
西沙宣德群岛海域浮游植物群落及其影响因子
2023-04-11 03:16刘晓红石军
热带亚热带植物学报 2023年2期
刘晓红, 石军
西沙宣德群岛海域浮游植物群落及其影响因子
刘晓红1, 石军2
(1. 国家海洋局三沙海洋环境监测中心站, 海口 570311; 2. 国家海洋局海口海洋环境监测中心站, 海口 570311)
为了解西沙宣德群岛海域浮游植物群落结构特征,于2019年2月和11月对该海域进行采样调查,分析浮游植物群落组成及环境影响因素。两个航次共发现109种浮游植物,其中硅藻门最多,有81种。冬秋季节优势种不尽相同,冬季优势种为铁氏束毛藻()、红海束毛藻()和标志星杆藻()等,秋季优势种为菱形海线藻()、丹麦细柱藻()和劳氏角毛藻()。冬季浮游植物平均丰度[(5.27±6.14)×107cells/L]显著高于秋季[(1.56±1.40)×105cells/L]。浮游植物群落分布主要受亚硝酸盐、盐度等环境因子影响。冬秋季各站位浮游植物群落的多样性指数、均匀度指数和丰富度指数平均值分别为3.15、0.71、1.36和3.28、0.75、1.77。因此,宣德群岛海域冬秋季节浮游植物群落多样性较高,物种较为丰富,群落结构复杂稳定。
西沙宣德群岛;浮游植物;环境因子;群落结构
浮游植物对海洋生态系统的物质循环和能量流动起着至关重要的作用,浮游植物的光合作用为海洋提供了95%左右的初级生产力,产生的有机碳总量是高等植物的7倍左右,对CO2的吸收量约为全球年产量的一半[1–3]。同时,浮游植物群落能对生活环境的变化做出快速的响应,直接影响水质、生态系统物质流动、能量流动和生物资源的变动[4], 因而常作为评价海洋生态系统健康状况的一种指标[5]。浮游植物不仅是海水中溶解氧的“主力军”, 也是海洋碳氮流通的主要贡献者,为鱼虾贝类等提供直接或间接的食物来源[6]。
西沙宣德群岛位于西沙群岛的东侧偏北,在北纬15°43′~17°00′,东经112°10′~112°54′的海域上,其主体为宣德环礁,包括永兴岛-石岛、七连屿等岛屿。宣德群岛珊瑚礁生态系统是我国现存珊瑚礁群落中最古老最原始的群落,是我国沿海区域珊瑚礁生态系统的发源地,也是我国近海海域保存相当完好和珍贵的珊瑚礁区域[7]。目前对于该海域的研究主要集中在珊瑚礁[8–9]及鱼类[10–11]等方面,浮游植物与生态环境关系的研究较少,西沙群岛海域终年存在温跃层,真光层的营养盐被浮游生物消耗后,下层营养盐难以补充至上层,导致表层海水氮、磷营养盐浓度经常处于胁迫状态,从而抑制了浮游植物的生长,使得海区表层海水初级生产力维持在较低的水平[12–13]。陈露等[13]在2014年秋季在西沙海域进行营养盐加富研究。吴钟解等[14]于2007年调查西沙生态监控区浮游植物现状,共发现4门50属117种浮游植物。关于西沙宣德群岛浮游植物群落结构及环境因子关系的研究报道甚少,因此, 本研究以西沙宣德群岛附近海域为调查区域,对浮游植物种类、群落结构、优势种、多样性特征及相关环境因子进行调查、分析,以期为该区域生态结构与功能的深入研究及生态系统保护提供基础资料。
1 材料和方法
1.1 采样位置与采样时间
于2019年2月(冬季)和11月(秋季)分别对西沙宣德群岛附近海域进行浮游植物及相关环境因子调查,共布设10个站位(图1)。
1.2 样品采集及分析
依据海洋监测规范(GB/T 17378—2007)[15],用浅水Ⅲ型浮游生物网垂直拖网获得样品,所有浮游植物样品用饱和鲁哥试剂固定,静止浓缩后用ZEISS显微镜观察和计数。浮游植物鉴定参考《中国海藻志》第5卷(硅藻门)第1册(中心纲)[16]、《中国海域常见浮游硅藻图谱》[17]、《中国海洋浮游硅藻类》[18]、[19]等图谱资料。现场仪器测定水深、水温等。盐度、溶解氧和营养盐等化学数据,通过采集表层水样(0.5 m)进行测定[15]。

