
抗氧化系统参与水杨酸诱导烟草幼苗抗冷性提高的生化机制
2023-02-27 03:08:56孔春艳徐照丽
热带作物学报 2023年1期
孔春艳,赵 静,徐照丽,龚 明*
1.云南师范大学生命科学学院/生物能源持续开发利用教育部工程研究中心/云南省生物质能与环境生物技术重点实验室,云南昆明 650500;2.云南省农业科学院甘蔗研究所,云南开远 661699;3.云南省烟草农业科学研究院,云南昆明 650021
低温胁迫是限制植物地理分布、影响农作物产量和品质的主要环境因子之一[1]。烟草是喜温植物,对低温较为敏感,25~28℃是烟草生长的最适温度[2]。在烟草农业生产实践中,早春低温常常导致育苗期的烟草幼苗发生冷害,进而影响烟草大田移栽后的早生快发、大田生长及烟叶的产量和品质[3-4]。因此,通过各种农艺措施来提高烟苗的抗冷性对烟草农业生产有重要意义。
水杨酸(salicylic acid, SA)是广泛存在于高等植物体内的一种小分子酚类物质,被大量应用于缓解植物低温胁迫[5-7]。已有研究表明,由各种抗氧化酶和抗氧化剂组成的细胞抗氧化系统在植物对低温胁迫响应与适应中起重要作用[8-9]。陈丹等[10]研究发现,SA预处理对低温胁迫下蝴蝶兰具有缓解效应,能提高蝴蝶兰叶片中的叶绿素、抗坏血酸(ASA)和谷胱甘肽(GSH)含量,从而缓解低温胁迫对蝴蝶兰的伤害。辛慧慧等[11]研究表明,SA预处理可以显著降低棉花幼苗叶片丙二醛(MDA)的积累,提高超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的活性来适应低温环境。前期通过对烟草幼苗喷施不同浓度的 CaCl2、赤霉素 GA3、水杨酸和多效唑(PP333)进行预处理,比较了这4种化学调控剂对10℃低温胁迫下烟草幼苗耐冷性及其光合生理特性的影响,结果表明,SA综合效果最好[3]。SA预处理能够有效缓解低温胁迫对烤烟幼苗的伤害,减少低温胁迫下幼苗叶片膜脂过氧化作用,降低质膜透性,提高SOD活性,增大脯氨酸的含量[3, 12]。
目前,有关SA诱导烟草幼苗抗冷性形成的生化途径和分子机制尚未完全清楚。此外,在烟草实际生产中,烟草育苗都是在大棚或温室里进行,育苗期遭遇寒潮时棚内温度一般都能保持在10℃左右,但有关烟草幼苗低温处理实验通常在 4~5℃[2,12],难以准确模拟生产实践中烟草育苗期遭遇寒潮的实际情况。为此,本研究以生产上广泛应用的‘云烟97’和‘K326’2个主栽品种为材料,用浓度0、0.5、1.0、1.5 mmol/L的SA对三叶一心期的烟草幼苗进行预处理后进行10℃的低温处理 12 d,测定烟草幼苗的多项耐冷性指标,并详细研究和比较2个烟草品种中还原型/氧化型抗坏血酸及谷胱甘肽含量的变化,以及5种重要的抗氧化酶活性变化,旨在进一步阐明 SA诱导烟草幼苗抗冷性形成的生理基础和生化途径,为烟草抗冷育苗的实际生产奠定理论基础。
1 材料与方法
1.1 材料
试验材料为烤烟(Nicotiana tabacumL.)主栽品种‘云烟97’和‘K326’,按(GB/T 25241.1—2010)的方法进行漂浮育苗和管理[3]。
