
短期增温对亚热带常绿阔叶林林下植被物种多样性的影响
2023-04-11 03:27籍烨陈仕东熊德成胥超刘小飞何宗明杨智杰
热带亚热带植物学报 2023年2期
籍烨, 陈仕东, 熊德成, 胥超, 刘小飞, 何宗明, 杨智杰*
短期增温对亚热带常绿阔叶林林下植被物种多样性的影响
籍烨1,2, 陈仕东1,2, 熊德成1,2, 胥超1,2, 刘小飞1,2, 何宗明3, 杨智杰1,2*
(1. 福建师范大学地理科学学院,福州 350007;2. 福建三明森林生态系统国家野外科学观测研究站,福建 三明 365000;3. 福建农林大学林学院, 福州 350002 )
为了解气候变暖对亚热带常绿阔叶林林下植被物种多样性的影响,采用土壤增温方式,研究短期4年增温(4 ℃)对亚热带常绿阔叶天然林林下植被物种多样性的影响。结果表明,短期增温对林下植被物种组成影响不显著(>0.05),增温和对照样地林下植被共有植物38科59属77种,其中增温样地37科53属65种,对照样地36科52属63种。短期增温使乔木盖度增加了22.61%,草本和灌木盖度分别降低4.97%和21.75%;增温使草本、灌木和乔木的高度分别降低21.64%、3.37%和5.59%。草本植物中的蕨类重要值排名在增温后呈下降趋势,乔木重要值呈上升趋势(>0.05)。虽然植物物种多样性指数在增温后差异不显著(>0.05),但随温度增加均呈递减的趋势。因此,林下植被物种组成对短期增温响应不敏感,增温使草本植物中的蕨类重要值下降,对植物多样性指数产生一定负面影响,但这种响应并不敏感,预计长期增温将可能导致整个群落从草本向灌木和乔木方向演替。
林下植被; 物种多样性; 土壤增温;亚热带森林
气候变暖已成为不争的事实[1],IPCC (联合国政府间气候变化专门委员会, Intergovernmental Panel on Climate Change)第六次评估报告显示,预计未来20年全球平均升温将达到或超过1.5 ℃[2], 由温室效应的不断累积所引起的气候变暖将直接影响植物群落组成[3]、生长特性[4]及森林生产力[5],导致生态系统的稳定性和持续性发生改变[6],对陆地生态系统的结构和功能产生进一步影响[7]。
了解植物多样性应对气候变化的响应是生态学研究的重要课题[8]。森林生态系统是陆地生态系统的重要组成部分,其物种丰富度会因气候变暖而存在差异[9]。欧洲北部温带山区森林在变暖条件下的耐寒树种损失很少[10]。对青藏高原和四川盆地间的过渡山区107个长期监测样地进行重新调查,记录了所有树种40年间的变化,结果表明,气候变暖使森林树种丰富度显著增加[11]。西欧树种会以每10年29 m的速度向高海拔梯度显著上升[12]。但这些研究多以高大的乔木树种为研究对象,极易忽略乔木层下方的林下植被。
林下植被是森林生态系统的重要组成部分,其物种多样性和群落特征是衡量森林质量变化的重要指标,丰富的林下植被种类不仅是优化森林生态系统功能的基础,而且在促进土壤养分循环、提高水土保持能力、为上层乔木提供适宜环境发挥着重要功能[13]。因此,维持林下草本和灌木的生物多样性对于提高整个森林生态系统的整体功能起着至关重要的作用[14]。但目前由于研究的缺乏,林下植被多样性对气候变化响应仍然相对未知[15]。
热带亚热带低纬度地区植物对热适应的生态位相对较窄,其适应生态位温度范围以外温度的能力较弱[16],这可能会产生比高纬度地区更强的生物效应[17]。气候变暖会导致陆地生态系统的降雨格局发生改变,使纬向平均降水很可能在高纬度和部分中纬度地区增加,而在亚热带地区则会减少[18]。有研究推测,到2100年,热带低纬度地区植物物种丰富度会因温度和降水格局的变化而发生下降[19]。但这种推测可能具有很大的不确定性,增温控制试验将这种推测的不确定性变为可能,可以揭示出生态系统对气候变化的响应[20–21]。
