
大麦籽粒纤维素、半纤维素含量全基因组关联分析
2023-04-12 01:38:24刘新春潘佳佳梁丽静刘美池冯宗云
作物学报 2023年6期
田 敏 刘新春 潘佳佳 梁丽静 董 雷 刘美池 冯宗云
研究简报
大麦籽粒纤维素、半纤维素含量全基因组关联分析
田 敏 刘新春 潘佳佳 梁丽静 董 雷 刘美池 冯宗云*
西南作物基因资源发掘与利用国家重点实验室 / 四川农业大学农学院大麦青稞等健康功能作物研究中心, 四川成都 611130
纤维是衡量大麦饲用品质的重要指标之一, 其遗传机制解析对饲用大麦品种选育具有一定的指导意义。本研究以316份大麦品种为材料, 连续2年种植于成都、康定两地, 利用分布于大麦基因组上的75,289个SNPs对籽粒纤维素、半纤维素含量进行全基因组关联分析。利用TASSEL软件的混合线性模型, 纤维素、半纤维素性状分别重复检测到65个、34个显著SNPs位点(≤0.05/N), 标记的平均解释表型变异率分别为13.18%、14.10%。纤维素、半纤维素含量在3个及以上环境中重复检测到的显著相关位点分别为4个、1个, 2个性状检测到了相同的SNP位点, 说明2个性状存在遗传相关性。纤维素性状挖掘到6个候选基因, 半纤维素性状挖掘到1个候选基因, 为大麦纤维的遗传与分子机理研究及遗传改良奠定了基础。
大麦; 全基因组关联分析; 纤维素; 半纤维素
大麦(L.)是全球第五大粮食作物, 栽培历史悠久, 种植面积广泛。其营养成分种类丰富, 含有多种蛋白、纤维、维生素、矿物质元素以及生物活性物质等, 具有较高的营养价值, 广泛应用于酿造、食品、饲用等领域[1]。膳食纤维影响动物的消化吸收, 其中的纤维含量是影响饲用大麦营养品质的重要因素。可溶性膳食纤维、粗纤维、酸性洗涤纤维、中性洗涤纤维含量等性状被作为大麦饲草品质的重要评价指标[2-4]。已有不少学者对饲草品质性状进行QTL定位分析, 发掘相关性状的遗传位点[5-9]。
纤维素和半纤维素作为大麦纤维的两大成分, 其遗传调控机制成为了研究热点。纤维素合成酶类相关基因是合成纤维素和半纤维素的关键基因, 围绕大麦纤维素合酶基因(cellulose-synthase,)和类纤维素合酶基因(cellulase synthase-like,)的鉴定和功能研究已展开大量工作。大麦基因家族至少有8个成员, 其中、和与初级细胞壁发育有关,、和影响次级细胞壁的构建[10-11]。是半纤维素合成的重要基因家族, 利用生物信息学分析方法, Burton等[12]鉴定到20个基因, 王晓雨[13]鉴定到24个基因, Nishantha等[14]鉴定到18个基因。大麦中,()、等基因也调控纤维素的合成与组装过程, 进而影响细胞壁的发育[15]。水解酶GH9中的内切-(1,4)-β-葡聚糖酶(纤维素酶)能水解(1,4)-β-糖苷键, 使纤维素分解,和等即为该家族基因成员, GH1和GH3家族的一些成员能水解源自纤维素的寡糖[16]。GH9家族中还包括内切木聚糖酶和木葡聚糖酶等, 参与半纤维素的水解[17]。MYB和NAC转录因子是细胞壁生物合成的关键调控因子, 是纤维素和半纤维素合成的主转录开关, 能够起始二者的合成[18-19]。