1.2 方法
1.2.1 SA预处理及低温胁迫处理 将‘云烟97’和‘K326’2个品种的包衣种子在温室进行播种,采用漂浮育苗方式,温室温度为 25~28℃,自然光照。待幼苗长到三叶一心时,叶面分别喷施浓度为0(CK)、0.5、1.0、1.5 mmol/L的水杨酸,每隔4 d喷施一次,共喷施 5次。喷施时以全株淋湿,药液欲滴下为宜。最后一次喷完后让其生长2 d,后转入人工气候室[光周期为14 h/10 h,光照强度为 400 μmol/(m2·s),室内湿度为 75%~80%]进行10℃低温处理12 d,分别在处理的0、6、12 d测定电导率和根系活力,并进行根系扫描,后取叶片,液氮速冻后保存在-80℃超低温冰箱,用于后续各种测定[3]。
1.2.2 相对生长速率的计算 在低温处理的 0 d(t1)和12 d(t2)分别取长势一致的幼苗15株,每5株1个重复,洗净晾干水分后杀青烘干,称其干重,用下述公式计算相对生长速率(RGR)。
式中:W1、W2分别表示在时间t1和t2时的干重。
1.2.3 MDA含量的测定 MDA含量的测定采用硫代巴比妥酸(TCA)法,参照赵世杰等[13]的方法。
1.2.4 叶片电解质渗漏率的测定 叶片电解质渗漏率的测定采用电导法,参照李合生[14]的方法。
1.2.5 叶绿素含量的测定 叶绿素含量的测定根据王学奎[15]的方法,用95%乙醇提取,分光光度法测定。
1.2.6 根系活力的测定 根系活力的测定根据王学奎[15]的方法,用TTC法测定。
1.2.7 抗氧化剂含量的测定 还原型抗坏血酸(ASA)、氧化型抗坏血酸(DHA)、还原型谷胱甘肽(GSH)、氧化型谷胱甘肽(GSSG)含量的测定参照本实验室方法[16]。
1.2.8 抗氧化酶含量的测定 愈创木酚过氧化物酶(GPX)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)、谷胱甘肽还原酶(GR)酶液的提取参照李忠光等[17]的方法,GPX测定按照CHANCE等[18]的方法,SOD的测定采用氮兰四唑(NBT)还原法测定[19],CAT测定参照 AEBI[20]的方法,APX测定参照NAKANO等[21]的方法,GR测定参照HALLIWELL等[22]的方法。
1.3 数据处理
每组实验进行3次重复,每个实验重复2次。数据用SPSS 21.0软件进行统计分析和t检验,采用SigmaPlot 12.0软件作图。
2 结果与分析
2.1 SA预处理对低温胁迫下烟草幼苗相对生长速率的影响
如图1所示,10℃低温显著抑制烟草幼苗的生长。‘云烟97’和‘K326’在10℃低温处理12 d后,各处理的相对生长速率均呈负值,表明在这段时间,烟草幼苗不但没有生长且干物质消耗量大于合成量。但经SA预处理后,2个品种幼苗的干物质消耗量显著低于对照(0 mmol/L),在1.0 mmol/L SA预处理下尤为明显。这些结果表明,SA预处理能减缓烟草幼苗在低温胁迫下的干物质消耗速度(图1)。
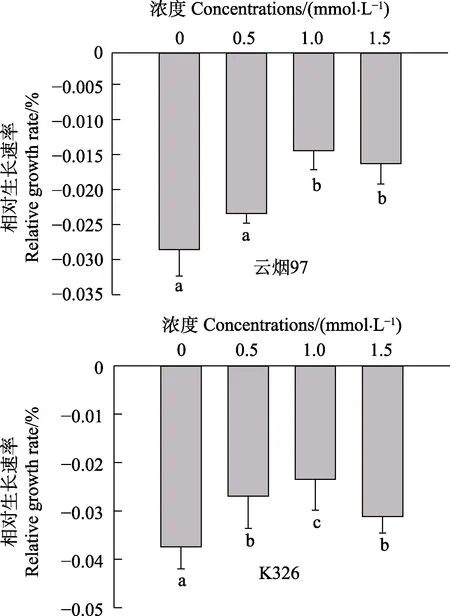