本文以中亚热带常绿阔叶天然林野外原位增温样地为研究对象,采用电缆增温技术(增温4 ℃),4年后调查增温林下植被多样性的变化,研究增温对亚热带常绿阔叶天然林林下植被多样性的影响, 为亚热带气候变暖条件下林下植被多样性的保护提供基础数据和理论依据。
1 材料和方法
1.1 研究区概况
试验样地位于福建三明森林生态系统国家野外科学观测研究站的格氏栲天然林增温平台内(26° 9′~26°12′ N,117°27′~117°29′ E)。以低山丘陵地貌为主,海拔250~500 m,属中亚热带季风气候,年均温19.6 ℃,年均降水量1 700 mm,相对湿度81%。研究区内主要以花岗岩发育的黄壤和红壤为主[22]。
1.2 增温样地设置
2014年10月在格氏栲天然林随机设置10个6 m×6 m的试验样地,设对照(不增温)和增温(+4 ℃)处理,每处理5个重复。在每个样地布设6个温度传感器(德国JUMO)和6个水分传感器(美国Decagon),其中3个温度传感器埋在10 cm处, 用于监测和控制土壤增温,另外3个温度传感器分别监测10、20和40 cm土壤温度,水分传感器测定土壤10、20、40 cm含水量。在增温样地中布设增温电缆,电缆埋在土壤10 cm深处,间距20 cm, 温度和湿度传感器均布置在两条电缆线中间位置。土壤增温采用半自主研发的土壤主动增温控制系统(哈佛森林增温系统升级版),增温样地比对照样地温度高4 ℃,土壤温度和含水量数据每30 min储存1次。样地于2015年10月开始增温[23],至2019年调查时增温已满4年。2019年的土壤增温和对照样地的土壤温度和含水量变化见图1,增温对土壤温度和水分的影
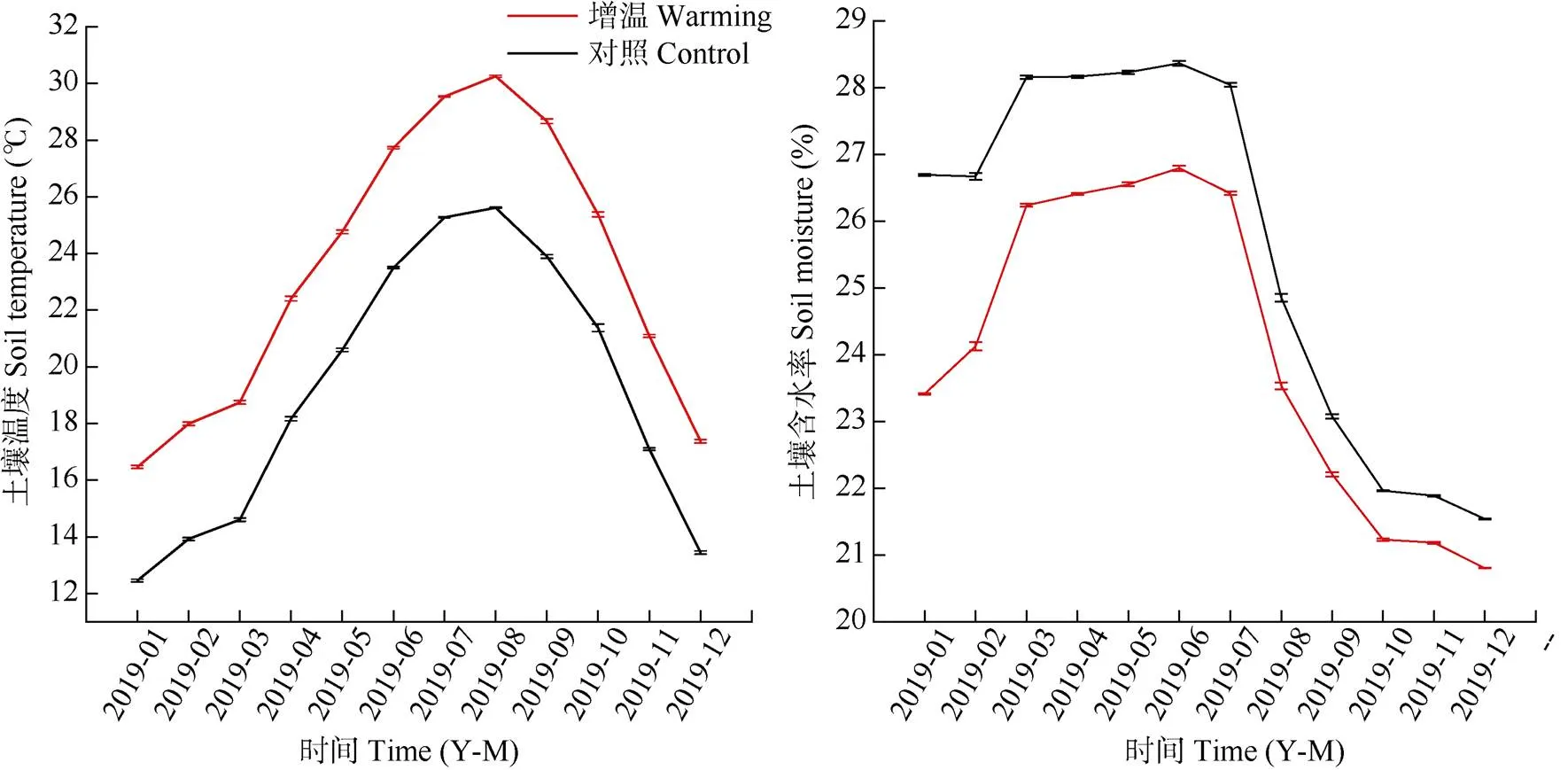
图1 0~10 cm土壤温度和含水率的年变化
1.3 调查方法
于2019年10月1—8日进行林下植被物种多样性调查,每个样地内划分9个2 m×2m小样方, 共90个。样地按二维模型Y、X轴进行,所有植物物种按照生长型划分为草本、灌木、乔木3种类型, 在每个小样方内调查记录所有乔木(幼树)、灌木、草本的种类、个体数、高度和盖度。木质藤本因物种少且高度不高,并入灌木进行分析,草质藤本也因物种数少并入草本分析。计算群落内各植物类型物种的多度、频度、优势度、重要值等[24]。物种学名及其分类学信息查阅《中国植物志》。
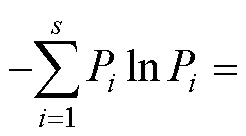
1.4 数据的统计和分析
各样地物种的科属种统计、重要值分析、不同生长型植物重要值的数据和物种多样性指数在Microsoft Excel 2016完成; 样地物种多样性指数在不同增温处理下的变化情况图由Origin 9.1软件绘制; 物种多样性在不同处理下的显著性检验用SPSS 19.0软件进行独立样本检验分析得到。
2 结果和分析
2.1 增温对林下植被物种组成的影响
增温和对照样地林下植被共有植物38科59属77种,其中增温样地37科53属65种,对照样地36科52属63种。为了探明增温和对照样地的优势科属组成情况,将属数≥3的科定为优势科,种数≥3的属定为优势属。增温样地的优势科为茜草科(Rubiaceae)、樟科(Lauraceae)、紫金牛科(Myrsi- naceae), 分别占增温样地总属数的10.8%、10.8%和8.1%。山矾属()为增温样地的优势属,占增温样地总种数的6.3%,其次为冬青属(),占4.7%。樟科为对照样地的优势科,所占总属数比例最高,达到7.9%,其次为茜草科、山茶科(Theaceae)、紫金牛科,分别占6.4%、4.8%和4.8%。优势属为冬青属、杜英属()、润楠属(), 各占总种数的4.8%。增温和对照样地共有科为茜草科、樟科、紫金牛科,共有属为冬青属。