全基因组关联分析(genome-wide association study, GWAS)是一种通过关联全基因组遗传标记和表型变异, 定位目标性状的遗传位点, 在群体水平解析复杂数量性状的遗传信息的方法[20]。随着测序技术的发展, 许多植物完成了基因组测序工作, GWAS 方法已成为解析植物数量性状遗传构成的重要方法, 广泛应用于水稻[21]、小麦[22]、玉米[23]、油菜[24]等作物性状研究中。在大麦中, 利用GWAS分析了农艺性状[25-26]、营养品质[27-28]、抗逆性[29]、抗病性[30]等性状, 并发掘出控制这些性状变异的遗传位点和功能基因。本研究以316份大麦品种作为材料, 基于前期课题组利用简化基因组开发的高通量SNP分子标记对连续2年种植于成都、康定两地的大麦籽粒纤维素、半纤维含量性状进行全基因组关联分析, 以期挖掘调控相关性状变异的遗传位点或候选基因, 为培育新型专用型大麦品种提供理论基础。
1 材料与方法
1.1 供试材料与田间设计
试验材料由国内外收集的316份大麦品种构成, 其中,国内材料有296份, 主要来自西藏(159份)、青海(50份)、四川(32份)等省(区), 国外材料有6份, 美国和墨西哥各2份, 加拿大和日本各1份, 14份材料未知来源。所有材料均由四川农业大学农学院功能健康作物研究中心提供, 材料编号、类型及来源见附表1。
供试材料于2019年、2020年种植在四川成都, 次年收获, 2020年、2021年种植在四川康定, 当年收获。试点按随机区组试验设计, 行长1.5 m, 行距0.4 m, 株距0.05 m, 每行播30粒种子。生长期间管理措施同大田。成熟期收获种子, 风干后扬净贮藏、选用颗粒大小相近的籽粒用粉碎机粉碎, 干燥保存备用。
1.2 品质测定与表型统计
参照安玉民等[31]方法测定纤维素含量和半纤维素含量, 重复3次。
通过Microsoft Excel 2010分析整理数据, 利用SPSS 25.0软件对数据进行描述性统计、方差分析及相关性分析。
1.3 群体结构分析
采用Structure 2.3.4软件, 基于贝叶斯数学模型对供试的316个大麦种质资源进行群体结构分析, 将设置为2~10, 计算每个值下的Δ值, 绘制Δ值随值变化的折线图, 确定群体亚群数。
1.4 全基因组关联分析
利用本课题组前期已获得该关联分析群体的75,289个SNP标记分型数据, 运用TASSEL 5.2.8软件中的混合线性模型(mixed linear model, MLM)进行纤维素含量、半纤维素含量全基因组关联分析研究。将显著性临界值设置为=0.05/=6.64×10–7(为本研究的标记数)。
1.5 候选基因预测
根据关联分析结果, 将发掘到显著性SNP标记的物理位置与大麦基因组数据库中序列的物理位置进行比对, 以上下游各延伸200 kb作为候选基因区段。利用大麦基因组网站中的基因功能注释信息, 挖掘性状相关候选基因。
2 结果与分析
2.1 纤维素、半纤维素表型分析
2年两点大麦籽粒纤维素、半纤维素含量的基本描述统计如表1。2019成都(19 CD)、2020康定(20 KD)、2020成都(20 CD)、2021康定(21 KD)环境纤维素含量均值分别为9.12%、9.82%、8.37%、8.73%, 最低为2.65%, 最高为22.29%; 半纤维素含量均值分别为12.58%、12.37%、11.47%、12.91%, 最低为6.99%, 最高为27.74%, 各环境下2个性状的变异系数在22.00%~45.28%之间, 说明材料间表型差异明显, 不同环境下群体的整体表现有所不同。纤维素、半纤维素含量在4个环境的偏度均大于0, 呈正偏态, 峰度多小于1.5, 整体看成拟合正态分布, 符合数量性状特征。