图1 SA预处理对10℃低温胁迫下‘云烟97’和‘K326’幼苗相对生长速率的影响Fig.1 Effect of SA pretreatment on relative growth rate of‘Yunyan 97’ and ‘K326’ seedlings under low temperature stress at 10℃
2.2 SA预处理对低温胁迫下烟草幼苗 MDA含量和电解质渗漏率的影响
MDA含量和电解质渗漏率是衡量植物在低温胁迫下细胞膜受损伤的重要指标[2-3]。图2显示,经0.5、1.0、1.5 mmol/L不同浓度的SA预处理后(即低温胁迫的 0 d),‘云烟 97’和‘K326’幼苗叶片中 MDA含量和电解质渗漏率均比对照有不同程度的减少;在10℃低温胁迫过程中,2个品种的 MDA含量和电解质渗漏率均为先上升后下降。其中,在低温胁迫12 d后,‘云烟97’中经0.5、1.0、1.5 mmol/L SA预处理后的幼苗MDA含量分别比对照降低14.5%、19.6%、19.5%(图2A),电解质渗漏率分别减少 16.3%、19.3%、25.0%(图2C);在‘K326’中,则是 MDA分别减少16.8%、28.6%、20.2%(图2B),电解质渗漏率分别减少14.7%、18.2%、23.6%(图2D)。相比之下,3个SA浓度的作用效果在‘云烟97’中1.5 mmol/L最好,在‘K326’中则是1.0 mmol/L好于另外2个。说明SA预处理能降低烟草幼苗在叶片中的MDA含量和电解质渗漏率,减缓低温下的细胞膜损伤,且这种作用在低温胁迫过程中能一直保持,SA浓度不同,作用效果不同。
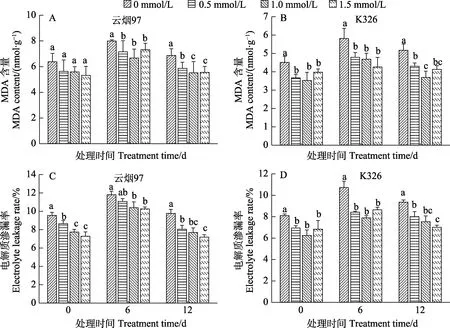
图2 SA预处理对低温胁迫下‘云烟97’和‘K326’幼苗叶片MDA含量(A、B)和电解质渗漏率(C、D)的效应Fig.2 Effects of SA pretreatment on MDA content (A, B) and electrolyte leakage rate (C, D) in ‘Yunyan 97’ and ‘K326’seedling leaves under low temperature stress
2.3 SA预处理对低温胁迫下烟草幼苗总叶绿素含量的影响
在10℃低温胁迫下,2个品种烟草叶片的幼苗绿素含量下降(图3)。从图3可以看出,1.0、1.5 mmol/L SA预处理显著提高了低温处理前‘云烟97’和‘K326’烟草幼苗的叶绿素含量。经低温胁迫12 d后,叶绿素含量比0 d减少,但经0.5、1.0、1.5 mmol/L不同浓度的SA预处理后的幼苗叶绿素含量显著高于对照,‘云烟 97’中 3个不同浓度的处理分别比对照高40.4%、12.9%、9.9%,‘K326’中则分别高17.9%、33.4%、13.4%,表明SA预处理能缓解10℃低温胁迫下烟草幼苗叶片叶绿素的降解。
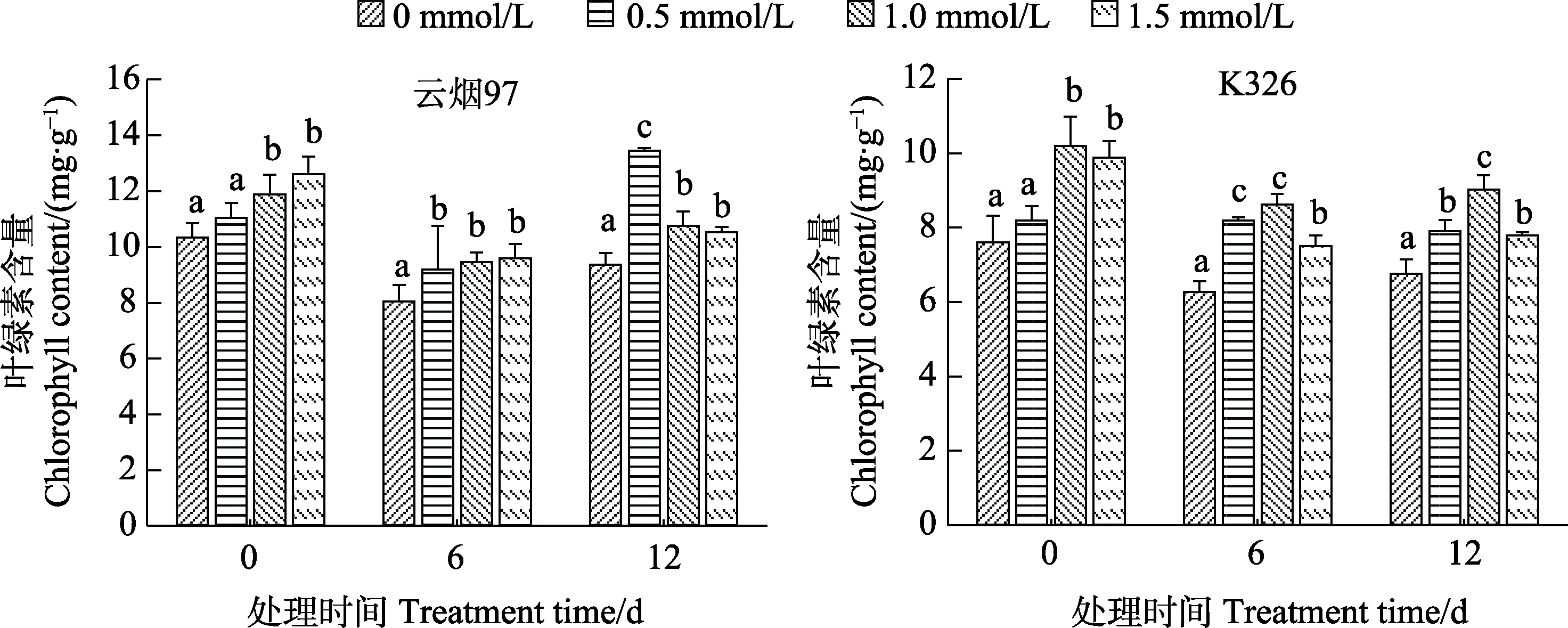