2.2 增温对林下植被不同生长型植物重要值的影响
由表1可见,增温对草本重要值大体呈下降趋势,对灌木和乔木大体呈上升趋势。增温后草本中的蕨类植物重要值降幅明显,狗脊蕨()已从对照样地中的第4位(重要值指数为4.369, 下同)降至增温样地中的第11位(2.458),下降幅度为43.74%;团叶鳞始蕨(,3.248)从对照样地的第7位降至第9位(2.917),下降幅度为10.19%;草本植物华山姜()在增温和对照样地中仍处于优势地位,增温反而使其重要值上升5.48%。灌木的重要值大体呈现上升趋势。山血丹()重要值涨幅明显, 已从对照样地的第3位(4.610)升至增温样地中的第2位(7.140),重要值上升54.88%;网脉酸藤子()增温后重要值提高1.41%。乔木的重要值大体也呈上升趋势。增温后华南木姜子()重要值虽下降了22.22%,但在样地中仍位居第3位;山矾()重要值上升了3.82%;光叶山矾()从对照样地的第11位(2.159)升至增温样地的第6位(3.589),重要值上升66.23%。
总之,乔木和灌木的优势地位在上升,草本优势地位大体在下降,这表明增温并不利于草本植物的生存,尤其是草本中的蕨类植物。
2.3 增温对不同生活型植物的影响
与对照相比,增温使草本、灌木和乔木数量呈现上升趋势,但均未达到显著性差异(>0.05);增温对草本、灌木和乔木的数量、盖度和高度没有显著性影响(>0.05)。增温乔木盖度增加22.61%,但草本和灌木的盖度分别降低4.97%和21.75%。草本、灌木和乔木的高度分别降低21.64%、3.37%和5.59% (图2)。
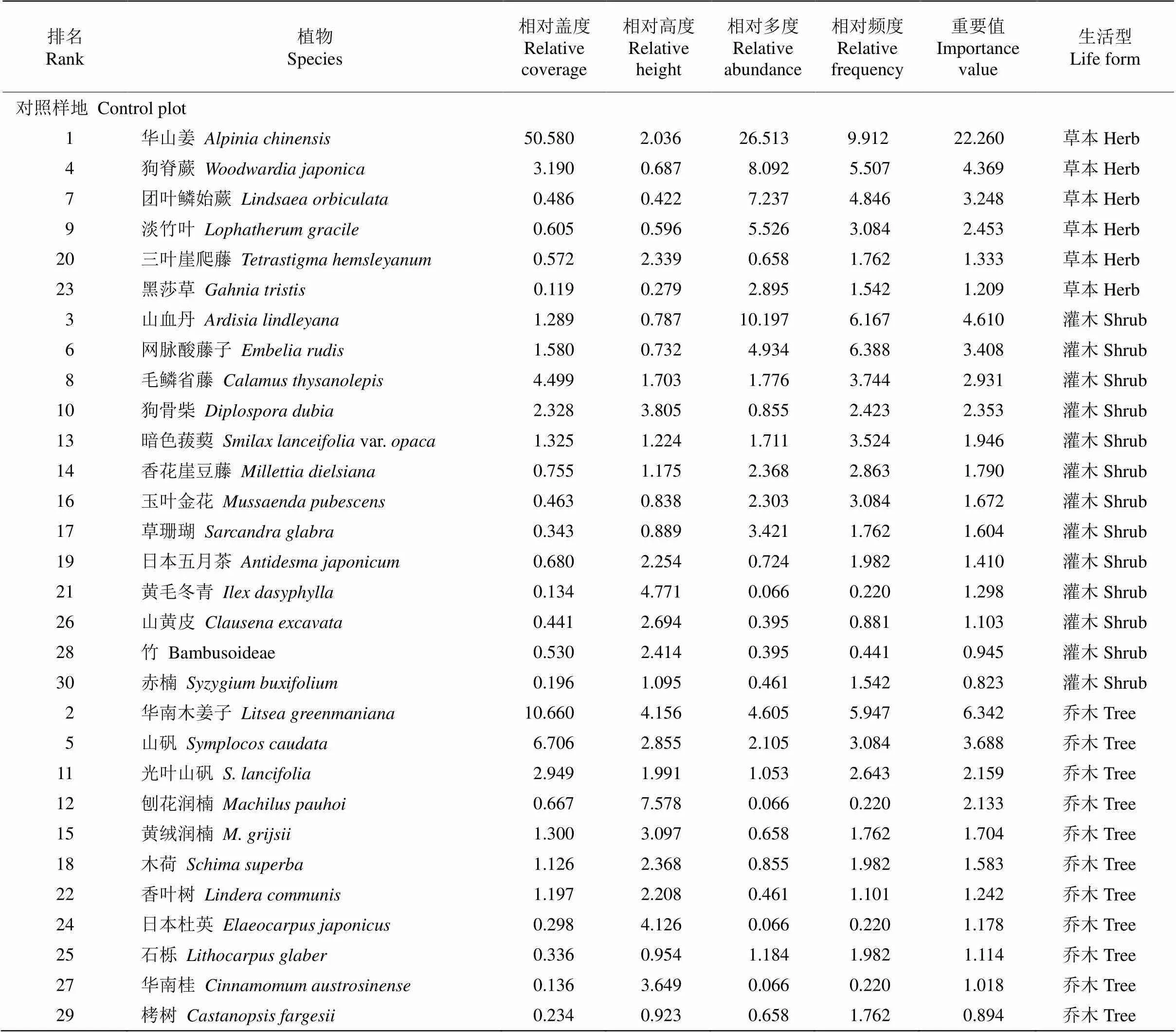
表1 增温和对照样地植物的重要值
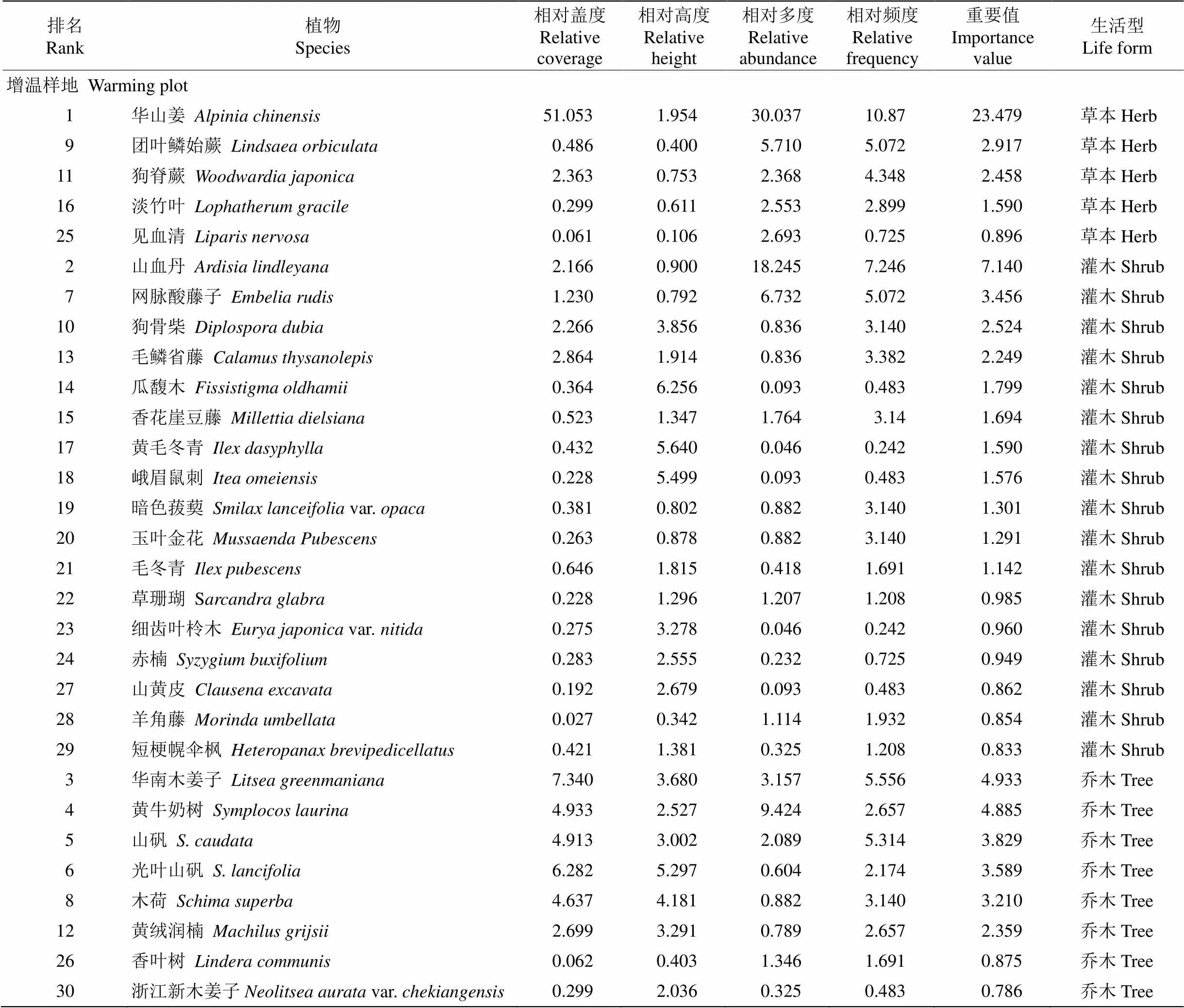
续表(Continued)
2.4 林下植被物种多样性对增温的响应
增温样地植物的Shannon指数、Simpson指数和Pielou指数随增温均呈递减趋势(图3),相比灌木和乔木,草本植物各指数在增温处理下降幅度最大。相比对照,增温导致乔木的Shannon-Wiener指数、Simpson指数和均匀度指数分别降低11.48%、9.35%和12.03%;而灌木和草本分别降低18.88%、16.43%、15.68%和24.79%、21.04%、10.06%, 但均未达显著差异(>0.05),表明增温虽有降低林下植被物种多样性指数的趋势,对物种多样性产生了一定的负面影响,但对林下植被物种多样性无显著差异。
3 结论和讨论
3.1 增温对林下植被物种及优势科属的影响
在全球变暖背景下,植物群落中一些物种因对温度升高敏感,因此可能改变种间竞争,使优势物种和群落组成发生改变[29]。本研究结果表明,增温后华山姜的重要值依然排第1位,重要值上升5.48%;山血丹从对照样地的第3位,升至增温样地中的第2位,重要值上升54.88%;华南木姜子在增温样地中位于第3位;山矾的重要值上升了3.82%,这些物种的重要值依然保持在前列,证明增温并没有改变这些物种的优势地位,这可能与优势物种在群落中的稳定性有关[30]。Ma等[31]在青藏高原高寒草地增温试验表明,以异真草()、藏南犁头尖()、毛细嵩草()等优势物种的稳定性是影响群落稳定性的重要因素。Wu等[32]在内蒙古锡兹旺旗的沙漠大草原进行了为期10年的野外增温试验,认为增温(1.3 ℃)引起的物种丰富度下降并不是群落稳定性下降的重要驱动力,而是优势物种[短花针茅()、银灰旋花()、木地肤()和无芒隐子草()]稳定性下降引起的。
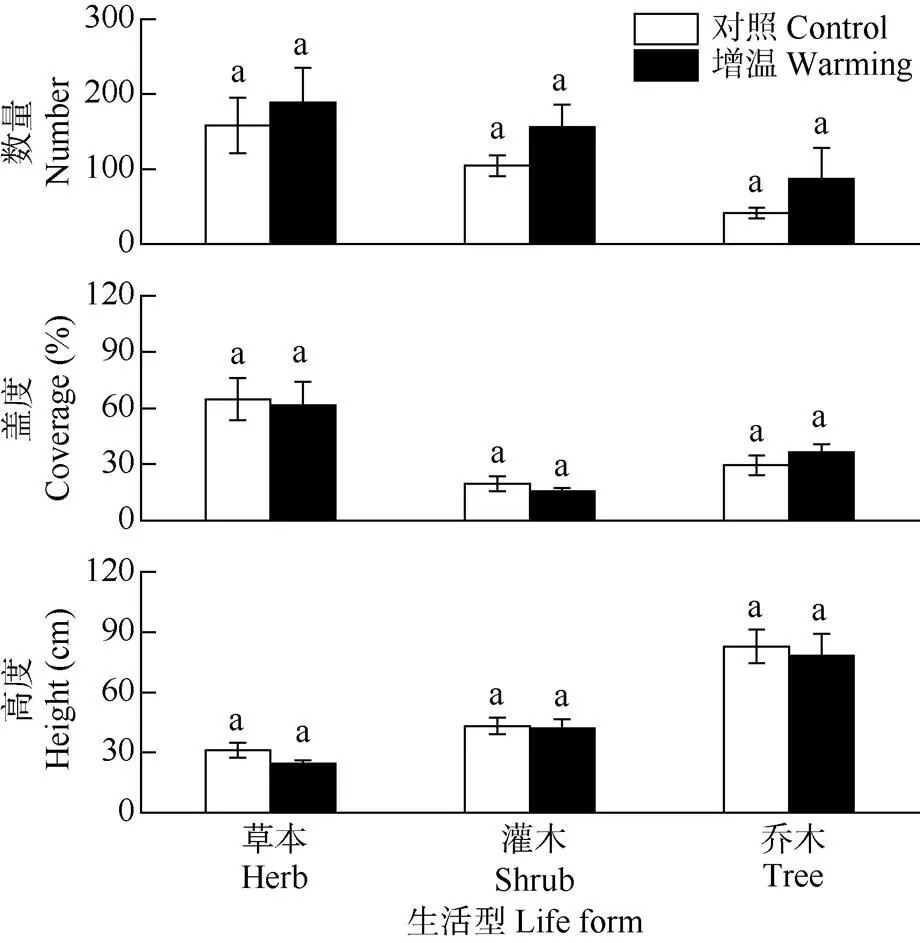
图2 增温对不同生活型植物数量、盖度、高度的影响
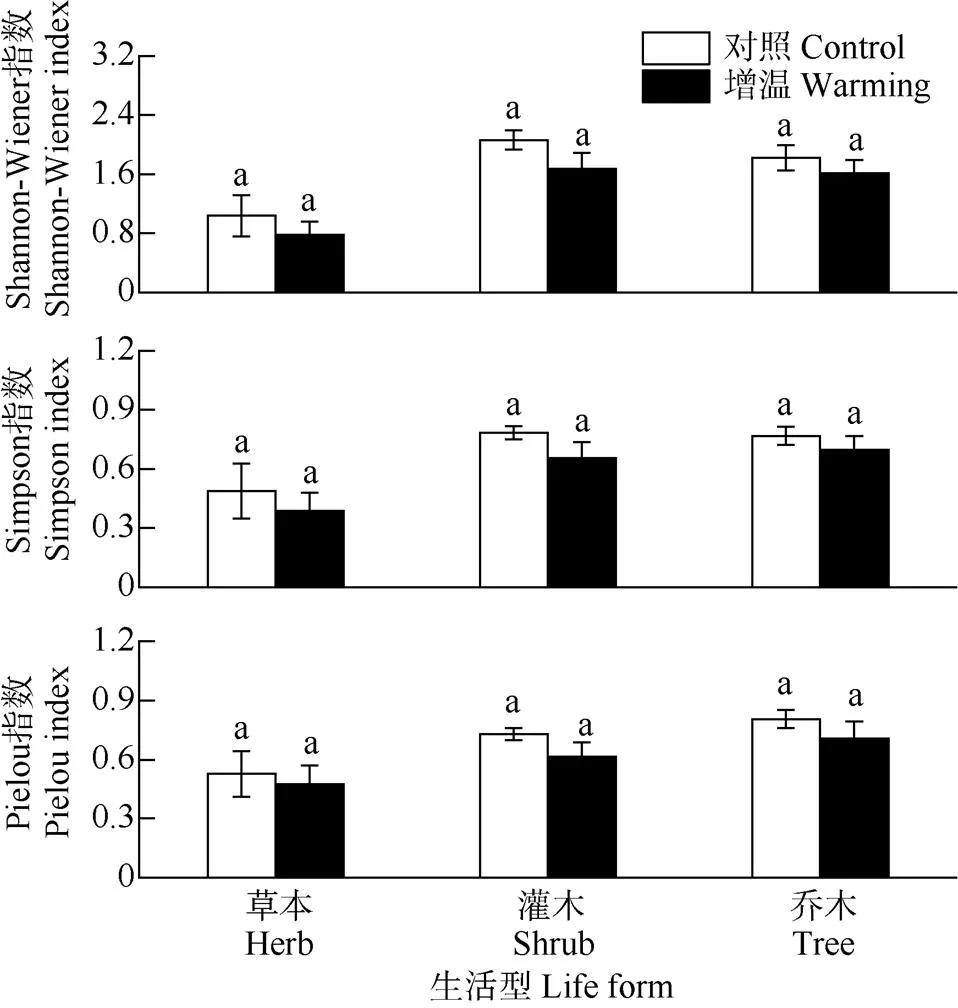