多点联合方差分析表明, 基因型、环境以及基因型与环境互作存在极显著差异(表2), 说明关联群体纤维素、半纤维素含量遗传变异丰富, 且受环境影响较大, 广义遗传率分别为79.45%、75.29%, 说明遗传因素是影响性状的主要因素。4个环境两性状的相关系数分别为0.313、0.880、0.976和0.977, 在<0.01水平上显著相关, 表明2个性状存在相关性。

表1 不同试点大麦籽粒纤维素和半纤维素含量的表现
CD: 成都; KD: 康定。CD: Chengdu; KD: Kangding.
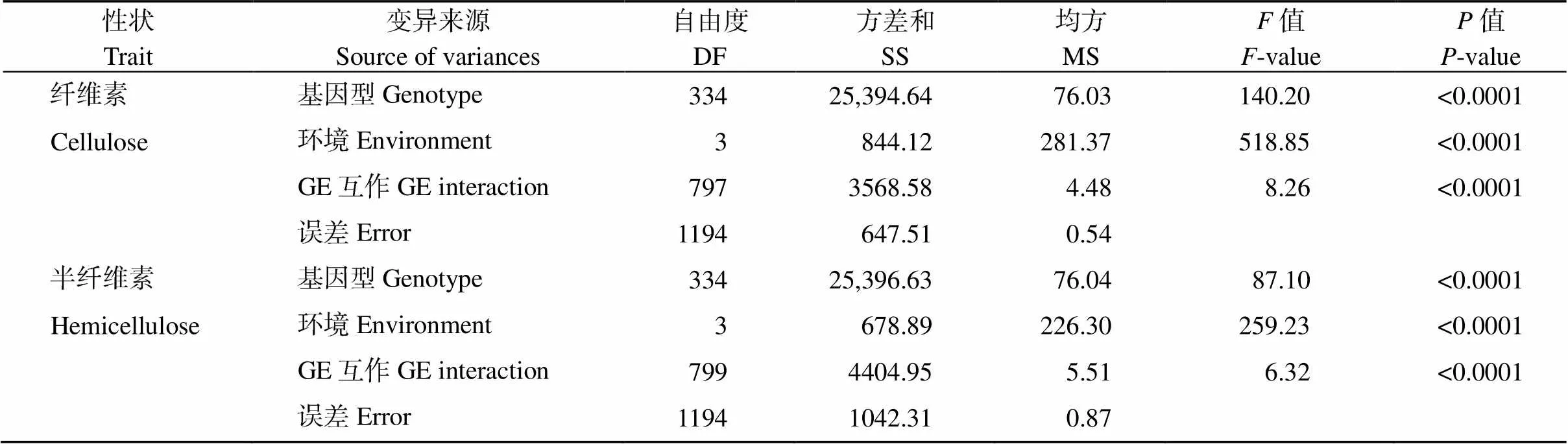
表2 大麦籽粒纤维素和半纤维素含量的多环境联合方差
2.2 群体结构分析
采用Structure软件对供试材料进行群体结构分析。根据计算出的Δ值绘制Δ值随值变化(= 2~10)的折线图(图1-A),=3时Δ出现明显的峰值, 其余值对应的Δ值均趋于0, 因此选定=3为最适的群体数目, 将本群分为3个亚群(图1-B)。第1亚群包含102份材料(32.28%), 第2亚群包含139份材料(43.99%), 第3亚群包含75份材料(23.73%)。其中来源于西藏的159份材料主要位于第2亚群, 占该类群总数的71.07%, 青海的50份材料中有35份分布于第1亚群(64.00%), 四川的32份材料中有23份分布于第1亚群(71.88%), 贵州、河南、黑龙江、江苏、山西、陕西等省的材料主要分布于第3亚群, 同一来源地的材料之间亲缘关系较近。
2.3 全基因组关联分析
利用混合线性模型(MLM)对籽粒纤维素、半纤维素含量进行全基因组关联分析(图2)。在4个环境重复检测到65个纤维素含量显著SNP分子标记位点, 这些位点可解释的平均表型变异率为13.18%, 分布在大麦所有的染色体上, 其中在5H染色体上发现的显著性标记数最多, 有21个。2号染色体上的SNP位点25574694和5号染色体上的SNP位点471635005在2020康定、2020成都、2021康定3个环境中检测到, 2号染色体上的SNP位点28768234和5号染色体上的SNP位点437943810在2019成都、2020成都、2021康定3个环境中检测到, 在2020成都和2021康定发现较多相同的显著位点。在4个环境重复检测到34个半纤维素含量相关的显著SNP位点, 位点的平均表型变异率为14.10%, 在5H染色体上发现的显著性标记数最多, 有14个, 其余染色体上也定位到显著相关位点。在2019成都、2020成都和2021康定3个环境中检出一个相同的显著位点, 为2号染色体上的SNP位点28768234, 在2020成都和2021康定检出较多相同的显著位点。纤维素和半纤维素有多个相同显著SNP位点, 也有遗传距离相隔极近的显著位点, 说明2个性状遗传上具有相关性(附表2和附表3)。

图1 316份大麦材料群体结构分析