图3 SA预处理对低温胁迫下‘云烟97’和‘K326’幼苗叶片总叶绿素含量的效应Fig.3 Effects of SA pretreatment on total chlorophyll content of ‘Yunyan 97’ and ‘K326’ seedling leaves under low temperature stress
2.4 SA预处理对低温胁迫下烟草幼苗根系活力和根系发育的影响
从图4可以看出,SA预处理会显著提高正常温度培养的‘云烟97’和‘K326’幼苗的根系活力,在‘云烟97’中,0.5、1.0、1.5 mmol/L的SA预处理的根系活力分别比对照提高 15.5%、16.3%、33.1%,在‘K326’中则分别提高65.8%、88.0%、54.5%;在10℃低温胁迫过程中,2个品种的根系活力均是先下降,而后又有所回升,但经3个浓度的SA预处理后,幼苗根系活力普遍显著高于对照,其中‘云烟97’为1.5 mmol/L最显著,‘K326’中则是1.0 mmol/L最好(图4)。

图4 SA预处理对低温胁迫下‘云烟97’和‘K326’幼苗根系活力的效应Fig.4 Effects of SA pretreatment on root vigor of ‘Yunyan 97’ and ‘K326’ seedlings under low temperature stress
对根系扫描的结果表明,SA预处理增加了正常培养条件下‘云烟97’和‘K326’幼苗的根长、表面积、体积和根尖数,但对平均直径的影响不是特别明显(表1、表2)。这些结果说明 SA预处理能促进烟草幼苗根系的发育并使其具有较高的根系活力,并且这种优势在10℃低温胁迫下能一直保持。
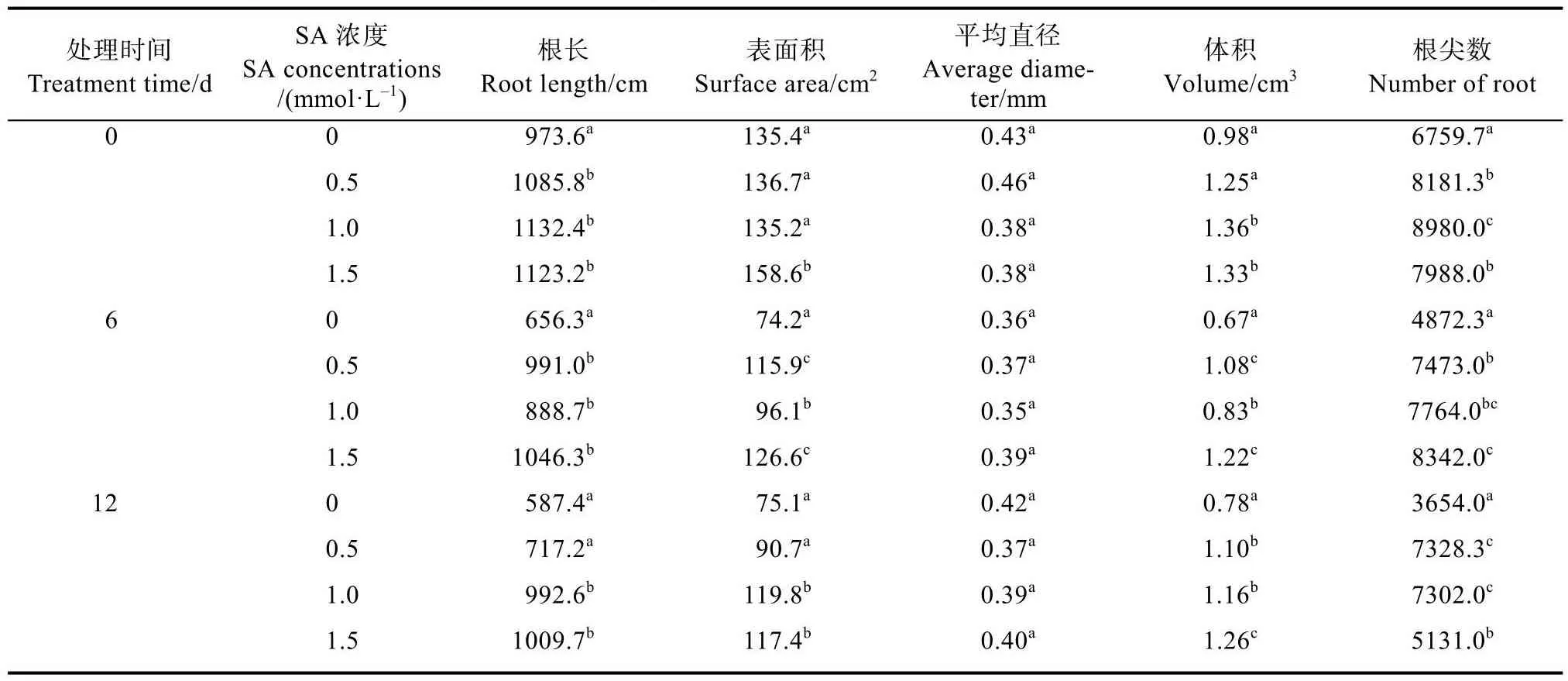
表1 SA预处理对低温胁迫下‘云烟97’幼苗根系发育情况分析Tab.1 Analysis of root development of ‘Yunyan 97’ seedlings with SA pretreatment under low temperature stress
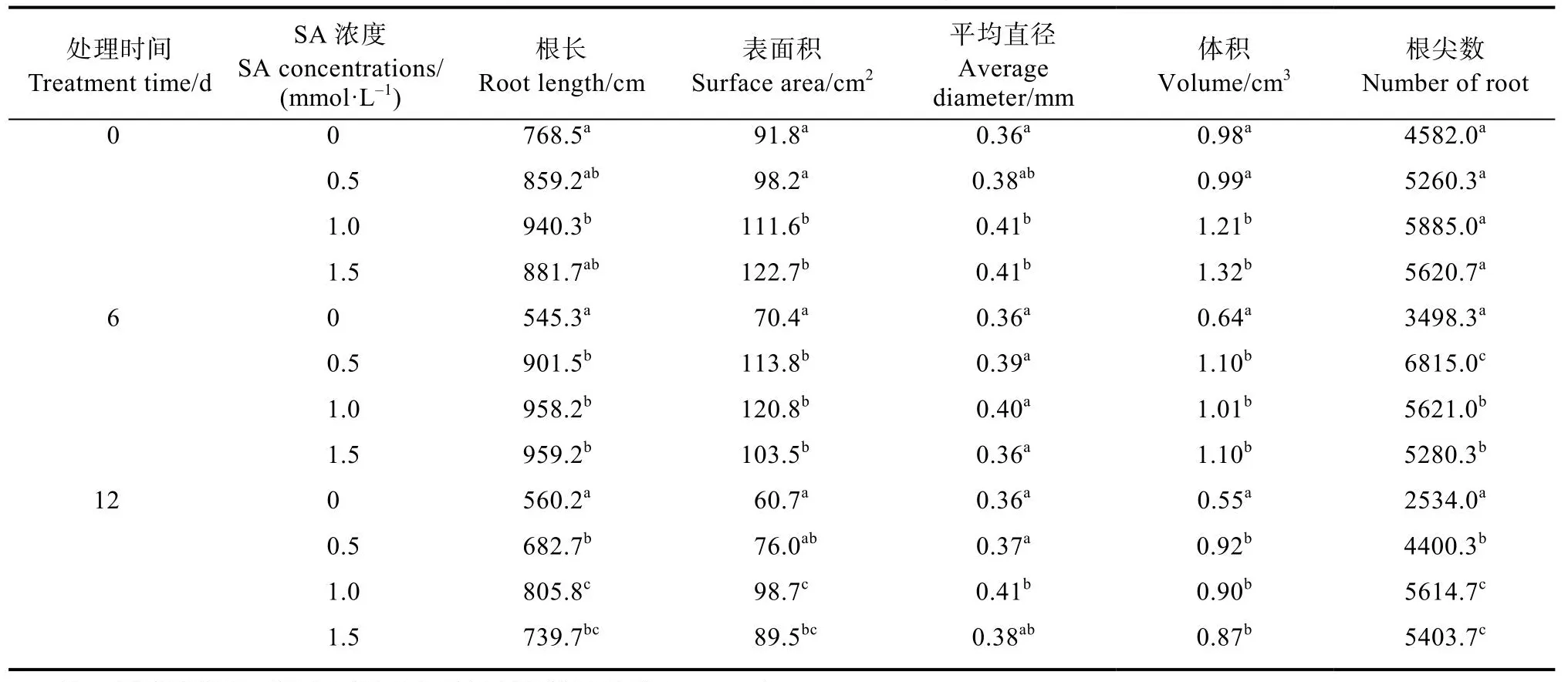
表2 SA预处理对低温胁迫下‘K326’幼苗根系发育情况分析Tab.2 Analysis of root development of ‘K326’ seedlings with SA pretreatment under low temperature stress
2.5 SA预处理对低温胁迫下烟草幼苗抗氧化剂含量的影响
2.5.1 低温胁迫下烟草幼苗还原型/氧化型抗坏血酸的变化 植物在遭受低温胁迫时会产生大量活性氧(ROS),破坏植物的氧化还原平衡,影响植物的正常生长,还原型抗坏血酸ASA是存在于植物体内有效清除ROS的小分子抗氧化剂,对于维持植物的氧化还原平衡有重要作用[1,7]。