图3 增温对不同生活型植物多样性指数的影响
与对照样地相比,山茶科不再是增温样地中的优势科,增温与对照样地中共同优势科中的茜草科、樟科和紫金牛科,所占比例分别上升了4.46%、2.87%和3.35%。这类植物的地理分布范围广,且主要位于热带亚热带地区,加之本地区年均温为19.6 ℃,冬天无严寒,植物对于全年的高温变化产生一定的适应性,相对于其他中高纬度地区,本地区植物对于高温的适应能力更强[33–35];并且这些植物以乔灌木为主,地下部分根系较草本植物更深, 有利于吸收更深层的土壤水分[36],从而缓解与适应增温带来的土壤水分降低造成的生理胁迫;另一方面,茜草科、樟科和紫金牛科的革质叶面结构,可提高植物的能量反射、降低蒸腾速度,更有利于在缺水环境下生存[37–38]。冬青属作为增温和对照样地中仅存的共同优势属,因其在温带至热带大范围内分布,对温度变化适应性强,同时同样具有革质叶面结构,可在较干旱、贫瘠土壤中生存[39],这些特征均可能是导致此类植物成为林下植被物种优势属的重要原因。
3.2 植被群落特征对增温的响应
本研究中增温使草本、灌木和乔木的数量呈现上升趋势,乔木盖度增加,草本和灌木的盖度分别降低,同时3种生长型植物高度也分别降低,但各植被指标的方差分析均未达到显著水平,表明短期增温对植被影响不显著。在高寒草甸,李娜等[40]通过2年增温试验,认为植物群落高度整体呈增加趋势,群落上层的建群种和主要伴生种的盖度有所增加。增温对松嫩草原的羊草()种群高度的影响不显著,而对密度具有一定的促进作用[41]。吴红宝等[42]的研究表明,放牧和模拟增温对高寒草地植物高度具有显著交互作用,不放牧条件下增温显著增加了植物高度。赵艳艳等[43]研究也表明,增温后灌丛群落的高度和盖度均显著高于对照,说明增温对不同群落的影响不同,16年的长期增温更有利于灌丛群落的生长。但这些研究结果与本文具有一定差异,这表明增温会因时间尺度、区域差异及植被类型的不同来影响植被群落特征,进而在一定程度上影响植物生长以及群落发育,即增温对植物产生的影响既有时间尺度依赖性,同时增温改变土壤湿度[44],加剧土壤水分蒸发,加之土壤养分含量的差异变化[45],最终改变植物对土壤水分和养分的吸收[46],进而影响植物的群落组成。
3.3 增温对不同生活型植物重要值的影响
增温可通过改变土壤水分,对群落的结构和组成产生影响[3]。华山姜是本研究中林下植被重要值最大的草本植物,在群落中占主导地位,增温使其重要值上升了5.48%,但下层草本中的蕨类植物, 如狗脊蕨、团叶鳞始蕨的重要值分别下降43.74%和10.19%,这表明增温不利于草本植物中蕨类生长。灌木中的山血丹、网脉酸藤子的重要值分别上升54.88%和1.41%,上层植物中的光叶山矾重要值上升66.23%,木荷上升幅度为102.78%。这些主要的灌木及乔木优势树种的重要值开始上升,在林下逐渐成为优势树种,草本植物有逐渐被灌木及乔木所取代的趋势,这与赵艳艳等[43]的16年长期观察结果一致,这可能是由于增温后导致土壤中含水量降低,林下蕨类植物根系较浅,并相对处于缺水状态,同时与林下萌生的乔木幼树产生强烈的竞争而生长不良或干枯死亡导致。
此外,不同生长型植物对增温的不同响应,可能也与植物的生长特性和生活习性有关。华山姜作为林荫下常见的多年生草本,其根茎处萌芽数量多,根系发达,能吸收更远及更深层土壤中的水分,更有利于在相对缺水的土壤环境下存活[47]。而华南木姜子在增温后重要值下降,可能与增温缩小其生长分布有关。例如郑维艳等[48]利用最大墒模型(Maxent)预测中国4种木姜子属()资源植物在我国当代、未来(2061—2080年)气候变暖条件下的潜在分布区,结果显示华南木姜子在两种二氧化碳浓度情景(RCP 2.6、RCP 8.5)中适宜区面积分别减少1.0%和5.6%,其生长范围的收缩可能随未来气候的变化而发生改变,华南地区的温度上升和降水的减少,以及极端高温事件发生频率明显升高而导致该种的适宜分布区逐渐缩小。