图1 调查站位(S1~S10)示意图
1.3 数据处理和分析
计算各站位的浮游植物群落多样性指数()、丰富度指数()和均匀度指数(),比较研究区域的浮游植物群落结构。采用CANOCO 4.5软件对浮游植物物种数据和环境因子进行多元统计分析。使用SPSS 20.0软件进行Tukey显著差异法和相关性分析。

优势种是由优势度[23]决定的,=P×f, 式中,f为第种物种在各个站位出现的频率;P为第种物种占各站位物种总数的比例,以>0.02作为优势种。
选取≥0.01且在各站位的出现频率≥50%的浮游植物,对物种数据矩阵进行log(+1)转换后, 应用CANOCO 4.5软件对研究海域浮游植物物种数据和环境因子进行了多元统计分析。将物种数据进行去趋势对应分析(detrended correspondence analysis,DCA)表明4个排序轴中梯度最大值<3,故选用冗余分析(redundancy analysis, RDA)线性模型进行排序分析。
2 结果和分析
2.1 种类组成及分布
宣德群岛附近海域冬季和秋季2个航次调查共采集到109种浮游植物,隶属于3门45属,其中硅藻种类最多,34属81种,占总种类数的74.31%;甲藻8属24种,占总种类数的20.02%;蓝藻3属4种,占总种类数的3.67%。其中,以硅藻的角毛藻属为优势类群,共13种,占总种类数的16.88%。冬季调查共发现浮游植物77种,硅藻57种,占冬季总种类数74.03%;甲藻16种,占冬季总种类数20.78%;蓝藻4种,占冬季总种类数5.19%。秋季调查共发现浮游植物79种,硅藻63种,占秋季总种类数79.75%;甲藻13种,占秋季总种类数16.45%;蓝藻3种,占秋季总种类数3.80%。冬季S6和S9站位浮游动物种类最多,为30种,S1最少,为7种;秋季S3站位浮游动物种类最多,为29种,S4站位最少,为12种(图2)。

图2浮游植物物种数
冬季,浮游植物细胞丰度为(0.04~21.06)× 107cells/L,平均为(5.27±6.14)×107cells/L。S10站位浮游植物丰度最高,为21.06×107cells/L,其中, 铁氏束毛藻()大量聚集,成为调查区域的优势种之一;S4站位浮游植物丰度最低, 为0.04×107cells/L。秋季,浮游植物细胞丰度为(0.29~4.86)×105cells/L, 平均为(1.56±1.40)×105cells/L;浮游植物丰度最高站位同样出现在S10, 为4.86× 105cells/L;浮游植物丰度最低出现在S6站位,为0.29×105cells/L (图3)。
2.2 优势种与群落结构指数评价
冬季调查共得到优势种7种,分别属于硅藻、甲藻和蓝藻(表1)。其中,蓝藻优势最为明显,铁氏束毛藻为第1优势种(0.203),平均丰度为2.18×107cells/L, 占总丰度的28.93%;红海束毛藻()为第2优势种(0.108), 平均丰度为0.89×107cells/L,占总丰度的13.56%。秋季调查的优势种均属硅藻类,菱形海线藻()具有绝对优势(0.463),平均丰度为0.72×105cells/L,占总丰度的46.29%。冬季的优势种标志星杆藻()、中沙角管藻()、念珠梯楔形藻()、长菱形藻()、二齿双管藻()、红海束毛藻和铁氏束毛藻在秋季均有出现,但未成为秋季的优势种。

图3 浮游植物丰度

表1 冬秋季浮游植物的优势种组成
: 优势度指数;: 平均细胞丰度;: 优势种出现频率。
: Dominance index;: Mean cell abundance;: Frequency of dominant species.
