
施用生物炭和硅肥对增温稻田土壤酶活性的影响
2023-02-15 01:44杜泽云陶思敏娄运生邢钰媛
江苏农业科学 2023年1期
杜泽云, 陶思敏, 娄运生, 邢钰媛, 王 坤, 刘 健
(南京信息工程大学气象灾害预报预警与评估协同创新中心/南京信息工程大学江苏省农业气象重点实验室,江苏南京 210044)
政府间气候变化专门委员会(IPCC)第6次评估报告指出,1850年以来,全球地表平均温度已上升约1 ℃,未来20年温升预计将达到或超过 1.5 ℃[1]。气候变暖表现为昼夜不对称性增温,即相较于白天,夜间温度增幅更大。水稻是我国主要的粮食作物之一。夜间增温显著降低水稻分蘖数、株高、叶面积指数和叶绿素(SPAD值)[2-5],降低水稻穗粒数、结实率和千粒质量,从而影响干物质积累和产量[6-8]。目前,增温对水稻地上部植株生长、产量及干物质积累的影响研究较多,但对水稻地下部土壤生物过程和特性(如土壤酶活性)有何影响,则关注较少。
土壤酶主要来自植物根系分泌物和土壤微生物活动产物,参与土壤中的物质循环和能量流动过程[9]。温度是影响土壤酶活性的重要环境因子。以往研究表明,模拟增温处理下,麦田土壤酶活性显著增强[10],温度与土壤酶活性间存在显著指数关系[11]。温度升高促进土壤呼吸、增加酶活性,有利于土壤微生物活动和新陈代谢,增强土壤养分利用率,促进土壤物质循环和能量转化。
生物炭富含有机碳,是一种有益的土壤改良剂[12]。施用生物炭可明显降低土壤容重,改善土壤结构,增加土壤含水量。施用生物炭对土壤氮磷相关酶活性有显著影响[13-14],生物炭可通过吸附酶分子来限制酶促反应[15]。施用生物炭可提高土壤过氧化氢酶活性,对蔗糖酶活性提高的贡献较小[16],但与氮肥结合后水稻土壤蔗糖酶活性显著提高[17]。
水稻是典型的喜硅作物,穗硅含量为10%~15%,远高于氮磷钾的含量。施用硅可以促进土壤酶活性[18-19]。施硅可明显提高土壤脲酶、转化酶、过氧化氢酶等活性[20],其原因可能在于,施硅改善了土壤结构,扩大了土壤微生物生长空间,进而提高了土壤酶活性[21]。