作为像狗脊蕨一类的草本植物,因其高度低于生长在其上部的华山姜以及像华南木姜子一类的高大乔木,而在林下形成一种郁闭环境,使草本植物地上部分生长空间受限(盖度下降,高度降低)。另外,从本地全年土壤含水率的数据来看,增温使10 cm的土壤含水率极显著下降(<0.01),植物根系可利用的水分逐渐降低,物种间竞争加强[49],植物为了获取充足的水分和营养物质,使根系向土壤深处生长分布(根系的向水性)[50]。本地区土壤增温对深根植物杉木细根形态特征的研究也表明,增温不仅使各土层单位根窗面积细根根长大于对照,在10~20和20~30 cm土层中,单位根窗面积细根根长及细根数量均大于表层[51],这表明增温可引起乔木地下根系向土壤深层转移,草本植物因自身根系较浅,受土壤水分限制严重[52], 土壤增温使其很难获得浅层土壤中更多的水分,最终不利于其生长发育,这可能是导致草本植物重要值下降的重要原因。
3.4 增温对不同生长型植物物种多样性的影响
全球气候变化引起的温度和降水格局的改变是影响生物多样性的重要因素[53]。本研究表明,草本、灌木和乔木的Simpson指数、Shannon指数和Pielou指数在增温状态下呈现下降趋势,表明增温对物种多样性产生了一定的负面影响(>0.05)。
有研究认为,土壤水分减少、养分含量的改变及优势物种稳定性是导致物种多样性对增温不敏感的原因。在高寒草甸为期6年的模拟增温试验中,物种丰富度在4种不同梯度增温处理下(+0.6 ℃、+0.98 ℃、+1.25 ℃、+1.88 ℃)没有显著变化,说明物种丰富度对短期增温的响应并不敏感[54],而长达16年的增温表明,草甸群落土壤的各层全氮、全钾、有机质含量均降低,而各层速效钾、速效磷含量均增加,说明增温改变了其养分总体组成含量,加速了养分的分解利用,导致植物物种多样性略微增加[43]。Yang等[29]的研究表明,半灌木冷蒿()对高达55%的群落覆盖有贡献,群落稳定性与其稳定性呈正相关,表明优势物种的稳定性是影响群落稳定性的重要因素。
此次天然林林下植被多样性的调查是在增温后的第4年,因增温导致土壤水分蒸发,使局部小气候变得干燥,物种多样性呈现下降趋势,但这种趋势并不明显,可能原因是优势物种是保持群落稳定性的重要原因[32],随着增温进行,这类温度适应性较强的植物物种可能表现出吸热的生存策略[55]。Yang等[56]报道,为期8年的增温,物种多样性指数波动较小,Shi等[29]进行了连续14年的增温试验, 由于入侵物种和优势物种的负竞争,物种多样性在第8年开始下降,预计长期变暖可能改变草原群落结构。这些研究结果再次证实物种的消失或是新物种的出现均是一个长时间序列下的群落演替过程[57],这些均可能是导致本地区物种多样性对增温的响应不敏感的原因。但如果增温持续性发展,很大程度上会使物种趋于单一化发展,导致群落发生演替,最终引起物种多样性降低。
综上,短期增温对林下植被物种数的改变并不明显,各生活型植物重要值在增温后呈现下降趋势(>0.05);Shannon指数、Simpson指数和Pielou指数随增温均呈递减趋势,表明增温对物种多样性产生了一定负面影响。随温度升高,不同生活型植物由草本向灌木和乔木转化。这可能是增温处理下, 浅层土壤温度增加抑制了草本植物生长,从而导致整个群落向灌木和乔木方向进行演替。
[1] KAY J E. Early climate models successfully predicted global warming [J]. Nature, 2020, 578(7793): 45–46. doi: 10.1038/d41586-020-0024 3-w.
