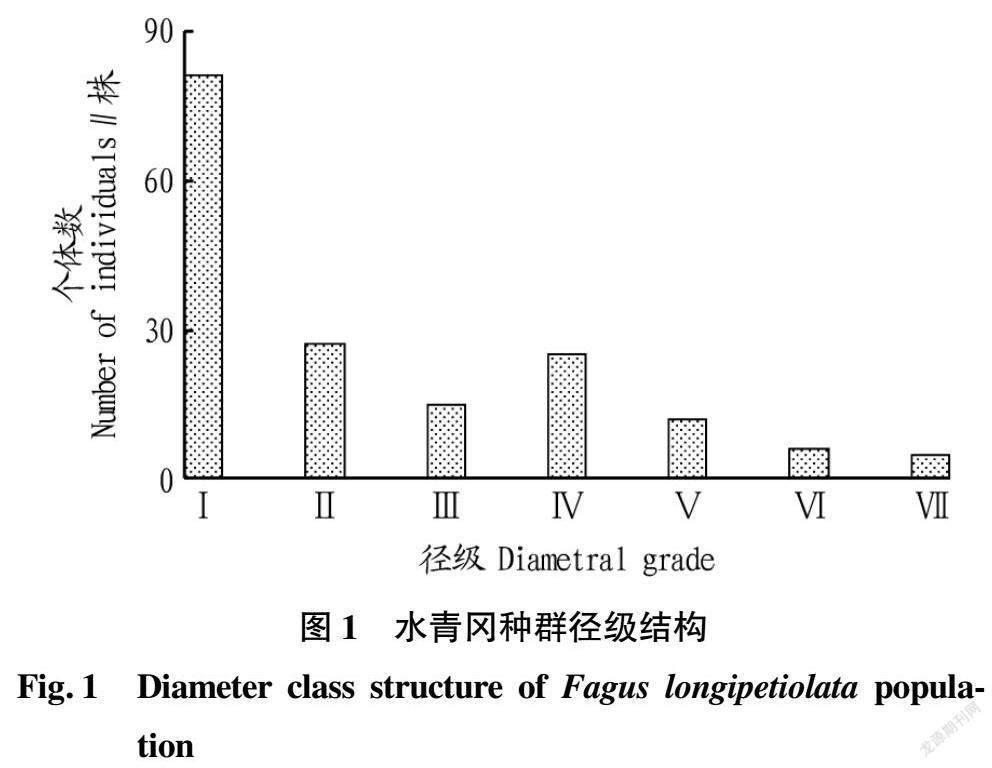
通过对水青冈种群的径级结构统计,可以直观反映出该种群的更新特征。共调查到水青冈共计171 株,水青冈种群个体数在Ⅰ~Ⅱ径级的幼苗幼树个体最多(108株),占种群总数量的63.16%,说明种群中幼龄个体数量充足,水青冈种子自然更新明显。在Ⅲ~Ⅵ径级的中龄个体数(58株),占个体总数量的33.92%,说明其在随后的演替过程中由于种间或种内竞争,乔灌层荫蔽性增强,缺少光照,很多幼龄个体死亡,只有少部分个体存活到中龄个体。Ⅶ径级的老龄个体数量最少,只有5株,该水青冈种群幼龄个体数>中龄个体数>老龄个体数,种群结构呈金字塔形(图1),属于典型的增长型种群。
3.2 种群静态生命特征
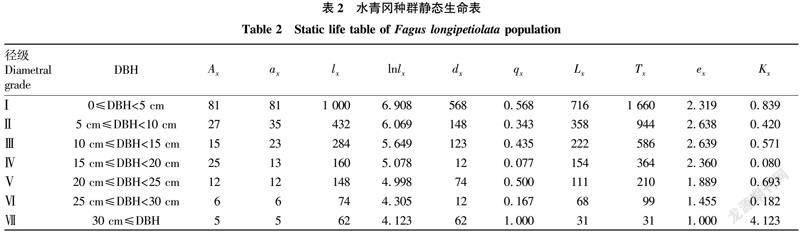
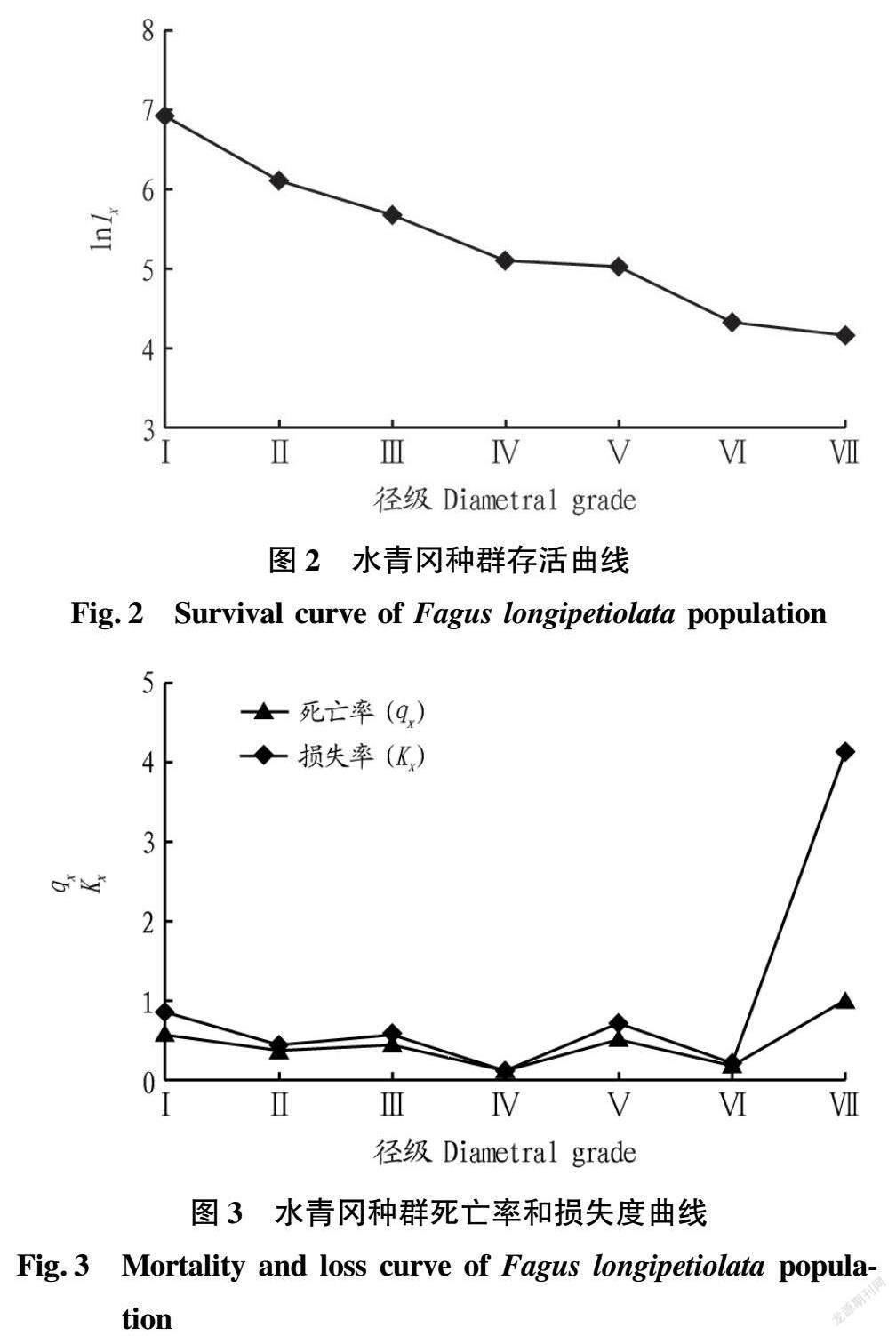
由表2可知,种群数量Ax随着径级结构的增加呈现出减小—增大—减小的趋势,而种群标准化存活个体数lx和标准化存活对数lnlx随着径级的增大逐渐减小,从x到x+1径级间隔期内标准化死亡数dx呈现出下降—上升—下降—上升的趋势;种群死亡率qx随径级的增加呈现出减小—增大—减小—增大—减小—增大的趋势,qx在Ⅳ径级中出现较大变化,其qx明显小于其他径级,说明水青冈种群从Ⅳ径级到Ⅴ径级的演替过程中不易死亡而被淘汰;从x到x+1径级,存活个体数Lx呈現出随径级的增大逐渐减小的趋势;个体平均期望寿命ex随着径级的增加呈现出先增加后降低的趋势;损失度Kx呈现出下降—上升—下降—上升—下降—上升的趋势,其损失度Kx在Ⅳ径级最小,其次为Ⅵ径级。
3.3 死亡率和亏损率曲线
以标准化存活对数lnlx为纵坐标,以径级为横坐标绘制存活曲线(图2),以各径级的死亡率和损失度为纵坐标,以径级为横坐标,绘制死亡率和损失度曲线(图3)。由图2可知,水青冈种群的标准化存活个体数随径级的增大逐渐减小。由图3可知,水青冈种群死亡率qx和损失度Kx曲线变化趋势一致,均呈现出降低—升高—降低—升高—降低—升高的复杂动态变化,波动频繁,表明种群易受外界环境干扰,处于极不稳定的状态。在Ⅳ级时,种群死亡率qx和损失度Kx明显低于其他径级,出现低谷,原因可能是由于Ⅰ~Ⅲ级其受环境强烈筛选,使得Ⅳ级存活下来的种群个体具有很强的生存能力,绝大多数能够顺利过渡到Ⅴ级阶段;而在Ⅴ级时出现高峰的原因可能是向Ⅵ级过渡过程中,随着水青冈径级的增大,种内或种间对光照、水分、生长空间等生存因子的需求加剧,致使大量个体死亡;而Kx在第Ⅶ级为最大值,表明植株个体接近其生理衰老而形成死亡高峰。