两个航次浮游植物的和相似,冬季浮游植物的和分别为1.94~3.95和0.40~0.86,平均为3.15±0.64和0.71±0.13; 秋季分别为1.62~4.09和0.36~0.93,平均为3.28±0.72和0.75±0.17 (表2)。除冬季6号站位和秋季10号站位浮游植物的和处于较低水平外,其他站位浮游植物物种分布较为均匀。两个季节浮游植物的种类丰富度指数()存在显著差异(<0.05),冬季浮游植物为0.47~1.82,平均为1.36±0.44;秋季为1.00~2.25,平均为1.77± 0.33,说明秋季浮游植物物种数分布比冬季均匀。
2.3 环境特征
从表3可见,宣德群岛附近海域冬季和秋季的盐度、叶绿素和硝酸盐存在极显著差异(<0.01), 铵盐存在显著差异(<0.05),其他环境因素冬季和秋季没有显著变化。两个季节无机态氮磷比存在极显著差异,冬季无机态氮磷比(164.30±87.81)高于秋季(32.92±27.47),且冬季所有站位的无机态氮磷比均大于32,为78.01~370.17。
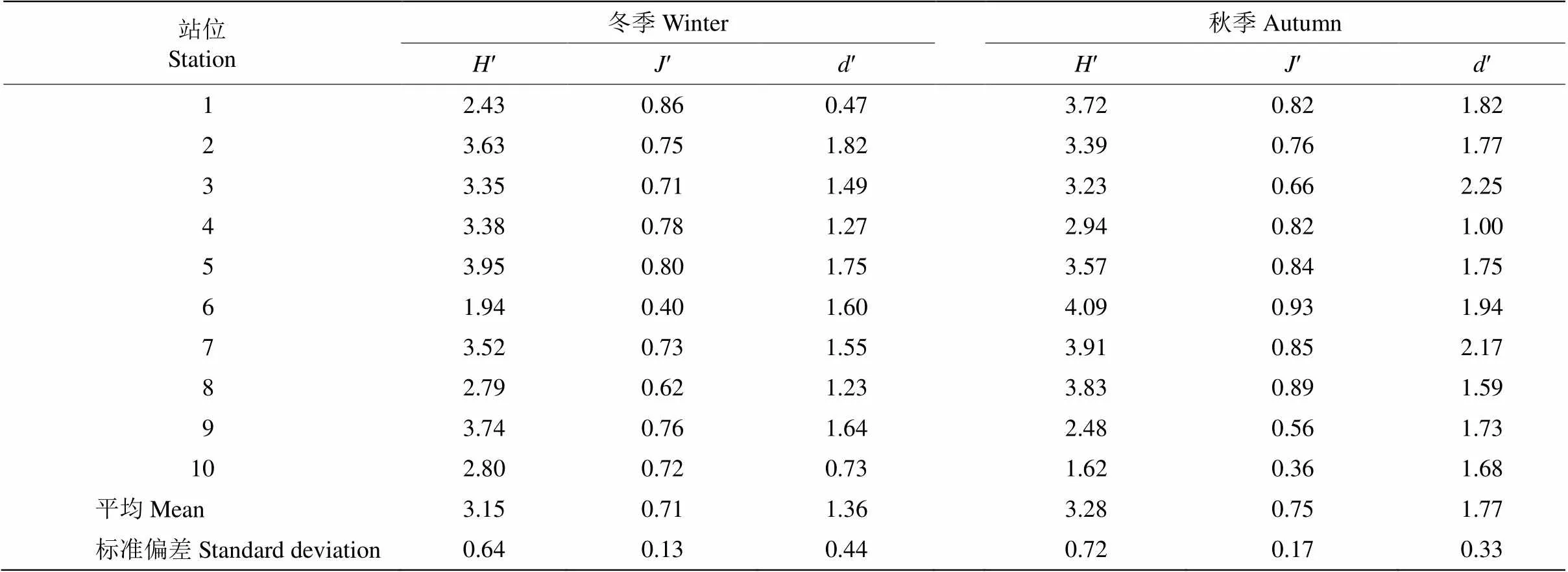
表2 冬秋季浮游植物群落参数分析
: 多样性指数: 均匀度指数;: 丰富度指数。
: Shannon-Wiener diversity index;: Pielou evenness index;: Margalef richness index.
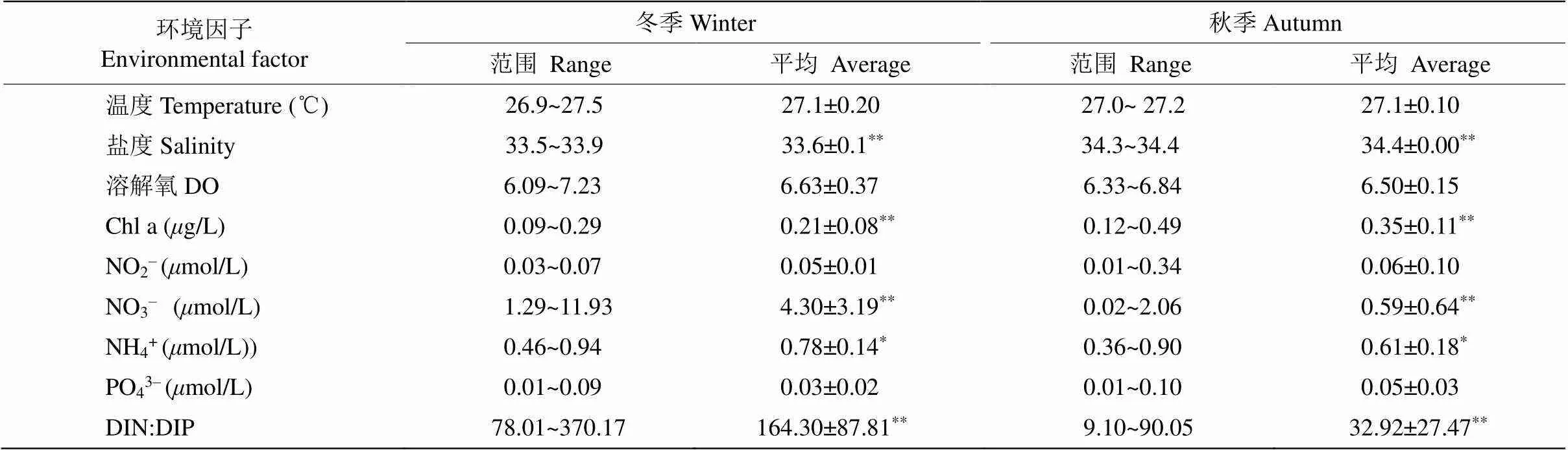
表3 冬秋季表层环境因子的变化
**:<0.01; *:<0.05
2.4 浮游植物群落与环境因子的关系
将冬季和秋季两个航次浮游植物物种与8个环境因子进行RDA排序分析,结果表明(图4):冬季RDA排序图中,8个共解释了浮游植物总变量的70.44%,前两个排序轴的特征值分别为0.291和0.187,分别解释了29.14%和47.87%的物种变量; 秋季RDA排序图中,8个共解释了浮游植物总变量的73.22%,前两个排序轴的特征值分别为0.340和0.234,分别解释了33.96%和57.38%的物种变量,两个航次环境因子和物种排序轴间的相关系数均大于0.95, 较好地反映了物种与环境之间的关系。
利用向前引入法逐步筛选环境因子,在冬季航次调查结果中,8个环境因子的检验结果均未达到显著水平;秋季航次中有两个环境因子达到显著水平,分别是亚硝酸盐(=1.9,=0.018)和盐度(=2.4,=0.010),这说明冬季浮游植物群落结构不受单一环境因子影响,亚硝酸盐和盐度是影响秋季浮游植物群落结构的主要环境因子。
浮游植物和环境因子的RDA排序图表明(图4),冬季浮游植物主要集中在第一和第三象限,与环境因子有明显关系,可以分为三类:第一类包括铁氏束毛藻、粗股角毛藻、长菱形藻、秘鲁角毛藻、尖刺伪菱形藻、笔尖形根管藻与硝酸盐和N/P呈正相关,与氨盐呈负相关;第二类包括细长翼根管藻、劳氏角毛藻、二齿双管藻、标志星杆藻、念珠梯楔形藻、中沙角管藻与无机磷酸盐和DO呈负相关; 第三类包括日本角毛藻、日本船藻、菱形藻、红海束毛藻与温度和盐度呈正相关,与无机磷酸盐和DO呈负相关。秋季浮游植物与环境因子的关系出现明显变化,翼根管藻、金色角毛藻、角毛藻与亚硝酸盐呈正相关,与硝酸盐、无机磷酸盐、DO和N/P呈负相关;劳氏角毛藻、尖刺拟菱形藻、柔弱拟菱形藻、菱形海线藻、大洋角管藻与硝酸盐、温度和盐度呈负相关;丹麦细柱藻、翼根管藻、舟形藻与硝酸盐、无机磷酸盐、DO和N/P呈正相关, 与亚硝酸盐呈负相关。