前人有关稻田土壤养分对增温、施生物炭、施硅的单因素或双因素响应开展了大量研究工作[22-24],得到一些对生产有指导意义的结论,但三因素耦合作用对土壤酶活性的影响,目前报道较少。本文通过田间模拟试验探讨夜间增温下施用生物炭和硅肥对水稻关键生育期稻田土壤酶活性的影响,以期进一步阐明气候变暖对稻田土壤碳氮转化及温室气体产生和排放机制的影响,为水稻低碳绿色生产提供试验参考依据。
1 材料与方法
1.1 试验区概况
2019年6—10月于南京信息工程大学农业气象试验站(118.8°E、32.0°N)进行田间模拟试验。该站位于亚热带季风气候区,年均气温为15.6 ℃,年均降水量大于1 000 mm。供试稻田土壤为灰马肝土属潴育型水稻土,土壤有机碳含量 19.4 g/kg,全氮含量1.45 g/kg,黏粒含量26.1 g/kg,pH值6.2(1 g ∶1 mL土水比)。供试生物炭为稻壳生物炭,碳含量为50%。供试硅肥选用含有效硅(SiO2)14.21%、铁氧化物(Fe2O3)22.89%、pH值为8.09(土水比1 g ∶10 mL)的钢渣硅肥和有效硅(SiO2)含量32.26%、铁氧化物(Fe2O3)含量0.81%、pH值为9.22(土水比1 g ∶10 mL)的矿粉硅肥[25]。
1.2 试验设计
本试验采用3因素3水平正交试验设计(表1)。移栽前按试验处理将生物炭和硅肥分别均匀翻耕混入稻田土壤,同时各小区施入200 kg/hm2氮磷钾高浓度复合肥料(N-P2O5-K2O,俄罗斯产)作为基肥。
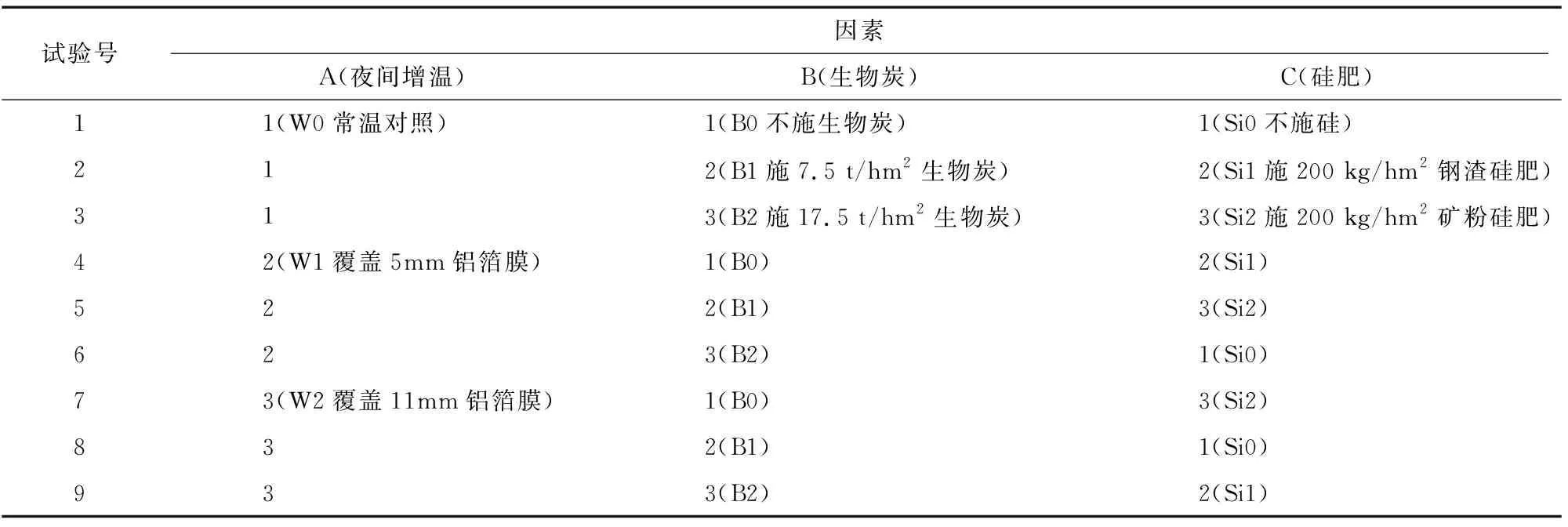
表1 L9(34)正交试验设计方案
选用南粳5055为供试水稻品种,育苗期为2019年5月10日至6月13日,6月14日移栽。采用开放式被动增温方式模拟夜间增温,在小区四周架设不锈钢架,夜间(19:00至次日6:00)将铝箔反光膜置于钢架上方,用来覆盖水稻的冠层,及时调整铝箔膜的高度以适应水稻生长,保持铝箔膜与植株冠层间距为0.3 m。为防止降雨及大风天气(风速>10 m/s)覆盖铝箔膜所带来的不利影响,故夜间有雨和大风天气不覆盖铝箔膜。小区面积为 4 m2(2 m×2 m),随机排列。稻田水深保持在5 cm左右,晒田期为2019年7月28日至8月12日。