A: 通过假定不同值计算得到的Δ值; B: 通过Structure软件分析的遗传结构(红色: 第1亚群; 绿色: 第2亚群; 蓝色: 第3亚群)。
A: Δ-value calculated by assuming different-values; B: genetic structure analyzed by structure software (red: the first subgroup; green: the second subgroup; blue: the third subgroup).
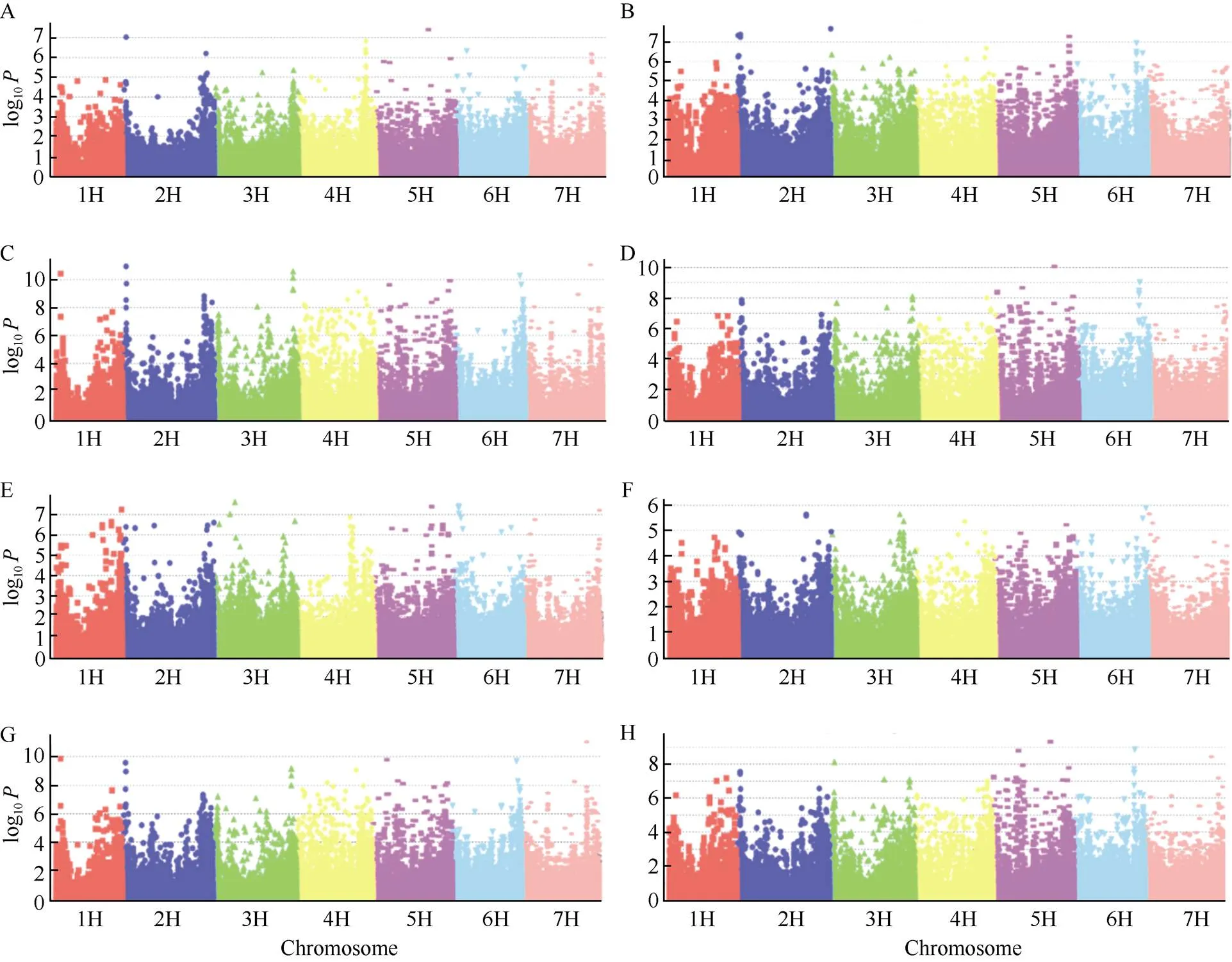
图2 不同环境大麦籽粒纤维素、半纤维素含量全基因组关联分析
A、B、C、D分别表示2019成都、2020康定、2020成都、2021康定纤维素含量的曼哈顿图; E、F、G、H分别表示2019成都、2020康定、2020成都、2021康定半纤维素含量的曼哈顿图。
A. B, C, and D represent Manhattan diagram of cellulose content in 2019 Chengdu, 2020 Kangding, 2020 Chengdu, and 2021 Kangding, respectively; E, F, G, and H represent the Manhattan diagram of hemicellulose content in 2019 Chengdu, 2020 Kangding, 2020 Chengdu, and 2021 Kangding, respectively.
2.4 预测候选基因
据关联分析结果, 在4个环境重复3次检测到的性状显著位点上下共400 kb区间范围内寻找候选基因。根据已知纤维素、半纤维素基因的编码蛋白类型与数据库候选基因功能注释信息, 初步筛选到6个与纤维素有关的候选基因, 1个与半纤维素相关的候选基因,和编码UDP-糖基转移酶, 其中也是半纤维素相关的候选基因,和编码糖转运蛋白,编码UDP-葡萄糖4-差异构酶家族蛋白,编码内切葡聚糖酶(表3)。
3 讨论
关联分析是解析复杂性状遗传基础的常用方法, 群体大小、标记数量、表型数据获取和统计分析方法等因素会影响关联分析的准确性, 因此需要从多方面进行考虑, 以确保结果的准确性和可靠性[32-34]。为提高结果可信度, 可将关联分析与其他分析方法整合, 如GWAS与QTL定位结合[35-36]、GWAS与WGCNA (加权基因共表达网络分析)结合[37-38]等, 同时可对结果进行进一步的验证。
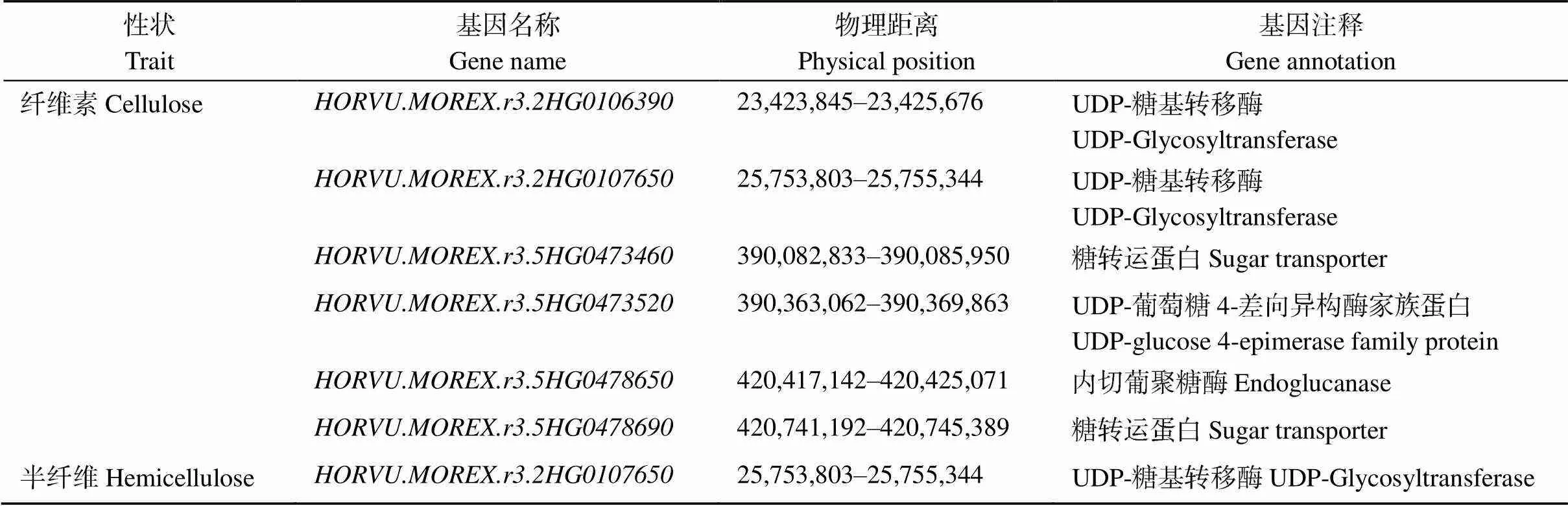
表3 纤维素、半纤维素含量相关候选基因信息
本研究对纤维素和半纤维素含量进行相关分析发现, 2个性状在4个环境下均存在显著的正相关性, 说明二者紧密联系, 相互影响。后续关联分析2个性状发掘出相同的显著SNP位点, 马珍珍等[39]对甘蓝型油菜中的纤维素和半纤维素含量QTL分析也发现二者存在重叠区段, 说明纤维素和半纤维素性状具有一定的遗传相关性。海拔高度、土壤土质、气候条件、农艺措施等会影响植物的生长状况, 从而改变植物的性状表现。从试验结果来看, 不同地区、不同年份大麦籽粒纤维素和半纤维素含量存在差异,环境对2个性状有较大影响。因此在全基因组关联分析时, 主要关注检测出的重合位点, 将在3个及以上环境中重复检测出的显著位点作为2个性状的遗传调控位点, 并在这些位点的目标区段挖掘候选基因。