图4 冬季和秋季主要浮游植物种类与环境因子的RDA排序图。T: 温度; S: 盐度; DO: 溶解氧; NO2-N: 亚硝酸盐氮; NO3-N: 硝酸盐氮; NH3-N: 氨氮; PO4-P: 无机磷酸盐; N/P: 无机氮磷比。An: 标志星杆藻; Cz: 中沙角管藻; Cs: 角毛藻; Cf: 粗股角毛藻; Cl: 劳氏角毛藻; Cn: 日本角毛藻; Cp: 秘鲁角毛藻; Ca: 金色角毛藻; Cm: 念珠梯楔形藻; Ij: 日本船藻; Nis: 菱形藻; Nl: 长菱形藻; Pp: 尖刺伪菱形藻; Pd: 柔弱拟菱形藻; Rla: 细长翼根管藻;Rs: 笔尖形根管藻; Ra: 翼根管藻; Rc: 距端根管藻; Ab: 二齿双管藻; La: 巨大鞘丝藻; Te: 红海束毛藻; Tt: 铁氏束毛藻; Cpe: 大洋角管藻; Ld: 丹麦细柱藻; Nas: 舟形藻; Nc: 新月菱形藻; Tn: 菱形海线藻。
3 结论和讨论
3.1 浮游植物群落组成及变化
本次调查冬季和秋季航次共发现浮游植物109种,两季皆以硅藻为主,其次为甲藻和蓝藻,这与吴钟解等[14]在西沙珊瑚礁监控区的浮游植物生态现状调查研究结果相符。冬季浮游植物细胞平均丰度(5.27±6.14)×107cells/L显著高于秋季浮游植物细胞平均丰度(1.56±1.40)×105cells/L (<0.01),2个季节优势种差异较大,冬季铁氏束毛藻和红海束毛藻分布广泛,秋季菱形海线藻占主导地位。由于西沙海域浮游植物群落调查研究较少,目前只看到吴钟解等[16]在西沙珊瑚礁监控区的浮游植物调查结果, 其平均细胞丰度为2.08×107cells/L,两者调查虽时间不同,但地理位置相似,与本次调查数量级相当。西沙和南沙海域特征具有相似性,前人[24–25]调查南沙海域的浮游植物数量级为104cells/m3(换算后为107cells/L),与本次调查数量级也相当。本调查冬秋季结果差异较大,可能与季节有关,李红飞等[3]总结了1998年以来南海浮游植物群落调查,认为南海浮游植物的物种组成和丰度分布具有明显的季节性,冬季明显高于夏季,春秋两季略低,主要优势物种为硅藻,其后为甲藻、蓝藻。
可以反映群落的复杂度,浮游植物种类数越多或分布越均匀,越大[26]。同时又可作为生态环境质量评估的依据[27],当≥3时,生态环境优良;当2≤<3时,生态环境一般;当1≤<2时,生态环境差。在实践应用中,通常将0.3作为判定浮游植物群落结构稳定的阈值[28]。而且浮游植物和及群落结构的稳定性呈正相关,所以越大,群落结构越稳定,生态系统抗干扰能力越强[29]。本研究表明,冬季各站位浮游植物群落的和平均分别为3.15和0.71,秋季的分别为3.28、0.75,说明该海域浮游植物多样性较高,物种较为丰富,群落结构稳定性较好,生态系统具有较强的抗干扰能力。
3.2 影响浮游植物群落的环境因子
温度、盐度、营养盐和水动力等环境因子及浮游动物摄食等生态学过程均是影响浮游植物生长的主要因素。西沙海域面积广阔,理化条件复杂, 形成了千差万别的水文环境。影响浮游植物群落结构的既有营养盐分布等影响较大的因素,也有盐度、温度、溶解氧等影响微弱的因素[3]。在本研究中, RDA结果表明冬季浮游植物主要受硝酸盐、氮磷比、铵盐、温度及盐度的影响,秋季主要受亚硝酸盐、温度和盐度的影响。
海水中的营养盐结构对浮游植物的生长至关重要,浮游植物对营养盐的利用有特定比例和极限浓度[30–31]。目前公认的有Justić等[32]提出的绝对限制法,从浮游植物生长的最低阈值来衡量营养盐对浮游植物生长的影响,当海水中硅浓度<2mol/L、无机氮浓度<1mol/L、无机磷<0.1mol/L时,判定该元素是唯一的限制因子。宣德群岛海域两个季节无机磷浓度较低,所有站位均低于0.1mol/L, 秋季部分站位无机氮浓度较低(<1mol/L),平均浓度为(1.26±0.70)mol/L,所以宣德群岛海域为磷限制海域,秋季部分区域为氮磷双限制海域。该结果与吴钟解等[14]调查结果一致,同时也再次印证了宣德群岛海域为贫营养海域[33]。该海域冬季浮游植物丰度高,可能是因为冬季宣德群岛盛行东北季风,使表层海水与中层海水搅动形成混合层,同时伴随上升流的形成对海水表层的营养盐起到了一定的补充作用,进而促进浮游植物的生长[34]。不断上涌的上升流将低温而富营养盐的海水带到表层,提高了局部海域硝酸盐、亚硝酸盐等营养盐浓度[35],促进浮游植物的生长[36],上升流中的主要优势种是硅藻,其次为甲藻和蓝藻。氮磷比随硅藻的生长而升高,随甲藻的生长而下降[37]。调查区域秋冬氮磷比一直处于较高水平,是硅藻大量生长成为绝对优势种类群的结果。