4 结论与讨论
该研究结果表明,水青冈种群结构整体呈金字塔形;种群中幼龄个体所占比例最大,中龄树次之,老年树最少,种群的幼龄个体数充足,其个体数量占种群总数量的63.16%,为典型的增长型种群。说明在后期的群落演替过程中,水青冈种群将在很长一段时间内保持增长趋势。
水青冈种群静态生命分析表明,种群数量Ax随着龄级结构的增加呈现出减小—增大—减小的趋势,而种群标准化存活个体数lx和标准化存活对数lnlx随着径级的增大逐渐减小,个体平均期望寿命ex随着径级的增加呈现出先增加后降低的趋势,该研究水青冈种群的存活曲线为 Deevey-Ⅲ型。种群死亡率qx和损失度Kx曲线变化趋势一致,均呈现出降低—升高—降低—升高—降低—升高的复杂动态变化,表明种群易受外界环境干扰,处于极不稳定的状态。
通过实际调查发现,虽然水青冈种群在该区为典型的增长型种群,幼树幼苗充足,种群在后期的群落演替过程中将在很长一段时间内保持增长趋势。但在该区水青冈在群落乔木层中处于绝对的优势地位,导致乔木层物种单一,且在灌木层中,雷山方竹同样占据着绝对的优势地位,且整个群落的郁闭度极高,从而导致灌木层、草本层物种较为单一,这不仅会严重影响水青冈幼苗幼树的生长,也会直接导致水青冈所处群落的物种多样性降低,从而致使群落稳定性减弱,易受外部条件干扰而处于不稳定状态。通过水青冈种群死亡率qx和损失度Kx曲线的复杂动态变化过程也侧面印证了水青冈种群易受外界环境干扰,处于极不稳定的状态。因此,建议针对这一类似的水青冈群落进行适当的人工干预,清除群落中部分影响水青冈生存生长的常见物种,不仅能为灌、草层物种腾出生存空间,增加物种丰富度,也能为水青冈的幼苗幼树生长创造有利条件,促进其水青冈种群朝有利方向发展。
参考文献
[1] 牛翠娟,娄安如,孙儒泳,等.基础生态学[M].北京:高等教育出版社,2002.
[2] 刘普幸.疏勒河中下游绿洲胡杨种群结构与动态研究[J].自然资源学报,2011,26(3):429-439.
[3] 秦爱丽,马凡强,许格希,等.珍稀濒危树种峨眉含笑种群结构与动态特征[J].生态学报,2020,40(13):4445-4454.