水稻移栽时每小区选取长势良好、大小相近的幼苗移入根袋内植于稻田土壤中,1个根袋植入1株水稻幼苗,每小区4个根袋,在水稻4个主要生育期各采集1个根袋水稻植株,分别于根袋内及根袋外采集土样,根袋内为根际土,根袋外非根际土[26-27]。将所采土样装入塑料自封袋,挑去根系残体、石块等杂物,然后把土样放置在通风处风干,研磨后用筛子筛选干净,使用塑料保鲜袋保存备用。
1.3 土壤酶活性测定
土壤脲酶活性采用靛酚比色法[28]测定;土壤转化酶活性采用比色法测定;土壤纤维素酶活性采用蒽酮比色法测定;土壤蛋白酶活性采用茚三酮比色法[29]测定。
1.4 数据处理
采用SPSS 19.0软件进行试验方差分析和极差分析,采用Excel 2019、Origin Pro 2021进行数据处理和图表绘制。
2 结果与分析
2.1 不同处理对水稻土脲酶活性的影响
由图1可知,随着水稻生长期变化,根际土脲酶活性部分呈现先增加而后降低趋势,孕穗期脲酶活性较高,开花期活性较弱,除W1B2Si0、W2B0Si2、W1B1Si2处理外,其余处理的根际土脲酶活性,开花期脲酶活性最低,拔节期和孕穗期根际土脲酶活性最高。不同处理的非根际土脲酶活性总体变化幅度不大,表现为随着水稻的生长先平稳而后降低趋势,灌浆期除W0B1Si1和W1B0Si1处理外,其余处理的非根际土脲酶活性灌浆期最低。根际土中,W0B2Si2处理的4个生育期均值最高,W2B1Si0处理的均值最低;非根际土中,W1B2Si0处理的均值最高,W0B0Si0处理均值最低。

平均值和极差分析结果表明,在根际土中,三因素的影响程度从大到小依次为:生物炭、夜间增温、硅肥。W1条件下,脲酶活性最高,W1B2Si1为最优水平。对于非根际土,三因素的影响程度从大到小依次为:夜间增温、生物炭、硅肥(表2)。W1条件下脲酶活性最高,W1B2Si1为最优水平。

表2 水稻全生育期脲酶均值极差分析
方差分析结果表明(表3),在根际土中,夜间增温(拔节期)和施用生物炭(孕穗期)对土壤脲酶活性的影响达到显著水平(P<0.05),而施用硅肥的影响不显著。在非根际土中,夜间增温对水稻拔节期和开花期对脲酶活性的影响达到极显著水平(P<0.01),而施用生物炭和硅肥对关键生育期水稻土脲酶活性的影响均不显著。可见,夜间增温在水稻部分生育期影响脲酶活性,其影响随着覆膜的厚度的增加而减小;而施生物炭仅在根际土孕穗期显著影响脲酶活性,且随着生物炭施用量的增加,影响逐渐增大。