温度对浮游植物的光合作用和呼吸作用都有重要影响,其通常通过影响细胞代谢相关酶的效率而影响浮游植物光合作用和呼吸作用的效率[38]。冬季航次中蓝藻细胞丰度占比高达48%,成为主导优势种,这与戴明等[25]在2013年调查南沙群岛的结果相似,这是否与季节有关值得进一步研究。冬季航次中的蓝藻主要种类为铁氏束毛藻和红海束毛藻。研究表明,束毛藻是广泛分布于热带、亚热带寡营养盐海域中的固氮生物,他们可以将大气中的N2转换为浮游植物所需的氨氮,是寡营养盐海域中新氮的重要来源和新生产力的主要贡献者[25,39–40]。温度是影响束毛藻分布的重要因素,可能该海域冬季温度更适宜束毛藻生长。
综上,本次调查研究发现宣德群岛附近海域浮游植物多样性较高,物种较为丰富,群落结构稳定性较好,生态系统具有较强的抗干扰能力。该海域浮游植物丰度具有明显的季节性,冬季浮游植物丰度显著高于秋季。宣德群岛附近海域浮游植物主要受亚硝酸盐、温度及盐度等多种环境因子共同影响。
[1] FALKOWSKI P G, BARBER R T, SMETACEK V. Biogeochemical controls and feedbacks on ocean primary production [J]. Science, 1998, 281(5374): 200–206. doi: 10.1126/science.281.5374.200.
[2] MA W, SUN J, XUE B, et al. Phytoplankton community of northern South China Sea in autumn of 2010 [J]. Acta Ocean Sin, 2016, 38(4): 43–53. [马威, 孙军, 薛冰, 等. 2010年秋季南海北部浮游植物群落结构研究 [J]. 海洋学报, 2016, 38(4): 43–53. doi: 10.3969/j.issn. 0253-4193.2016.04.004.
[3] LI H F, LIN S J. Research progresses of phytoplankton ecology in the South China Sea [J]. J Xiamen Univ (Nat Sci), 2019, 58(1): 1–10. [李红飞, 林森杰. 南海浮游植物生态学研究进展 [J]. 厦门大学学报(自然科学版), 2019, 58(1): 1–10. doi: 10.6043/j.issn.0438-0479.2018 05038.]
[4] NING X, CHAI F, XUE H, et al. Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea [J]. J Geophys Res Oceans, 2004, 109(C10): C10005. doi: 10.1029/2004JC002365.
[5] SUIKKANEN S, LAAMANEN M, HUTTUNEN M. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea [J]. Estuar Coast Shelf Sci, 2007, 71(3/4): 580–592. doi: 10.1016/j.ecss.2006.09.004.