图5和图6结果表明,SA预处理显著提高了正常培养的‘云烟97’和‘K326’幼苗的ASA含量(图5A、图6A),显著降低了氧化型抗坏血酸 DHA含量(图5B、图6B),总抗坏血酸(ASA+DHA)含量在‘云烟97’中也得到了提高(图5C),对‘K326’则影响不大(图6C),ASA/(ASA+DHA)比值同样得到了提高(图5D、图6D);在低温胁迫结束后,2个品种中的ASA含量、DHA含量和ASA+DHA含量比对照(0 d)均降低,但经SA预处理后的幼苗ASA含量和ASA/(ASA+DHA)比值仍显著高于对照,DHA含量则显著低于对照,ASA+DHA含量差异不显著。结果表明,SA预处理能增加烟草幼苗及其在低温胁迫过程中的 ASA含量和ASA/(ASA+DHA)的比值,而减少DHA含量。
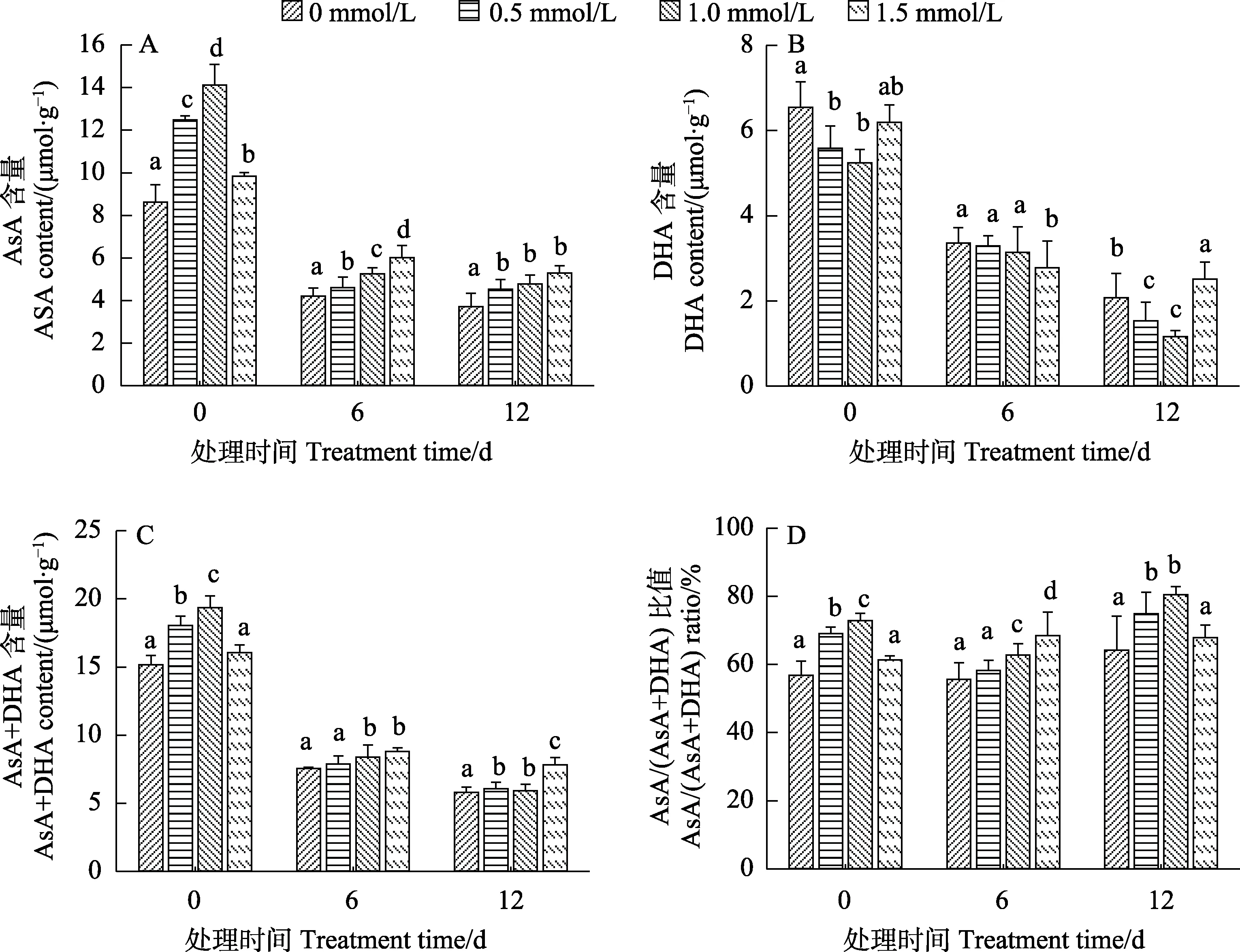
图5 SA预处理对低温胁迫下‘云烟97’幼苗还原型/氧化型抗坏血酸的影响Fig.5 Effects of SA pretreatment on reduced/oxidized ascorbic acid of ‘Yunyan 97’ seedlings under low temperature stress
2.5.2 低温胁迫下烟草幼苗还原型/氧化型谷胱甘肽的变化 谷胱甘肽是植物体内含量极为丰富且富含巯基的低分子肽,能有效清除植物体内的自由基,分为还原型(GSH)和氧化型(GSSG)2种状态[23]。图7和图8结果表明,SA预处理显著提高了正常培养的‘云烟97’和‘K326’幼苗中的GSH含量(图7A、图8A)、GSH+GSSG含量(图7C、图8C)和GSH/(GSH+GSSG)比值(图7D、图8D)。在低温胁迫结束后,经SA预处理过的幼苗,三者的含量均显著高于对照,其中,‘云烟97’中1.5 mmol/L浓度处理的效果最好,‘K326’中则是1.0 mmol/L最好;GSSG含量(图7B、图8B)2个品种基本为先上升后降低,胁迫结束时‘云烟 97’和‘K326’都是 1.0 mmol/L浓度的处理显著低于对照。该结果表明,SA预处理能增加烟草幼苗在低温胁迫过程中的 GSH含量、GSH+GSSG含量和GSH/(GSH+GSSG)比值,浓度不同效果也不同,其中‘云烟 97’中1.5 mmol/L的作用效果最显著,‘K326’中则是1.0 mmol/L最好。
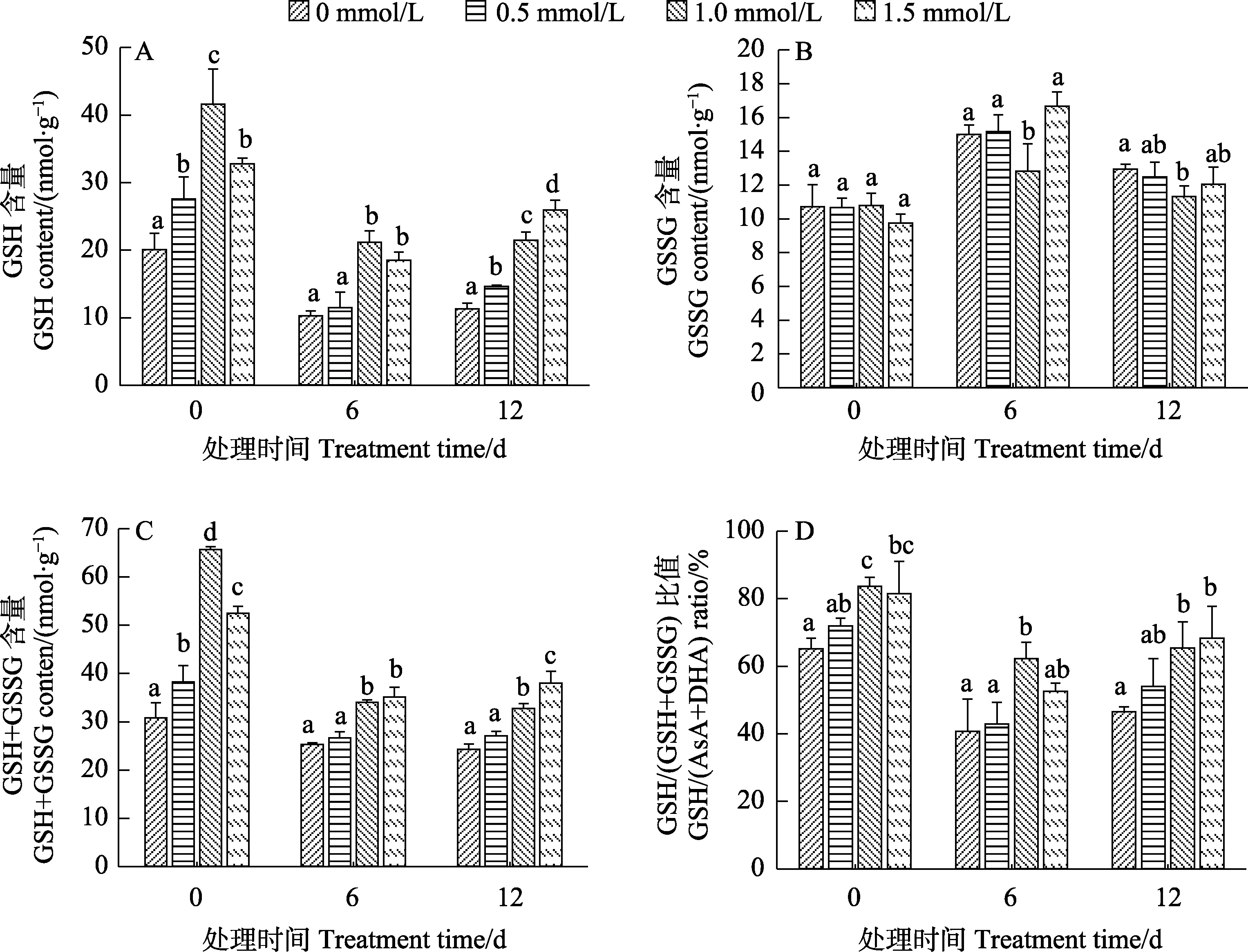
图7 SA预处理对低温胁迫下‘云烟97’幼苗还原型/氧化型谷胱甘肽的影响Fig.7 Effects of SA pretreatment on reduced/oxidized glutathione of ‘Yunyan 97’ seedlings under low temperature stress
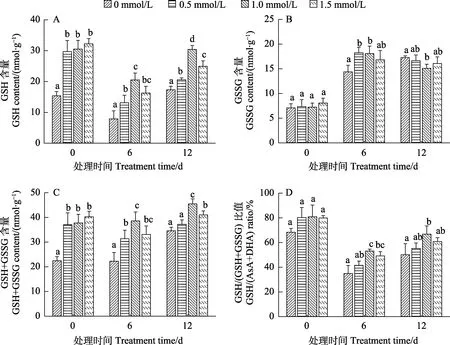
图8 SA预处理对低温胁迫下‘K326’幼苗还原型/氧化型谷胱甘肽的影响Fig.8 Effects of SA pretreatment on reduced/oxidized glutathione of ‘K326’ seedlings under low temperature stress