[2] IPCC. Climate change 2021: The physical science basis [J]. Chem Int, 2021, 43(4): 22–23. doi: 10.1515/ci-2021-0407.
[3] ZHU J T, ZHANG Y J, YANG Xet al. Warming alters plant phylo- genetic and functional community structure [J]. J Ecol, 2020, 108(6): 2406–2415. doi: 10.1111/1365-2745.13448.
[4] WU T, QU C, LI Y Y, et al. Warming effects on leaf nutrients and plant growth in tropical forests [J]. Plant Ecol, 2019, 220(7/8): 663–674. doi: 10.1007/s11258-019-00943-y.
[5] CHEN F, YU S L, SHANG H M, et al. The productivity of low- elevation juniper forests in central Asia increased under moderate warming scenarios [J]. J Geophys Res Biogeosci, 2021, 126(4): e2021JG006269. doi: 10.1029/2021JG006269.
[6] HE Y Z, HUANG W D, ZHAO X, et al. Review on the impact of climate change on plant diversity [J]. J Desert Res, 2021, 41(1): 59–66. [何远政, 黄文达, 赵昕, 等. 气候变化对植物多样性的影响研究综述[J]. 中国沙漠, 2021, 41(1): 59–66. doi: 10.7522/j.issn.1000-694X. 2020.00104.]
[7] SEDDON A W R, MACIAS-FAURIA M, LONG P R, et al. Sensitivity of global terrestrial ecosystems to climate variability [J]. Nature2016, 531(7593): 229–232. doi: 10.1038/nature16986.
[8] KENNEDY D, NORMAN C. What don’t we know? [J]. Science2005, 309(5731): 75. doi: 10.1126/science.309.5731.75.
[9] LI T, XIONG Q L, LUO P, et al. Direct and indirect effects of environmental factors, spatial constraints, and functional traits on shaping the plant diversity of montane forests [J]. Ecol Evol, 2020, 10(1): 557–568. doi: 10.1002/ece3.5931.
[10] KULONEN A. Faster, Taller, More-Patterns and Drivers of Floristic Change on European Mountain Summits [M]. Bergen: University of Bergen, 2017.
[11] LI T, LUO P, XIONG Q L, et al. Spatial heterogeneity of tree diversity response to climate warming in montane forests[J]. Ecol Evol, 2021, 11(2): 931–941. doi: 10.1002/ece3.7106.
[12] LENOIR J, GÉGOUT J C, MARQUET P A, et al. A significant upward shift in plant species optimum elevation during the 20th century [J]. Science, 2008, 320(5884): 1768–1771. doi: 10.1126/science.1156831.
[13] XU Y X, WANG Z C, ZHU W K, et al. Litterfall and nutrient cycling of Eucalyptus plantation with different ages on Leizhou Peninsula [J]. J Trop Subtrop Bot, 2019, 27(4): 359–366. [许宇星, 王志超, 竹万宽, 等. 雷州半岛桉树人工林凋落物量和养分循环研究 [J]. 热带亚热带植物学报, 2019, 27(4): 359–366. doi: 10.11926/jtsb.3986.]
[14] LIAO Y J, HONG W, CHEN F Q, et al. Studies on species composition and diversity of-mixed forest in Guangzhou [J]. J Trop Subtrop Bot, 2021, 29(5): 494–502. [廖宇杰, 洪维, 陈富强, 等. 广州红锥-马占相思林物种组成与多样性研究[J]. 热带亚热带植物学报, 2021, 29(5): 494–502. doi: 10.11926/jtsb. 4361.]
[15] LI X J, LIU X F, LIN C F, et al. Effects of experimental soil warming on plant biomass allocation during the early stages of succession in a subtropical forest in China [J]. Acta Ecol Sin, 2017, 37(1): 25–34. [李晓杰, 刘小飞, 林成芳, 等. 土壤增温调节中亚热带森林更新初期植物生物量分配格局 [J]. 生态学报, 2017, 37(1): 25–34. doi: 10. 5846/stxb201607261529.]
[16] DUSENGE M E, WAY D A. Warming puts the squeeze on photo- synthesis: Lessons from tropical trees [J]. J Exp Bot, 2017, 68(9): 2073–2077. doi: 10.1093/jxb/erx114.
[17] KRAUSE G H, CHEESMAN A W, WINTER K, et al. Thermal tolerance, net CO2exchange and growth of a tropical tree species,, cultivated at elevated daytime and nighttime tempe- ratures [J]. J Plant Physiol, 2013, 170(9): 822–827. doi: 10.1016/j. jplph.2013.01.005.
[18] Dong S Y, Gao X J. Long-term climate change: Interpretation of IPCC fifth assessment report [J]. Prog Inquisit Mutat Clim, 2014, 10(1): 56–59. [董思言, 高学杰. 长期气候变化——IPCC第五次评估报告解读 [J]. 气候变化研究进展, 2014, 10(1): 56–59. doi: 10.3969/j.issn. 1673-1719.2014.01.012.]
[19] SOMMER J H, KREFT H, KIER G, et al. Projected impacts of climate change on regional capacities for global plant species richness [J]. Proc R Soc B, 2010, 277(1692): 2271–2280. doi: 10.1098/rspb.2010.0120
[20] COTTO O, WESSELY J, GEORGES D, et alA dynamic eco-evolu- tionary model predicts slow response of alpine plants to climate warming [J]. Nat Commun, 2017, 8: 15399. doi: 10.1038/ncomms 15399.