[4] 董鸣.生态学透视:种群生态学[M].北京:科学出版社,2015:143-319.
[5] 吴其超,臧凤岐,李呈呈,等.濒危树种五莲杨种群结构与动态特征[J].生态学报,2021,41(12):5016-5025.
[6] 张文辉,许晓波,周建云,等.濒危植物秦岭冷杉种群数量动态[J].应用生态学报,2005,16(10):1799-1804.
[7] 姜在民,和子森,宿昊,等.濒危植物羽叶丁香种群结构与动态特征[J].生态学报,2018,38(7):2471-2480.
[8] 姜顺邦,袁丛军,余德会,等.贵州特有植物黔中杜鹃种群结构及其动态分析[J].浙江林业科技,2020,40(4):1-9.
[9] 方小平,刘映良.水青冈种子萌发研究[J].种子,2009,28(12):23-25.
[10] 郑万钧.中国树木志:第2卷[M].北京:中国林业出版社,1985:2200-2202.
[11] 杨礼旦.雷公山水青冈林乔木层主要植物种群空间分布格局[J].贵州林业科技,2008,36(3):18-21.
[12] 曲仲湘,吴玉树,王焕校,等.植物生态学[M].2版.北京:高等教育出版社,1983.
[13] 何亚平,费世民,蒋俊明,等.不同龄级划分方法对种群存活分析的影响:以水灾迹地油松和华山松种群生存分析为例[J].植物生态学报,2008,32(2):448-455.
[14] 茹文明,張桂萍,毕润成,等.濒危植物脱皮榆种群结构与分布格局研究[J].应用与环境生物学报,2007,13(1):14-17.
[15] 闫淑君,洪伟,吴承祯,等.丝栗栲种群生命过程及谱分析[J].应用与环境生物学报,2002,8(4):351-355.
[16] 洪伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报,2004,15(6):1109-1112.
[17] 江波,周先容,尚进,等.中国特有植物巴山榧树的种群结构与动态[J].生态学报,2018,38(3):1016-1027.
[18] 陈应江,袁丛军,李晓芳,等.贵州楠杆自然保护区楠木种群动态研究[J].安徽农业科学,2019,47(14):122-125.
[19] 江洪.云杉种群生态学[M].北京:中国林业出版社,1992:7-13.