2个性状分别预测到了6个、1个候选基因, 皆是酶类基因, 涉及糖的合成、转运与分解过程。和均编码糖基转移酶, 具有尿苷二磷酸(UDP)糖基转移酶活性。纤维素和半纤维素的合成以核苷酸糖为底物, 尿苷二磷酸葡萄糖是纤维素合成的重要底物, 多种尿苷二磷酸糖和鸟苷二磷酸糖可作为糖基转移酶的底物参与半纤维素的合成。Nishantha等[14]利用GWAS发掘影响大麦秆纤维素含量的候选基因, 鉴定到2个UDP-葡萄糖基转移酶基因(和), 并对进行共表达分析, 结果推测其调控次生壁纤维素的合成。和编码糖转运蛋白。核苷酸糖多在胞质中合成, 然后转运到高尔基体腔, 这一过程需要定位于高尔基体的核苷酸糖转运蛋白。水稻OsNST1是一种UDP-葡萄糖转运蛋白,点突变后, 细胞壁中的单糖葡萄糖含量降低, 纤维素的含量也减小, 而其他细胞壁多糖的含量增加, 表明高尔基体定位的核苷酸糖转运蛋白对细胞壁生物合成的调控作用[40]。编码内切葡聚糖酶, 是糖基水解酶9 (cellulase E)家族成员, 能水解(1,4)-β-糖苷键, 参与纤维素的分解过程。
纤维素和半纤维素是大麦食用纤维的重要成分, 作为不可溶性膳食纤维具有通便之效。在大鼠食物中添加大麦膳食纤维, 与对照相比, 大鼠的增重较少, 排便增多, 胃肠道运输能力改善[41]。对人类来说, 膳食纤维能调节肠道菌群结构, 通过物理效应、免疫调节、结肠发酵等多种机制来改善机体健康状况, 有助于防治肥胖症、糖尿病、结肠癌等疾病[42]。纤维也是考察大麦饲用品质的重要指标, 粗饲料干物质中粗纤维含量应在18%以上, 中性洗涤纤维、酸性洗涤纤维影响饲料消化率、潜在采食量, 含量越低, 粗饲料品质越好[43]。茎秆强度与作物的产量、抗性直接相关, 是一个重要的农艺性状, 次生壁的组分结构与植株茎秆机械强度直接相关。次生壁纤维素含量降低, 会使壁变薄, 植株机械强度降低, 半纤维素含量变化也会引起茎秆强度的改变[44-45]。可见, 纤维素和半纤维素含量对于大麦的营养价值、饲用品质和农艺性状具有一定影响, 有重要的研究意义。
附表 请见网络版: 1) 本刊网站http://zwxb.chinacrops. org/; 2) 中国知网http://www.cnki.net/; 3) 万方数据http:// c.wanfangdata.com.cn/Periodical-zuowxb.aspx。
[1] 许伟利, 董伟志, 王军, 徐晶, 马云, 沙元赛. 大麦籽粒营养成分及开发研究进展. 大麦与谷类科学, 2019, 36(3): 52–55. Xu W L, Dong W Z, Wang J, Xu J, Ma Y, Sha Y S. Research progresses on the nutrients of barley grain and their utilization., 2019, 36(3): 52–55 (in Chinese with English abstract).