2.5.3 低温胁迫下烟草幼苗抗氧化酶活性的影响植物细胞抗氧化系统主要由抗氧化剂和抗氧化酶构成。其中,APX、CAT、GPX、GR和SOD是植物细胞最重要的抗氧化酶组分,在清除ROS和维持细胞氧化平衡动态中起重要作用[1,7]。图9结果表明,经SA预处理后,‘云烟97’和‘K326’幼苗的APX活力处理组和对照组差异均不显著,而在低温胁迫过程中,‘云烟97’幼苗的APX活力(图9A)逐渐升高,‘K326’幼苗的 APX活力(图9B)先升高后降低,但 2个品种的 APX活力在低温胁迫的6 d和12 d时经SA预处理过的均显著高于对照,3个浓度中,2个品种均是1.5 mmol/L浓度的处理效果最显著,在胁迫的6 d和 12 d‘云烟 97’的 APX活力分别比对照高75.2%、54.2%,‘K326’中则分别比对照高42.3%、22.6%;SA预处理显著提高了幼苗的CAT活力(图9C、图9D),在胁迫期间,2个品种各个处理的CAT变化趋势基本为逐渐下降,但经SA预处理后的幼苗CAT活力仍高于对照,‘云烟97’中在胁迫6 d为浓度1.0 mmol/L的效果最显著,在12 d则是 1.5 mmol/L最好,在‘K326’中均为1.0 mmol/L浓度的效果最显著;SA预处理也显著提高了幼苗的GPX活力,在胁迫期间,其变化趋势为‘云烟97’中(图9E)先降低后缓慢升高,‘K326’(图9F)则是逐渐降低,胁迫结束后其活力均是SA预处理组显著高于对照组,其中‘云烟 97’为 1.5 mmol/L浓度处理的效果最好,‘K326’为1.0 mmol/L最好;GR变化趋势在2个品种中(图9G、图9H)基本为先降低后缓慢升高,经1.0、1.5 mmol/L SA预处理过的幼苗在胁迫过程中CAT活力均显著高于对照;SOD活力变化在‘云烟97’中(图9I)为先下降后升高,胁迫结束时,3个不同SA预处理浓度的SOD活力分别比对照高21.3%、43.0%、66.8%,在‘K326’中(图9J)SOD活力则是逐渐降低,3个浓度的作用效果在胁迫6 d为1.0 mmol/L最好,在12 d则是1.5 mmol/L最好。以上结果表明,SA预处理能显著提高烟草幼苗的CAT、GPX和GR活力,且在低温胁迫过程中APX、CAT、GPX、GR和SOD五种酶均保持较高活性,显著高于对照,综合比较后,‘云烟 97’中 1.5 mmol/L浓度的效果最好,‘K326’为1.0 mmol/L最好。
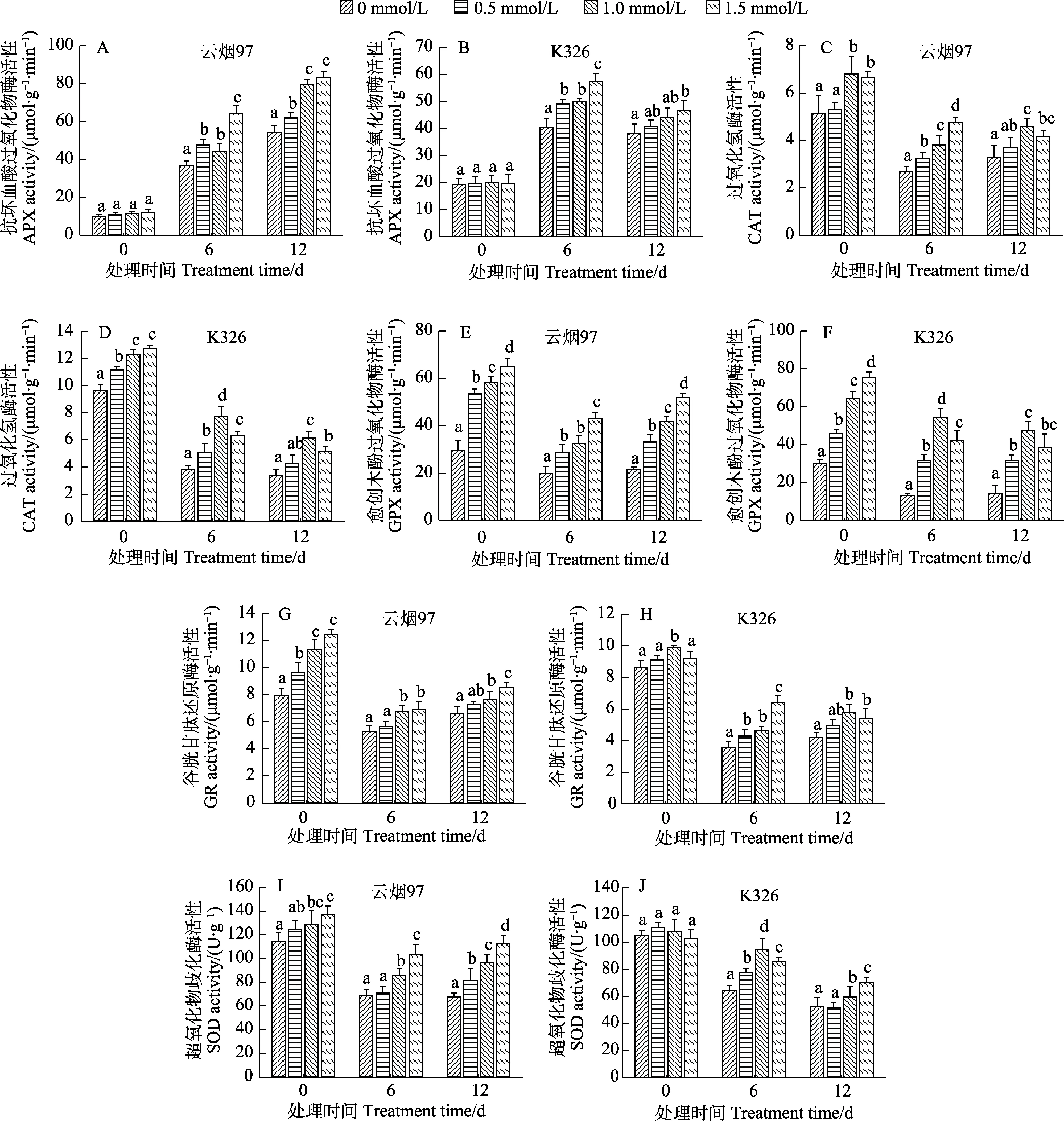
图9 SA预处理对低温胁迫下‘云烟97’和‘K326’幼苗抗氧化酶活性的影响Fig.9 Effects of SA pretreatment on antioxidant enzyme activities of ‘Yunyan 97’ and ‘K326’ seedlings under low temperature stress
3 讨论