表3 脲酶活性方差分析
2.2 不同处理对水稻土纤维素酶活性的影响
如图2所示,随着水稻生长期变化,根际土纤维素酶活性总体上呈现先降低而后增加趋势,开花期和灌浆期纤维素酶活性较高,孕穗期活性较低,并出现最小值。不同处理的非根际土纤维素酶活性总体呈现先增加而后降低的趋势,拔节期纤维素酶活性较低且达到最小值,孕穗期明显增加,开花期又开始降低(W0B0Si0、W1B0Si1除外),最后W1B2Si0、W2B0Si2、W2B1Si0、W2B2Si1处理在灌浆期缓慢增加。根际土中,W2B2Si1处理的4个生育期纤维素酶活性均值最高,W1B2Si0处理的均值最低;非根际土中,W1B1Si2处理的均值最高,W0B0Si0处理的均值最低。

平均值和极差分析表明,在根际土中,三因素的影响程度从大到小依次为:夜间增温、生物炭、硅肥。在根际土中,W2条件下纤维素酶活性最高,W2B1Si2为最优水平。在非根际土中,三因素的影响程度从大到小依次为:硅肥、夜间增温、生物炭,W1条件下纤维素酶活性最高, W1B1Si1为最优水平(表4)。
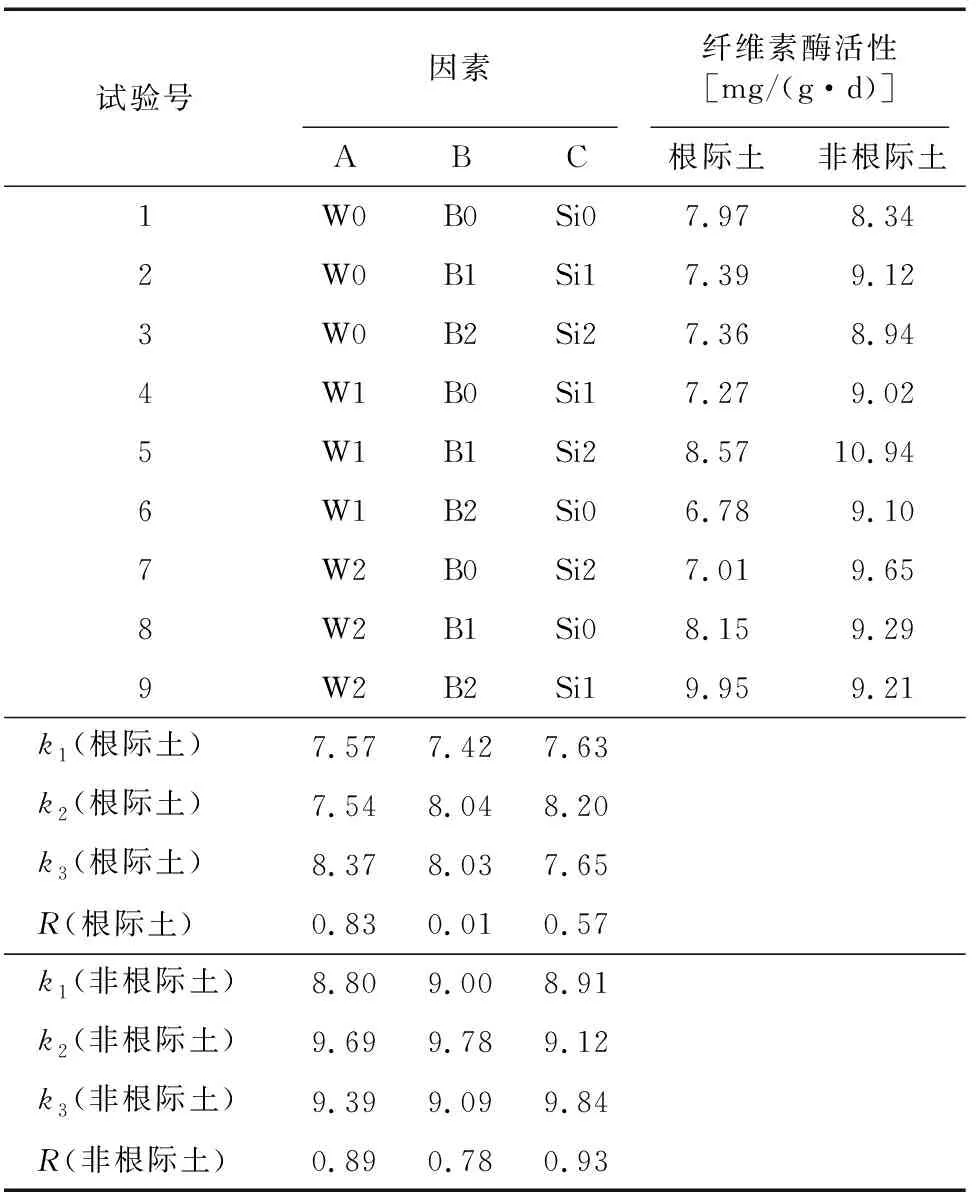
表4 水稻全生育期纤维素酶均值极差分析
方差分析表明(表5),根际土中,夜间增温、生物炭和硅肥均对水稻土纤维素酶活性无显著影响。非根际土中,硅肥对水稻拔节期纤维素酶活性有极显著影响,夜间增温和生物炭对纤维素酶活性无显著影响。

表5 纤维素酶活性方差分析
2.3 不同处理对水稻土转化酶活性的影响
如图3所示,随水稻生长期变化,根际土各处理转化酶活性变化趋势不同,部分处理孕穗期和开花期转化酶活性较高,拔节期活性较低。W0B1Si1、W0B2Si2、W2B0Si2处理的非根际土转化酶活性变化趋势总体上呈现先增加—降低—增加趋势。根际土中,W1B1Si2处理的4个生育期转化酶活性均值最高,W2B2Si1处理的均值最低;非根际土中,W1B0Si1处理的均值最高,W2B1Si0处理均值最低。

平均值和极差分析表明,在根际土中,三因素的影响程度从大到小依次为:夜间增温、生物炭、硅肥。在根际土中,W1条件下转化酶活性最高, W1B0Si1为最优水平。在非根际中,三因素的影响程度从大到小依次为:生物炭、夜间增温、硅肥(表6)。W1条件下转化酶活性最高, W1B0Si2为最优水平。