[2] 赵斌, 陈晓东, 季昌好, 朱斌, 王瑞. 不同刈割时期与干燥方式对大麦饲草品质的影响. 草原与草坪, 2020, 40(5): 98–101. Zhao B, Chen X D, Ji C H, Zhu B, Wang R. Effects of different cutting times and drying methods on the quality of barley forage., 2020, 40(5): 98–101 (in Chinese with English abstract).
[3] 赵加涛, 杨向红, 付正波, 字尚永, 刘猛道. 不同大麦品种饲草产量及品质研究. 中国农学通报, 2021, 37(27): 27–31. Zhao J T, Yang X H, Fu Z B, Zi S Y, Liu M D. Yield and quality of forage grass of different barley varieties., 2021, 37(27): 27–31 (in Chinese with English abstract).
[4] 黄水珍, 冯德庆, 黄秀声, 黄小云. 大麦‘花22’不同生育期的饲用品质及呕吐毒素含量. 农学学报, 2021, 11(4): 1–5. Huang S Z, Feng D Q, Huang X S, Huang X Y. Forage quality and vomitoxin content of barley Hua 22 at different growth stages., 2021, 11(4): 1–5 (in Chinese with English abstract).
[5] Han F, Ullrich S E, Romagosa I, Clancy J A, Froseth J A, Wesenberg D M. Quantitative genetic analysis of acid detergent fiber content in barley grain., 2003, 38: 167–172.
[6] Siahsar B A, Peighambari S A, Taleii A R, Naghavi M R, Nabipour A, Sarrafi A. QTL analysis of forage quality traits in barley (L.)., 2009, 37: 479–488.
[7] Grando S, Baum M, Ceccarelli S, Goodchild A, El-Haramein F Jaby, Jahoor A, Backes G. QTLs for straw quality characteristics identified in recombinant inbred lines of a× H. spontaneum cross in a Mediterranean environment., 2005, 110: 688–695.
[8] Surber L, Abdel-Haleem H, Martin J, Hensleigh P, Cash D, Bowman J, Blake T. Mapping quantitative trait loci controlling variation in forage quality traits in barley., 2011, 28: 189–200.
[9] Abdel-Haleem H, Bowman J G P, Surber L, Blake T. Variation in feed quality traits for beef cattle in Steptoe×Morex barley population., 2012, 29: 503–514.
[10] Burton R A, Shirley N J, King B J, Harvey A J, Fincher G B. The CesA gene family of barley. quantitative analysis of transcripts reveals two groups of co-expressed genes., 2004, 134: 224–236.
[11] Burton R A, Jobling S A, Harvey A J, Shirley N J, Mather D E, Bacic A, Fincher G B. The genetics and transcriptional profiles of the cellulose synthase-likegene family in barley., 2008, 146: 1821–1833.
[12] Burton R A, Ma G, Baumann U, Harvey A J, Shirley N J, Taylor J, Pettolino F, Bacic A, Beatty M, Simmons C R, Dhugga K S, Rafalski J A, Tingey S V, Fincher G B. A customized gene expression microarray reveals that the brittle stem phenotype fs2 of barley is attributable to a retroelement in thecellulose synthase gene., 2010, 14: 1716–1728.
[13] 王晓雨. 大麦β-葡聚糖、微量元素含量的全基因组关联分析及纤维素合成酶类基因家族的鉴定. 西北农林科技大学硕士学位论文, 陕西杨凌, 2020. Wang X Y. Genome-wide Association Study of the β-glucan and Trace Elements Content, and Identification of Cellulose Synthase Gene Family in Barley (). MS Thesis of Northwest A&F University, Yangling, Shaanxi, China, 2020(in Chinese with English abstract).
[14] Nishantha M D L C, Jeewani D C, Xing G W, Nie X J, Song W N. Genome-wide identification and analysis of thegene family barley (L.)., 2020, 10: 122–126.
[15] Houston K, Burton R A, Sznajder B, Rafalski A J, Dhugga K S, Mather D E, Taylor J, Steffenson B J, Waugh R, Fincher G B. A genome-wide association study for culm cellulose content in barley reveals candidate genes co-expressed with members of the CELLULOSE SYNTHASE A gene family., 2015, 10: e0130890.
[16] Buchanan M, Burton R A, Dhugga K S, Rafalski A J, Tingey S V, Shirley N J, Fincher G B. Endo-(1, 4)-β-Glucanase gene families in the grasses: temporal and spatial. Co-transcription of orthologous genes., 2012, 12: 235.
[17] Cantarel B L, Coutinho P M, Rancurel C, Bernard T, Lombard V, Henrissat B. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for glycogenomics., 2009, 37: D233–D238.
[18] Zhong R Q, Ye Z H. Complexity of the transcriptional network controlling secondary wall biosynthesis., 2014, 229: 193–207.
[19] Zhong R, Ye Z H. Secondary cell walls: biosynthesis, patterned deposition and transcriptional regulation., 2015, 56: 195–214.
[20] 赵宇慧, 李秀秀, 陈倬, 鲁宏伟, 刘羽诚, 张志方, 梁承志. 生物信息学分析方法Ⅰ: 全基因组关联分析概述. 植物学报, 2020, 55: 715–732. Zhao Y H, Li X X, Chen Z, Lu H W, Liu Y C, Zhang Z F, Liang C Z. Bioinformatics analysis methods. I: Overview of genome-wide association analysis., 2020, 55: 715–732 (in Chinese with English abstract).