[6] LE F F, SUN J, NING X R, et al. Phytoplankton in the northern South China Sea in summer 2004 [J]. Oceanol Limnol Sin, 2006, 37(3): 238– 248. [乐凤凤, 孙军, 宁修仁, 等. 2004年夏季中国南海北部的浮游植物 [J]. 海洋与湖沼, 2006, 37(3): 238–248. doi: 10.3321/j.issn: 0029-814X.2006.03.008]
[7] ZHANG Z D, SHAO K S, YANG Z X, et al. Evaluation of the Xisha coral reef ecosystem carrying capacity [J]. Mar Environ Sci, 2018, 37(4): 487–492. [张振冬, 邵魁双, 杨正先, 等. 西沙珊瑚礁生态承载状况评价研究 [J]. 海洋环境科学, 2018, 37(4): 487–492. doi: 10. 13634/j.cnki.mes.2018.04.003.]
[8] HUANG H, DONG Z J, LIAN J S. Establishment of nature reserve of coral reef ecosystem on the Xisha Islands [J]. Trop Geogr, 2008, 28(6): 540–544. [黄晖, 董志军, 练健生. 论西沙群岛珊瑚礁生态系统自然保护区的建立 [J]. 热带地理, 2008, 28(6): 540–544. doi: 10.3969/j.issn. 1001-5221.2008.06.010.
[9] HUANG H, YOU F, LIAN J S, et al. Species diversity and distribution of scleractinian coral at Xisha Islands, China [J]. Biodiv Sci, 2011, 19(6): 710–715. [黄晖, 尤丰, 练健生, 等. 西沙群岛海域造礁石珊瑚物种多样性与分布特点 [J]. 生物多样性, 2011, 19(6): 710–715. doi: 10.3724/SP.J. 1003.2011.06132.]
[10] LI Y C, WU Z J, CHEN S Q, et al. Discussion of the diversity of the coral reef fish in the shallow reefs along the Yongxing and Qilianyu Island [J]. Mar Environ Sci, 2017, 36(4): 509–516. [李元超, 吴钟解, 陈石泉, 等. 永兴岛及七连屿浅水礁区珊瑚礁鱼类多样性探讨 [J]. 海洋环境科学, 2017, 36(4): 509–516. doi: 10. 13634/j.cnki.mes.2017. 04.005.]
[11] YANG W D, HU J T, LIN B A, et al. Species diversity of coral reef fishes in Zhaoshu Island Waters, Xisha Islands [J]. J Xiamen Univ (Nat Sci), 2018, 57(6): 819–826. [杨位迪, 胡俊彤, 林柏岸, 等. 西沙赵述岛海域珊瑚礁鱼类物种多样性 [J]. 厦门大学学报(自然科学版), 2018, 57(6): 819–826. doi: 10.6043/j.issn.0438-0479.201807002.]
[12] LIN H Y, HAN W Y. Studies on the restriction of phytoplankton growth in Waters of Nansha Islands [C]// The 7th National Symposium on Oceanology and Limnology for Young Scholars Abstract. Qingdao: Chinese Society of Oceanology and Limnology, 2001: 14. [林洪瑛, 韩舞鹰. 南沙群岛海域营养盐对浮游植物生长的限制研究 [C]// 第七届全国海洋湖沼青年学者学术研讨会论文摘要集. 青岛: 中国海洋湖沼学会, 2001: 14.]
[13] CHEN L, LI C H, DAI M, et al. Effect of nutrient enrichment on phytoplankton growth in the adjacent waters of the Xisha Yongxing Islands in late fall [J]. S China Fish Sci, 2016, 12(4): 125–130. [陈露, 李纯厚, 戴明, 等. 西沙永兴岛附近海域秋末氮磷营养盐加富对浮游植物生长限制的影响 [J]. 南方水产科学, 2016, 12(4): 125–130. doi: 10.3969/j.issn.2095-0780.2016.04.016.]
[14] WU Z J, WANG D R, LI Y C, et al. The ecological status of phyto- plankton in monitored Xisha areas [J]. Chin J Trop Crops, 2010, 31(6): 1020–1025. [吴钟解, 王道儒, 李元超, 等. 西沙监控区浮游植物生态现状 [J]. 热带作物学报, 2010, 31(6): 1020–1025. doi: 10.3969/j.issn. 1000-2561. 2010.06.027.]
[15] General Administration of Quality Supervision, Inspection and Quaran- tine of the People’s Republic of China, Standardization Administration of the People’s Republic of China. GB 17378.4–2007 The specification for marine monitoring [S]. Beijing: Standards Press of China, 2008. [中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378.4–2007海洋监测规范[S]. 北京: 中国标准出版社, 2008.]
[16] GUO Y J. Flora Algarum Marinarum Sinicarum, Tomus V. Bacilla- riophyta, No. 1 Centricae [M]. Beijing: Science Press, 2003: 1–493. [郭玉洁. 中国海藻志第5卷(硅藻门)第1册(中心纲) [M]. 北京:科学出版社, 2003: 1–493.]
[17] YANG S M, DONG S G. Chromatograms of Common Marine Plank- tonic Diatoms in China [M]. Qingdao: China Ocean University Press, 2006: 1–267. [杨世民, 董树刚. 中国海域常见浮游硅藻图谱 [M]. 青岛: 中国海洋大学出版社, 2006: 1–267.]