起源于热带和亚热带的冷敏植物如水稻、玉米、黄瓜、烟草等在 0℃以上低温胁迫下,对植物的伤害大致可以分为2步:(1)生物膜由液晶态向凝胶态改变,并伴随着原生质流动停止,膜结合酶活性降低,膜透性增大,电解质及某些小分子有机物大量渗漏;(2)由于膜损坏而引起代谢紊乱,细胞 ROS产生与清除的平衡破坏,水解酶类的活性高于合成酶类,致使物质分解大于合成等[12,24-25]。杨利云等[26]、JINI等[27]、YU 等[28]、RAZMI等[29]的研究均表明SA预处理显著降低植物的质膜透性,减轻低温对植物的伤害。本研究结果表明,SA预处理能显著降低2个品种烟草幼苗在低温胁迫过程中的 MDA含量和电解质渗漏率,提高幼苗在低温胁迫前及胁迫过程中叶绿素的含量,且能缓解幼苗在低温胁迫过程中的干物质消耗速度,进而提高烟草幼苗的抗冷性。
植物根系的发育及其活力对地上部的生长状况有着直接影响,较低的土壤温度或灌溉用水水温往往通过抑制根系活力,进而造成植株的低温冷害。对漂浮育苗的烟草幼苗而言,高的根系活力和根群分布直接影响着烟草幼苗移栽到大田后尽早成活和早生快发,进而影响烟叶的产量和质量。课题组前期研究结果表明,SA预处理能促进正常培养条件下的烟株移栽后的早生快发[30]。目前研究结果表明,与未处理对照相比,SA预处理能显著提高烟草幼苗的根系活力,且增加根长、表面积、体积和根尖数,缓解低温胁迫对根系生长的抑制,该效应有助于烟草幼苗在移栽后的早成活或早生快发,对烟草幼苗的农艺管理和早期栽培有参考意义。
以上结果表明,SA预处理能降低烟草幼苗在低温胁迫过程中的 MDA含量和电解质渗漏率,减缓干物质的消耗速度,提高叶绿素含量和根系活力,促进根系生长,最终提高烟草幼苗的抗冷性,烟草品种不同,需要的最适SA浓度不同,综合比较之下,‘云烟 97’的最适浓度为 1.5 mmol/L,‘K326’为1.0 mmol/L。
但是 SA预处理为何能诱导烟草幼苗形成抗冷性?其中的生化机制尚待进一步阐明。低温对植物的伤害与ROS的大量产生密切相关,为此,植物体需要酶促和非酶促两类防御系统来保护机体不受伤害,ASA和 GSH是植物体内重要的非酶促类抗氧化物质,能清除ROS,降低氧化伤害,细胞的还原型/氧化型抗氧化剂比例代表着细胞的氧化还原状态,而维持较高的还原型/氧化型抗氧化剂比例有助于提高植物的抗逆性。邓世媛等[12]研究表明,SA预处理的烤烟叶片中ASA含量增加。本研究结果表明,SA预处理能显著提高在低温胁迫前即0 d时的ASA含量、GSH含量和还原型抗氧化剂在总抗氧化剂中的比值,经低温胁迫后,这三者的含量SA预处理组仍高于对照,品种及SA浓度不同,效果不同,‘云烟97’中以1.5 mmol/L浓度的效果最佳,‘K326’以1.0 mmol/L最好。
APX、CAT、GPX、GR和 SOD等抗氧化酶活性的高低在植物体清除 ROS中扮演着非常重要的作用。杨小环等[31]用0.5 mmol/L SA预处理玉米种子发现SA显著增加了SOD和POD活性,YU等[28]在研究SA预处理与冬小麦抗冷性时,发现SA能提高SOD、POD、CAT、APX活性,减少低温伤害。本研究中,SA预处理能显著提高‘云烟 97’和‘K326’幼苗中的 CAT、GPX和 GR活力,对APX和SOD活力无显著影响,在经过10℃的低温胁迫12 d后,APX活性增加,CAT、GPX、GR和SOD活性均出现不同程度的下降,但经SA处理后,幼苗中5种抗氧化酶活性均显著高于对照组,‘云烟97’中1.5 mmol/L浓度的处理效果最好,‘K326’则为1.0 mmol/L最好。