[21] Yang W, Zhao J L, Zhang S H, Chen L, Yang T F, Dong J F, Fu H, Ma Y M, Zhou L, Wang J, Liu W, Liu Q, Liu B. Genome-wide association mapping and gene expression analysis reveal the negative role ofin regulating bacterial blight resistance in rice.(NY), 2021, 14: 58.
[22] Liu P, Jin Y R, Liu J D, Liu C Y, Yao H P, Luo F Y, Guo Z H, Xia X C, He Z H. Genome-wide association mapping of root system architecture traits in common wheat (L.)., 2019, 215: 121.
[23] Wang M, Yan J B, Zhao J R, Song W, Zhang X B, XiaoY N, Zheng Y L. Genome-wide association study (GWAS) of resistance to head smut in maize., 2012, 196: 125–131.
[24] Wrucke D F, Mamidi S, Rahman M. Genome-wide association study for frost tolerance in canola (L.) under field conditions., 2019, 28: 211–222.
[25] 聂石辉, 王仙, 向莉, 张金汕, 李志强, 任毅, 方伏荣. 干旱胁迫对中亚大麦农艺性状的影响及其相关基因定位. 麦类作物学报, 2022, 42: 59–67. Nie S H, Wang X, Xiang L, Zhang J S, Li Z Q, Ren Y, Fang F R. Responses to drought stress and gene mapping of related agronomic traits of central Asian barley., 2022, 42: 59–67 (in Chinese with English abstract).
[26] He T H, Beate H C, Tolera A T, Zhang X Q, Chen K F, David M, Paul T, Sharon W, Li C D. Gene-set association and epistatic analyses reveal complex gene interaction networks affecting flowering time in a worldwide barley collection., 2019, 70: 5603–5616.
[27] Fan X Y, Sun Y D, Zhu J, Lyu C, Guo B J, Xu R G. A 191-bp insertion/deletion in GBSS1 region is responsible for the changes in grain amylose content in barley (L.)., 2017, 37: 81.
[28] 耿腊, 黄业昌, 李梦迪, 谢尚耿, 叶玲珍, 张国平. 大麦籽粒β-葡聚糖含量的全基因组关联分析. 作物学报, 2021, 47: 1205–1214. Geng L, Huang Y C, Li M D, Xie S G, Ye L Z, Zhang G P. Genome-wide association study of β-glucan content in barley grains., 2021, 47: 1205–1214 (in Chinese with English abstract).
[29] Hazzouri K M, Hazzouri K M, Khraiwesh B, Amiri K M A, Amiri K M A, Pauli D, Blake T, Mullath M S S K, Mansour D N A L, Salehi-Ashtiani K, Purugganan M, Masmoudi K. Mapping ofgene in barley using GWAS approach and its implication in salt tolerance mechanism., 2018, 9: 156.
[30] Aghnoum R, Bvindi C, Menet G, Hoop B D, Maciel L N, Niks R E. Host/nonhost status and genetics of resistance in barley against three pathotypes of Magnaporthe blast fungi., 2019, 215: 116.
[31] 安玉民, 王菊葵, 黄烨, 徐晓梅. 马铃薯秸秆中纤维素与半纤维素含量的测定. 现代农业科技, 2016, (17): 159–160. An Y M, Wang J K, Huang Y, Xu X M. Determination of cellulose and hemicellulose content in potato stalk., 2016, (17): 159–160 (in Chinese with English abstract).
[32] Zhu C S, Gore M, Buckler E S, Yu J M. Status and prospects of association mapping in plants., 2008, 1: 5–20.
[33] Kaler A S, Purcell L C. Estimation of a significance threshold for genome-wide association studies., 2019, 20: 618.
[34] Alqudah A M, Sallam A, Baenziger P S, Brner A. GWAS: Fast-forwarding gene identification and characterization in temperate Cereals: lessons from barley: a review., 2020, 22: 119–135.
[35] Lou Q J, Chen L, Mei H W, Wei H B, Feng F J, Wang P, Xia H, Li T M, Luo L J. Quantitative trait locus mapping of deep rooting by linkage and association analysis in rice., 2015, 66: 4749–4757.
[36] Zhang X, Ren Z Y, Luo B W, Zhong H X, Ma P, Zhang H K, Hu H M, Wang Y K, Zhang H Y, Liu D, Wu L, Nie Z, Zhu Y H, He W Z, Zhang S Z, Su S Z, Shen Y O, Gao S B. Genetic architecture of maize yield traits dissected by QTL mapping and GWAS in maize., 2022, 10: 436–446.
[37] Ma L L, Zhang M Y, Chen J, Qing C Y, He S J, Zou C Y, Yuan G S, Yang C, Peng H, Pan G T, Lübberstedt T, Shen Y O. GWAS and WGCNA uncover hub genes controlling salt tolerance in maize (L.) seedlings., 2021, 134: 3305–3318.
[38] Li K Q, Wang J, Kuang L Q, Tian Z, Wang X F, Dun X L, Tu J X, Wang H Z. Genome-wide association study and transcriptome analysis reveal key genes affecting root growth dynamics in rapeseed., 2021, 14: 178.