[21] SU J Q, HAN X, CHEN B M. Do day and night warming exert different effects on growth and competitive interaction between invasive and native plants? [J]. Biol Invasions2021, 23(1): 157–166. doi: 10.1007/s10530-020-02362-x.
[22] LI C. Effect of warming in different depth of soil carbon emissions on the natural forest of[D]. Fuzhou: Fujian Normal University, 2018. [李超. 增温对格氏栲天然林不同深度土壤碳排放的影响[D]. 福州: 福建师范大学, 2018.]
[23] YUAN S. Effect of warming on quantity and composition of soil dissolved organic matter in subtropical forest [D]. Fuzhou: Fujian Normal University, 2018. [袁硕. 增温对亚热带常绿阔叶林土壤可溶性有机物数量与组成的影响[D]. 福州: 福建师范大学, 2018.]
[24] JIANG Z K, JI Y, LIU Y H, et al. Composition and diversity of understory vegetation of 50 years old Chinese fir plantation [J]. J Subtrop Resour Environ, 2020, 15(3): 32–38. [蒋宗垲, 籍烨, 刘雨晖, 等. 老龄杉木人工林林下植被组成与多样性特征[J]. 亚热带资源与环境学报, 2020, 15(3): 32–38. doi: 10.19687/j.cnki.1673-7105. 2020.03.005.]
[25] GE F. Modern Ecology [M]. Beijing: Science Press, 2008: 1–643. [戈峰. 现代生态学 [M]. 北京: 科学出版社, 2008: 1–643.]
[26] CHAO Q, WEN J, YANG X Y, et al. Responses of subalpine meadow species diversity to simulated warming in the Yunding Mountain [J]. Environ Ecol, 2019, 1(4): 34–40. [晁倩, 温静, 杨晓艳, 等. 云顶山亚高山草甸植物物种多样性对模拟增温的响应 [J]. 环境生态学, 2019, 1(4): 34–40.]
[27] WHITTAKER R H. Evolution and measurement of species diversity [J]. Taxon, 1972, 21(2/3): 213–251. doi: 10.2307/1218190.
[28] PIELOU E C. Ecological Diversity [M]. New York: Wiley, 1975: 1–318.
[29] SHI Z, SHERRY R, XU X, et al. Evidence for long-term shift in plant community composition under decadal experimental warming [J]. J Ecol, 2015, 103(5): 1131–1140. doi: 10.1111/1365-2745.12449.
[30] YANG Z L, ZHANG Q, SU F L, et al. Daytime warming lowers community temporal stability by reducing the abundance of dominant, stable species [J]. Glob Change Biol, 2017, 23(1): 154–163. doi: 10. 1111/gcb.13391.
[31] MA Z Y, LIU H Y, MI Z R, et al. Climate warming reduces the temporal stability of plant community biomass production [J]. Nat Commun, 2017, 8: 15378. doi: 10.1038/ncomms15378.
[32] WU Q, REN H Y, WANG Z W, et al. Additive negative effects of decadal warming and nitrogen addition on grassland community stability [J]. J Ecol, 2020, 108(4): 1442–1452. doi: 10.1111/1365-2745. 13363.
[33] LO H S. Flora Reipublicae Popularis Sinicae, Tomus 71(1) [M]. Beijing: Science Press, 1999: 1–442. [罗献瑞. 中国植物志, 第71卷第1分册[M]. 北京: 科学出版社, 1999: 1–432.]
[34] LI X W. Flora Reipublicae Popularis Sinica, Tomus 31 [M]. Beijing: Science Press, 1982: 1–534. [李锡文. 中国植物志, 第31卷 [M]. 北京: 科学出版社, 1982: 1–534.]
[35] CHEN J. Flora Reipublicae Popularis Sinica, Tomus 58 [M]. Beijing: Science Press, 1979: 1–158. [陈介. 中国植物志, 第58卷 [M]. 北京: 科学出版社, 1979: 1–158.]
[36] YU X C, YAO B Q, ZHOU H K, et al. Variable responses to long-term simulated warming of underground biomass and carbon allocations of two alpine meadows on the Qinghai-Tibet Plateau [J]. Chin Sci Bull, 2015, 60(4): 379–388. [余欣超, 姚步青, 周华坤, 等. 青藏高原两种高寒草甸地下生物量及其碳分配对长期增温的响应差异 [J]. 科学通报, 2015, 60(4): 379–388. doi: 10.1360/n972014-00473.]
[37] CAO X, SHEN Q, LIU L, et al. Relationships of growth, stable carbon isotope composition and anatomical properties of leaf and xylem in seven mulberry cultivars: A hint towards drought tolerance [J]. Plant Biol, 2020, 22(2): 287–297. doi: 10.1111/plb.13067.
[38] FLETCHER L R, CUI H X, CALLAHAN H, et al. Evolution of leaf structure and drought tolerance in species of[J]. Am J Bot, 2018, 105(10): 1672–1687. doi: 10.1002/ajb2.1164.
[39] YAO X, SONG Y, YANG J B, et al. Phylogeny and biogeography of the hollies (L., Aquifoliaceae) [J]. J Syst Evol, 2021, 59(1): 73–82. doi: 10.1111/jse.12567.
[40] LI N, WANG G X, YANG Y, et al. Short-term effects of temperature enhancement on community structure and biomass of alpine meadow in the Qinghai-Tibet Plateau [J]. Acta Ecol Sin, 2011, 31(4): 895–905. [李娜, 王根绪, 杨燕, 等. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响[J]. 生态学报, 2011, 31(4): 895–905.]
[41] QI H, LUO W, SUN Y J, et al. Responses ofquantity characteristic to simulated warming and nitrogen application in Songnen Grassland [J]. J NE Norm Univ (Nat Sci), 2013, 45(2): 112– 117. [齐红, 罗微, 孙亚娟, 等. 松嫩草原羊草种群数量特征对增温和施氮的响应[J]. 东北师大学报(自然科学版), 2013, 45(2): 112– 117.]
[42] WU H B, GAO Q Z, GANJURJAV H, et al. Effects of grazing and simulated warming on plant community structure and productivity of alpine grassland in northern Xizang, China [J]. Chin J Plant Ecol, 2019, 43(10): 853–862. [吴红宝, 高清竹, 干珠扎布, 等. 放牧和模拟增温对藏北高寒草地植物群落特征及生产力的影响[J]. 植物生态学报, 2019, 43(10): 853–862. doi: 10.17521/cjpe.2018.0288.]
[43] ZHAO Y Y, ZHOU H K, YAO B Q, et al. The influence of long-term simulating warming to the plant community and soil nutrient of alpine meadow [J]. Acta Agrest Sin, 2015, 23(4): 665–671. [赵艳艳, 周华坤, 姚步青, 等. 长期增温对高寒草甸植物群落和土壤养分的影响[J]. 草地学报, 2015, 23(4): 665–671. doi: 10.11733/j.issn.1007-0435.2015. 04.001.]
[44] LIU H Y, MI Z R, LIN L, et al. Shifting plant species composition in response to climate change stabilizes grassland primary production [J]. Proc Natl Acad Sci USA, 2018, 115(16): 4051–4056. doi: 10.1073/pnas. 1700299114.