表6 水稻全生育期转化酶均值极差分析
方差分析表明,根际土中,夜间增温(孕穗期)和施用生物炭(灌浆期)对水稻土转化酶活性的影响达极显著水平,硅肥(开花期)对转化酶活性有显著影响。非根际土中,三因素对转化酶活性均无显著影响(表7)。
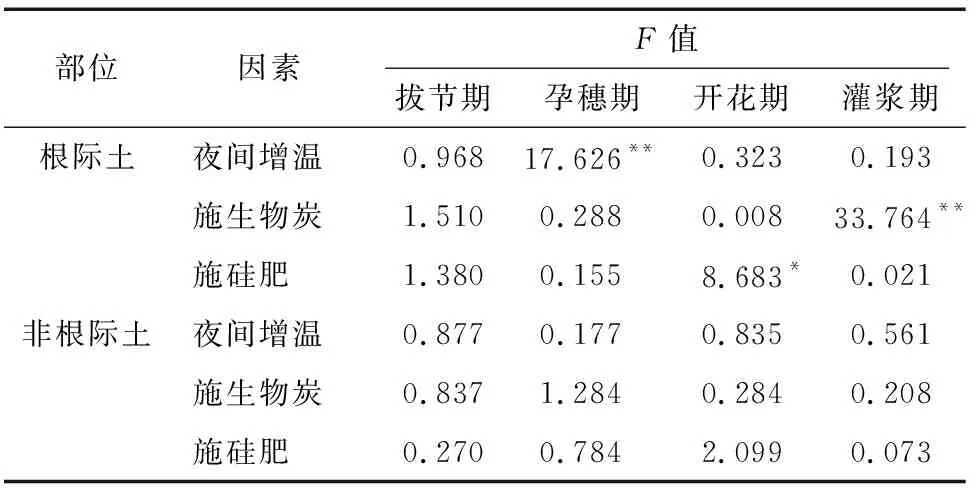
表7 转化酶活性方差分析
2.4 不同处理对水稻土壤蛋白酶活性的影响
由图4可知,随水稻生长期变化,根际土蛋白酶活性总体变化幅度不大。不同处理的非根际土蛋白酶活性变化趋势不同。根际土中,W0B0Si0处理蛋白酶活性均值最高,W2B2Si1处理的均值最低;非根际土中,W1B0Si1处理的均值最高,W1B2Si0处理的均值最低。

极差分析(表8)表明,对于根际土,三因素的影响程度从大到小依次为:生物炭、夜间增温、硅肥。在根际土中,W0条件下蛋白酶活性最高, W0B0Si1为最优水平。在非根际中,三因素的影响程度从大到小依次为:生物炭、硅肥、夜间增温。W0条件下蛋白酶活性最高, W0B0Si1为最优水平。

表8 水稻全生育期蛋白酶均值极差分析
方差分析(表9)表明,根际土中,夜间增温和施用硅肥对水稻土蛋白酶活性无显著影响, 施用生物炭在拔节期对蛋白酶活性有显著影响,并在孕穗期和灌浆期达到极显著水平。非根际土中, 施用生物炭对开花期和灌浆期蛋白酶活性的影响达到显著水平,夜间增温和施用硅肥对蛋白酶活性均无显著影响。

表9 蛋白酶活性方差分析
3 讨论
土壤酶活性是评价土壤质量和土壤生物活性的常用指标[9],在土壤养分循环转化中起着重要作用[30-32]。温度是影响土壤酶活性的重要环境因子[33]。本研究发现,夜间覆盖5 mm铝箔膜(W1)可促进稻田土壤根际土和非根际土脲酶活性、转化酶活性以及非根际土纤维素酶活性;而随着夜间覆膜厚度的增加,其对根际土和非根际土转化酶活性、蛋白酶活性以及根际土脲酶活性的影响减小。其原因可能是:(1)在适宜的温度范围内,土壤增温可以提高微生物的繁殖和代谢速度,产生较多的酶参与碳氮循环,扩大酶库增强酶活性。温度较高时,超过土壤蛋白酶活性的适宜温度范围,引起酶活性下降。(2)夜间增温使土壤铵态氮含量增加,铵态氮能抑制土壤脲酶和转化酶的活性[34]。增温对土壤酶活性的影响差异,可能是土壤和其它环境因子共同作用的结果[35]。