[18] JIN D X, CHEN J H, HUANG K G. Planktonic Diatoms from China Sea [M]. Shanghai: Shanghai Science and Technology Press, 1965: 1–230. [金德祥, 陈金环, 黄凯歌. 中国海洋浮游硅藻类 [M]. 上海: 上海科学技术出版社, 1965: 1–230.]
[19] OMURA T, IWATAKI M, VALERIANO B, et al. Marine Phyto- plankton of the Western Pacific [M]. Tokyo: Kouseisha Kouseikaku Co., Ltd., 2012.
[20] SHANNON C E, WEAVER W. The Mathematical Theory of Commu- nication [M]. Urbana: University of Illinois Press, 1949: 1–125.
[21] MARGALEF R. Perspectives in Ecological Theory [M]. Chicago: University of Chicago Press, 1968: 1–111.
[22] PIELOU E C. An Introduction to Mathematical Ecology [M]. New York: Wiley-Interscience, 1969:1-294.
[23] HE Q, SUN J. The Netz-phytoplankton community in Changjiang (Yangtze) River Estuary and adjacent waters [J]. Acta Ecol Sin, 2009, 29(7): 3928–3938. [何青, 孙军. 长江口及其邻近水域网采浮游植物群落 [J]. 生态学报, 2009, 29(7): 3928–3938. doi: 10.3321/j.issn:1000- 0933.2009.07.056.]
[24] SONG X Y, HUANG L M, QIAN S B, et al. Phytoplankton diversity in waters around Nansha Islands in spring and summer [J]. Biodiv Sci, 2002, 10(3): 258–268. [宋星宇, 黄良民, 钱树本, 等. 南沙群岛邻近海区春夏季浮游植物多样性研究 [J]. 生物多样性, 2002, 10(3): 258–268. doi: 10.3321/j.issn:1005-0094.2002.03.002.]
[25] DAI M, LIU H X, LIAO X L, et al. Characteristics of net phyto- plankton community and their relationships to environmental factors in the waters around Nansha Islands [J]. Chin J App Ecol, 2013, 24(12): 3553–3560. [戴明, 刘华雪, 廖秀丽, 等. 初秋南沙群岛海域网采浮游植物群落特征及其与环境因子的关系 [J]. 应用生态学报, 2013, 24(12): 3553–3560. doi: 10.13287/j.1001-9332.2013.0593.]
[26] GUO P Y, SHEN H T, LIU A C, et al. The species composition, community structure and diversity of zooplankton in Changjiang Estuary [J]. Acta Ecol Sin, 2003, 23(5): 892–900. [郭沛涌, 沈焕庭, 刘阿成, 等. 长江河口浮游动物的种类组成、群落结构及多样性[J].生态学报, 2003, 23(5): 892–900. doi: 10.3321/j.issn:1000-0933.2003. 05.008.]
[27] General Administration of Quality Supervision, Inspection and Quaran- tine of the People’s Republic of China, Standardization Administration of the People’s Republic of China. GB 17378.7–2007 The specification for marine monitoring: Ecological survey for offshore pollution and biological monitoring [S]. Beijing: Standards Press of China, 2008. [中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378.7–2007海洋监测规范近海污染生态调查和生物监测 [S]. 北京: 中国标准出版社, 2008.]
[28] WANG D, CHEN P M, LU J J, et al. Annual changes of phyto- plankton’s ecological features in Qinzhou Bay of South China [J]. Chin J App Ecol, 2013, 24(6): 1686–1692. [王迪, 陈丕茂, 逯晶晶, 等. 钦州湾浮游植物周年生态特征[J]. 应用生态学报, 2013, 24(6): 1686– 1692. doi: 10.13287/j.1001-9332.2013.0360.]
[29] LI K Z, GUO Y J, YIN J Q, et al. Phytoplankton diversity and abundance in Nansha Islands waters in autumn of 1997 [J]. J Trop Oceanogr, 2005, 24(3): 25–30. [李开枝, 郭玉洁, 尹健强, 等. 南沙群岛海区秋季浮游植物物种多样性及数量变化[J]. 热带海洋学报, 2005, 24(3): 25–30. doi: 10. 3969/j.issn.1009-5470.2005.03.004.]
[30] SHEN Z L. Long-term changes in nutrient structure and its influences on ecology and environment in Jiaozhou Bay [J]. Oceanol Limnol Sin, 2002, 33(3): 322–331. [沈志良. 胶州湾营养盐结构的长期变化及其对生态环境的影响 [J]. 海洋与湖沼, 2002, 33(3): 322–331. doi: 10. 3321/j.issn:0029-814X.2002.03.013.]
[31] SONG X X, YU Z M, YIN K D, et al. Temporal and spatial distribution of nutrients and Chl a in the coastal area of Hong Kong [J]. Oceanol Limnol Sin, 2013, 44(4): 846–852. [宋秀贤, 俞志明, 殷克东, 等. 香港近岸海域营养盐结构特征及其对浮游植物生长的影响 [J]. 海洋与湖沼, 2013, 44(4): 846–852.]