[45] YU C Q, HAN F S, FU G. Effects of 7 years experimental warming on soil bacterial and fungal community structure in the northern Tibet alpine meadow at three elevations [J]. Sci Total Environ, 2019, 655: 814–822. doi: 10.1016/j.scitotenv.2018.11.309.
[46] KLEIN J A, HARTE J, ZHAO X Q. Dynamic and complex microclimate responses to warming and grazing manipulations [J]. Glob Change Biol, 2005, 11(9): 1440–1451. doi: 10.1111/j.1365-2486. 2005.00994.x.
[47] GAO X M. Cloning and RNA in situ hybridization of calmodulin gene in[J]. Acta Bot Boreali-Occid Sin, 2005, 25(9): 1730–1734. [高雪梅. 华山姜中钙调素基因的克隆及其RNA原位杂交[J]. 西北植物学报, 2005, 25(9): 1730–1734. doi: 10.3321/j.issn: 1000-4025.2005.09.004.]
[48] ZHENG W Y, CAO K F. Impact of future climate change on potential geographical distribution of fourspecies in China [J]. Guihaia, 2020, 40(11): 1584–1594. [郑维艳, 曹坤芳. 未来气候变化对四种木姜子地理分布的影响[J]. 广西植物, 2020, 40(11): 1584–1594. doi: 10.11931/guihaia.gxzw201904020.]
[49] XU M H, LIU M, XUE X, et al. Warming effects on plant biomass allocation and correlations with the soil environment in an alpine meadow, China [J]. J Arid Land, 2016, 8(5): 773–786. doi: 10.1007/s 40333-016-0013-z.
[50] HOOVER D L, DUNIWAY M C, BELNAP J. Testing the apparent resistance of three dominant plants to chronic drought on the Colorado Plateau [J]. J Ecol, 2017, 105(1): 152–162. doi: 10.1111/1365-2745. 12647.
[51] XIONG D C, LIU X F, CHEN S D, et al. Effects of soil warming on fine root morphology of Chinese fir seedlings [J]. J Subtrop Res Environ, 2014, 9(3): 89–91. [熊德成, 刘小飞, 陈仕东, 等. 土壤增温对杉木幼苗细根形态特征的影响[J]. 亚热带资源与环境学报, 2014, 9(3): 89–91. doi: 10.3969/j.issn.1673-7105.2014.03.012.]
[52] ZHU J T. Effects of experimental warming on plant reproductive phenology in Xizang alpine meadow [J]. Chin J Plant Ecol, 2016, 40(10): 1028–1036. [朱军涛. 实验增温对藏北高寒草甸植物繁殖物候的影响[J]. 植物生态学报, 2016, 40(10): 1028–1036. doi: 10. 17521/cjpe.2016.0068.]
[53] LI P, SAYER E J, JIA Z, et al. Deepened winter snow cover enhances net ecosystem exchange and stabilizes plant community composition and productivity in a temperate grassland [J]. Glob Change Biol, 2020, 26(5): 3015–3027. doi: 10.1111/gcb.15051.
[54] MA L, ZHANG Q, ZHANG Z H, et al. Effects of gradient warming on species diversity and biomass in alpine meadows [J]. Acta Agrest Sin, 2020, 28(5): 1395–1402. [马丽, 张骞, 张中华, 等. 梯度增温对高寒草甸物种多样性和生物量的影响[J]. 草地学报, 2020, 28(5): 1395– 1402.]
[55] ZHANG C H, WILLIS C G, KLEIN J A, et al. Recovery of plant species diversity during long-term experimental warming of a species- rich alpine meadow community on the Qinghai-Tibet Plateau [J]. Biol Conserv, 2017, 213: 218–224. doi: 10.1016/j.biocon.2017.07.019.
[56] YANG Y W, LI X L, ZHOU X H, et al. Study on relationship between plant community degradation and soil environment in an alpine meadow [J]. Acta Agrest Sin, 2016, 24(6): 1211–1217. [杨元武, 李希来, 周旭辉, 等. 高寒草甸植物群落退化与土壤环境特征的关系研究 [J]. 草地学报, 2016, 24(6): 1211–1217. doi: 10.11733/j.issn.1007- 0435.2016.06.009.]
[57] KLEIN J A, HARTE J, ZHAO X Q. Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau [J]. Ecol Lett, 2004, 7(12): 1170–1179. doi: 10.1111/j. 1461-0248.2004.00677.x.
Effects of Short-term Warming on Species Diversity of Understory Vegetation in Subtropical Evergreen Broad-leaved Forest
JI Ye1,2, CHEN Shidong1,2, XIONG Decheng1,2, XU Chao1,2, LIU Xiaofei1,2, HE Zongming3, YANG Zhijie1,2*
(1. School of Geographical Sciences, Fujian Normal University,Fuzhou 350007, China; 2.Fujian Sanming Forest Ecosystem National Observation and Research Station,Sanming 365000, Fujian, China; 3. College of Forestry, Fujian Agriculture and Forestry University, Fuzhou 350002, China)
In order to understand the impact of climate warming on species diversity of understory vegetation, the effects of short-term 4-year warming (4 ℃) on understory vegetation diversity in subtropical evergreen broad-leaved natural forest were studied by means of soil warming. The results showed that short-term warming had no significant effect on the species composition of understory vegetation (>0.05). There were 77 common species in understory vegetation, belonging to 38 families and 59 genera, including 65 species 53 genera and 37 families in warming plot and 63 species 52 genera and 36 families in control plot. Short-term warming increased the coverage of trees by 22.61%, and decreased the coverage of herbs and shrubs by 4.97% and 21.75%, respectively, and warming reduced the height of herbs, shrubs and trees by 21.64%, 3.37% and 5.59%, respectively. The ranking of importance value of ferns in herbaceous plants decreased after warming, and the importance value of trees increased (>0.05). Although there was no significant difference in species diversity indexes after warming (>0.05), they all decreased with warming. Therefore, the species composition of understory vegetation was not sensitive to short-term warming, which decreased the importance value of ferns in herbaceous plants and negatively affected the species diversity indexes. However, this response was not sensitive, and it was predicted that long-term warming might lead to the succession of the whole community from herbaceous to shrub and tree.
Understory vegetation; Species diversity; Soil warming; Subtropical forest
10.11926/jtsb.4575
2021-11-22
2022-03-11
国家自然科学基金项目(31930071);国家重点基础研究发展计划课题(2014CB954003)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 31930071), and the National Key Basic Research and Development Planning Program (Grant No. 2014CB954003).
籍烨(1987年生),女,在读博士研究生, 主要从事生态环境与全球变化研究。E-mail: jiye0518@126.com
通讯作者Corresponding author.E-mail: zhijieyang@fjnu.edu.cn
猜你喜欢
中华建设(2023年10期)2023-10-07
建筑与预算(2022年6期)2022-07-08
少儿美术(2020年8期)2020-11-06
乡村科技(2018年8期)2018-02-13
现代园艺(2017年13期)2018-01-19
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
小说月刊(2014年1期)2014-04-23
建筑与预算(2014年5期)2014-04-11
河南科技(2014年16期)2014-02-27