此外,抗氧化酶APX和GR分别在维持细胞还原型/氧化型抗氧化剂抗坏血酸和谷胱甘肽的动态平衡中起重要作用,高活力的APX和GR有助于提高细胞的还原型ASA和GSH含量,使细胞保持一个更高的抗氧化能力[23,32]。本研究结果可明显看出,与对照相比,SA预处理能显著提高‘云烟97’和‘K326’幼苗在低温下的APX和 GR活力,并保持较高水平的还原型 ASA和GSH含量,这暗示着SA处理在某种程度上是通过维持 APX和 GR相对较高的活力使烟草幼苗在低温下能保留较高的还原型 ASA和 GSH含量。SA处理是否能通过激活APX和GR的基因表达、从而使其有较高的酶活力尚待进一步深入的研究。
‘云烟97’和‘K326’均为云南省和全国主要烟区的烟草主栽品种。从2个品种在人工气候室进行10℃低温处理12 d的表型及本研究所测的生理指标来看,其抗冷性无显著差异,但不同的品种需要不同的适宜 SA预处理浓度来提高其抗冷性。对比2个品种在低温处理过程的抗氧化系统来看,‘云烟97’有更高的还原型GSH含量,而所测的5个抗氧化酶在2个品种中的变化不尽相同,这可能是由于其遗传背景不同所致。但一个共同的趋势是,SA预处理后,与对照相比,在进入低温处理前和低温处理期间,烟草幼苗均能保持显著较高的还原型抗氧化剂水平和抗氧化酶活力,这可能是SA处理能诱导2个品种烟草幼苗抗冷性提高的重要生化基础。
4 结论
本研究以 2个国内的主栽烟草品种为材料,系统调查了 SA预处理对漂浮育苗的烟草幼苗抗冷性的影响,证实了 SA预处理能显著提高烟草幼苗的抗冷性,不同的品种需要不同的适宜 SA预处理浓度,‘云烟97’的最适浓度为1.5 mmol/L,‘K326’的最适浓度为 1.0 mmol/L。SA预处理后抗氧化系统中还原型抗氧化剂水平和还原型抗氧化剂在总抗氧化剂中的比例提高,部分抗氧化酶活性的增强以及这种优势在低温胁迫过程中的维持可能是 SA预处理诱导烟草幼苗抗冷性形成的重要生理基础。研究结果不仅对烟草抗冷育苗有参考意义,也对其他农作物和蔬菜的育苗和早期栽培有借鉴作用。
猜你喜欢
初中生学习指导·提升版(2023年11期)2023-12-16 12:44:18
奥秘(创新大赛)(2023年3期)2023-05-06 01:48:20
农业技术与装备(2020年10期)2020-11-03 11:25:44
浙江中西医结合杂志(2017年2期)2017-01-12 18:23:59
当代化工研究(2016年9期)2016-03-20 16:22:08
河北书画研究(2016年6期)2016-02-03 02:57:44
中国继续医学教育(2015年3期)2016-01-06 01:36:43
诗歌月刊(2014年3期)2015-04-14 17:50:51
传奇故事(破茧成蝶)(2015年8期)2015-02-28 09:29:41
声屏世界(2014年6期)2014-02-28 15:18:09