[32] JUSTĆ D, RABALAIS N N, TURNER R E, et al. Changes in nutrient structure of river-dominated coastal waters: Stoichiometric nutrient balance and its consequences [J]. Estuar Coast Shelf Sci, 1995, 40(3): 339–356. doi: 10.1016/S0272-7714(05)80014-9.
[33] CAI W J, DAI M H, WANG Y C, et al. The biogeochemistry of inorganic carbon and nutrients in the Pearl River estuary and the adjacent northern South China Sea [J]. Cont Shelf Res, 2004, 24(12): 1301–1319. doi: 10.1016/j.csr.2004.04.005.
[34] LIU J, LI L, HE J. Recent advances on characterization of the phyto- planktonic distribution in the South China Sea [J]. Mar Geol Quat Geol, 2010, 30(3): 133–142. [刘杰, 李丽, 贺娟. 南海海域浮游植物分布特征及研究进展 [J]. 海洋地质与第四纪地质, 2010, 30(3): 133–142. doi: 10.3724/SP.J.1140.2010.03133.]
[35] HUANG B Q, XIANG W G, ZENG X B, et al. Phytoplankton growth and microzooplankton grazing in a subtropical coastal upwelling system in the Taiwan Strait [J]. Cont Shelf Res, 2011, 31(S6): S48–S56. doi: 10.1016/j.csr.2011.02.005.
[36] BARTH J A, MENGE B A, LUBCHENCO J, et al. Delayed upwelling alters nearshore coastal ocean ecosystems in the northern California current [J]. Proc Natl Acad Sci USA, 2007, 104(10): 3719–3724. doi: 10.1073/pnas.0700462104.
[37] LI J. The research of nutrient structure and its function on the succession of phytoplankton predominant species in the high frequent harmful algae blooms occurrence areas in East China Sea [D]. Qingdao: Ocean University of China, 2008. [李京. 东海赤潮高发区营养盐结构及对浮游植物优势种演替的作用研究 [D]. 青岛: 中国海洋大学, 2008.]
[38] CHEN Y L, ZHAO D Z, YANG J H, et al. The quantitative study on ecological amplitude of sea temperature of algae [J]. Acta Oceanol Sin, 2009, 31(5): 156–161. [陈艳拢, 赵冬至, 杨建洪, 等. 赤潮藻类温度生态幅的定量表达模型研究[J]. 海洋学报, 2009, 31(5): 156–161.]
[39] JENKINS W J. Nitrate flux into the euphotic zone near Bermuda [J]. Nature, 1988, 331(6156): 521–523. doi: 10.1038/331521a0.
[40] CAPONE D G, ZEHR J P, PAERL H W, et al., a globally significant marine cyanobacterium [J]. Science, 1997, 276 (5316): 1221–1229. doi: 10.1126/science.276.5316.1221.
Phytoplankton Community and Its Influencing Factors in Xuande Islands of Xisha Sea Area
LIU Xiaohong1, SHI Jun2
(1.Sansha Marine Environmental Monitoring Central Station, State Oceanic Administration,Haikou 570311, China; 2. Haikou Marine Environmental Monitoring Central Station, State Oceanic Administration,Haikou 570311, China)
In order to understand the characteristics of phytoplankton community structure in Xisha Xuande Islands, the phytoplankton community composition and environmental factors sampled in February and November 2019 were analyzed. The results showed that 109 species of phytoplankton were found during the two voyages, among which Bacillariophyta were the most with 81 species.The dominant species varied between winter and autumn,,,, etc. were the dominant in winter, and did,andwere in autumn. The average abundance of phytoplankton in winter [(5.27±6.14)×107cells/L] was significantly higher than that in autumn [(1.56±1.40)×105cells/L]. The structure of phytoplankton community was mainly affected by nitrite, salinity and other environmental factors. The average of Shannon-Wiener diversity index (), Pielou evenness index () and Margalef richness index () inwinter and autumn were 3.15, 0.71, 1.36 and 3.28, 0.75, 1.77, respectively. As a whole, these indicated that phytoplankton community in Xuande Islands had high diversity and rich species, as well as complex and stable community structure in winter and autumn.
Xuande Islands of Xisha; Phytoplankton; Environmental factor; Community structure
10.11926/jtsb.4572
2021-11-17
2022-03-23
国家海洋局南海分局局长基金项目(180239);海南省自然科学基金项目(419QN381)资助
The work was supported by the Director Project of South China Sea Branch of State Oceanic Administration (Grant No. 180239), and the Project for Natural Science in Hainan (Grant No. 419QN381).
刘晓红(1990年生),女,工程师,主要从事海洋生态调查和海洋生态修复工作。E-mail: 1215893627@qq.com
猜你喜欢
大自然探索(2023年7期)2023-11-14
课堂内外(小学版)(2023年9期)2023-10-11
潍坊学院学报(2020年2期)2021-01-18
艺术品鉴(2020年10期)2020-11-27
黑龙江水利科技(2020年8期)2020-01-12
流行色(2019年10期)2019-12-06
艺术品鉴(2019年7期)2019-09-02
上海建材(2018年1期)2018-04-18
公关世界(2017年8期)2017-05-30
宝藏(2017年2期)2017-03-20