生物炭可通过改变土壤理化性质、土壤生物群落组成和丰富度以影响土壤酶活性。生物炭对土壤酶活性的影响较复杂,主要与土壤酶的类型有关[36]。生物炭可吸附酶底物,促进酶促反应;相反,酶分子也可被生物炭吸附,以掩蔽酶促反应的结合位点,抑制酶促反应[37]。本研究发现,在增温处理下施用生物炭可增加土壤脲酶活性。原因可能是温度和生物炭共同作用改善了土壤环境,增加了土壤有机质含量和土壤温度[36]。提高生物炭施用量,稻田土壤转化酶和蛋白酶活性呈下降趋势。其原因可能是生物炭具有多孔结构和较大比表面积,具有较强的吸附能力,引起土壤酶活性的反应底物降低[39]。
施用硅肥提高了水稻脲酶活性、纤维素酶活性、转化酶活性和根际土蛋白酶活性,并对拔节期非根际土纤维素酶活性和开花期根际土转化酶活性有显著促进作用。原因可能是施用钢渣硅肥和矿粉硅肥可改善稻田土壤结构和通气性,缓解土壤酸度,使pH值趋中性,改善土壤微生物环境,有利于土壤微生物活动,提高土壤酶活性[40]。
4 结论
本研究认为,本试验最佳处理组合为W1B0Si1,即夜间覆盖5 mm铝箔膜、施用200 kg/hm2钢渣硅肥但不施生物炭处理下,水稻主要生育期土壤酶活性可达较优水平。本研究为水稻大田模拟增温试验,易受降水、温度等环境因素的影响,拟通过田间连续定位试验及环境控制试验,深入探讨稻田土壤酶活性对气候变化、不同类型生物炭施用方式及水肥管理的响应特征,进一步研究气候变化对稻田土壤碳氮循环转化相关酶活性的影响机制,为区域水稻绿色低碳可持续发展提供参考依据。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
探索科学(学术版)(2021年1期)2021-04-26
生物技术通报(2020年12期)2020-12-21
腐植酸(2020年1期)2020-11-29
江西农业(2020年10期)2020-07-23
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2018年1期)2018-03-15
中国蔬菜(2016年8期)2017-01-15
现代农业(2016年4期)2016-02-28