[39] 马珍珍, 李加纳, Wittkop B, Frauen M, 阎星颖, 刘列钊, 肖阳. 甘蓝型油菜籽粒含油量、蛋白质、纤维素及半纤维素含量QTL分析. 作物学报, 2013, 39: 1214–1222. Ma Z Z, Li J N, Wittkop B, Frauen M, Yan X Y, Liu L Z, Xiao Y. QTL mapping for oil, protein, cellulose, and hemicellulose contents in seeds ofL., 2013, 39: 1214–1222 (in Chinese with English abstract).
[40] Zhang B C, Liu X L, Qian Q, Liu L F, Dong G J, Xiong G Y, Zeng D L, Zhou Y H. Golgi nucleotide sugar transporter modulates cell wall biosynthesis and plant growth in rice., 2011, 108: 5110–5115.
[41] Ryae J J, Hyuk C J. Lactic acid fermentation of germinated barley fiber and proliferative function of colonic epithelial cells in loperamide-induced rats., 2010, 13: 950–960.
[42] Gong J S, Yang C B. Advances in the methods for studying gut microbiota and their relevance to the research of dietary fiber functions., 2012, 48: 916–929.
[43] Li H Y, Xu L, Liu W J, Fang M Q, Wang N. Assessment of the nutritive value of whole corn stover and its morphological fractions., 2014, 27: 194–200.
[44] Zhang B C, Deng L W, Qian Q, Xiong G Y, Zeng D, Li R, Guo L B, Li J Y, Zhou Y H. A missense mutation in the transmembrane domain of CESA4 affects protein abundance in the plasma membrane and results in abnormal cell wall biosynthesis in rice., 2009, 71: 509–524.
[45] Vega-Sánchez M E, Verhertbruggen Y, Christensen U, Chen X, Sharma V, Varanasi P, Jobling S A, Talbot M, White R G, Joo M. Loss of cellulose synthase-like f6 function affects mixed-linkage glucan deposition, cell wall mechanical properties, and defense responses in vegetative tissues of rice., 2012, 159: 56–69.
Genome-wide association analysis of cellulose content and hemicellulose content in grains of barley
TIAN Min, LIU Xin-Chun, PAN Jia-Jia, LIANG Li-Jing, DONG Lei, LIU Mei-Chi, and FENG Zong-Yun*
State Key Laboratory of Crop Gene Exploration and Utilization in Southwest China / Research Center for Healthy Functional Crops such as Barley and Hulless Barley / College of Agronomy, Sichuan Agricultural University, Chengdu 611130, Sichuan, China
Fiber is one of the important indexes to evaluate the forage quality of barley, and the analysis of its genetic mechanism has important guiding significance for the selection and breeding of forage barley. In this study, 316 barley varieties were planted in Chengdu and Kangding for two consecutive years, and 75,289 SNPs distributed in barley genome were used for genome-wide association analysis of grain cellulose and hemicellulose content, 65 and 34 significant SNPs (≤ 0.05/N) were repeatedly detected for cellulose and hemicellulose traits using the mixed linear model of TASSEL software, respectively. The mean explained phenotypic variation rates of markers were 13.18% and 14.10%, respectively. There were four and one significant correlation loci repeatedly detected for cellulose and hemicellulose content in three or more environments, respectively. The same SNP loci were detected for two traits, indicating that there was genetic correlation between the two traits. Six candidate genes were found for cellulose traits, and one candidate gene was found for hemicellulose traits, which laid the foundation for genetic and molecular mechanism research and genetic improvement of barley fiber.
barley; genome-wide association analysis; cellulose; hemicellulose
10.3724/SP.J.1006.2023.21043
本研究由国家重点研发计划项目(2018YFD1000705), 四川省国际科技创新合作项目(2021YFH0113)和财政部和农业农村部国家现代农业产业技术体系建设专项(大麦青稞, CARS-05)资助。
This study was supported by the National Key Research and Development Program of China (2018YFD1000705), the Sichuan International Science and Technology Innovation Cooperation Project (2021YFH0113), and the China Agriculture Research System of MOF and MARA (Barley and Hulless Barley, CARS-05).
冯宗云, E-mail: zyfeng49@126.com
E-mail: 2030214850@qq.com
2022-06-22;
2022-11-27;
2022-12-13.
URL: https://kns.cnki.net/kcms/detail//11.1809.S.20221209.1820.002.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
Asian Herpetological Research(2022年4期)2023-01-05 03:03:46
作文小学高年级(2022年5期)2022-06-16 06:22:50
环境卫生工程(2021年4期)2021-10-13 06:52:26
纺织科技进展(2021年3期)2021-06-09 08:07:14
陶瓷学报(2021年1期)2021-04-13 01:33:02
地震研究(2021年1期)2021-04-13 01:04:48
疯狂英语·新读写(2021年2期)2021-02-25 08:58:46
阿来研究(2020年1期)2020-10-28 08:09:58
华人时刊(2016年19期)2016-04-05 07:56:10
林业与生态(2016年3期)2016-02-27 14:24:18
